Assessment of the Main Compounds of the Lipolytic System in Treadmill Running Rats: Different Response Patterns between the Right and Left Ventricle

 , ,
, ,
Abstract
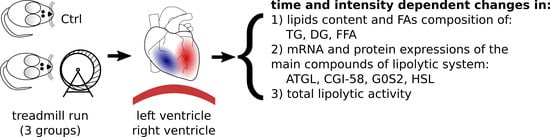
:
1. Introduction
2. Results
2.1. Blood Plasma FFA Content
2.2. The Left Ventricle
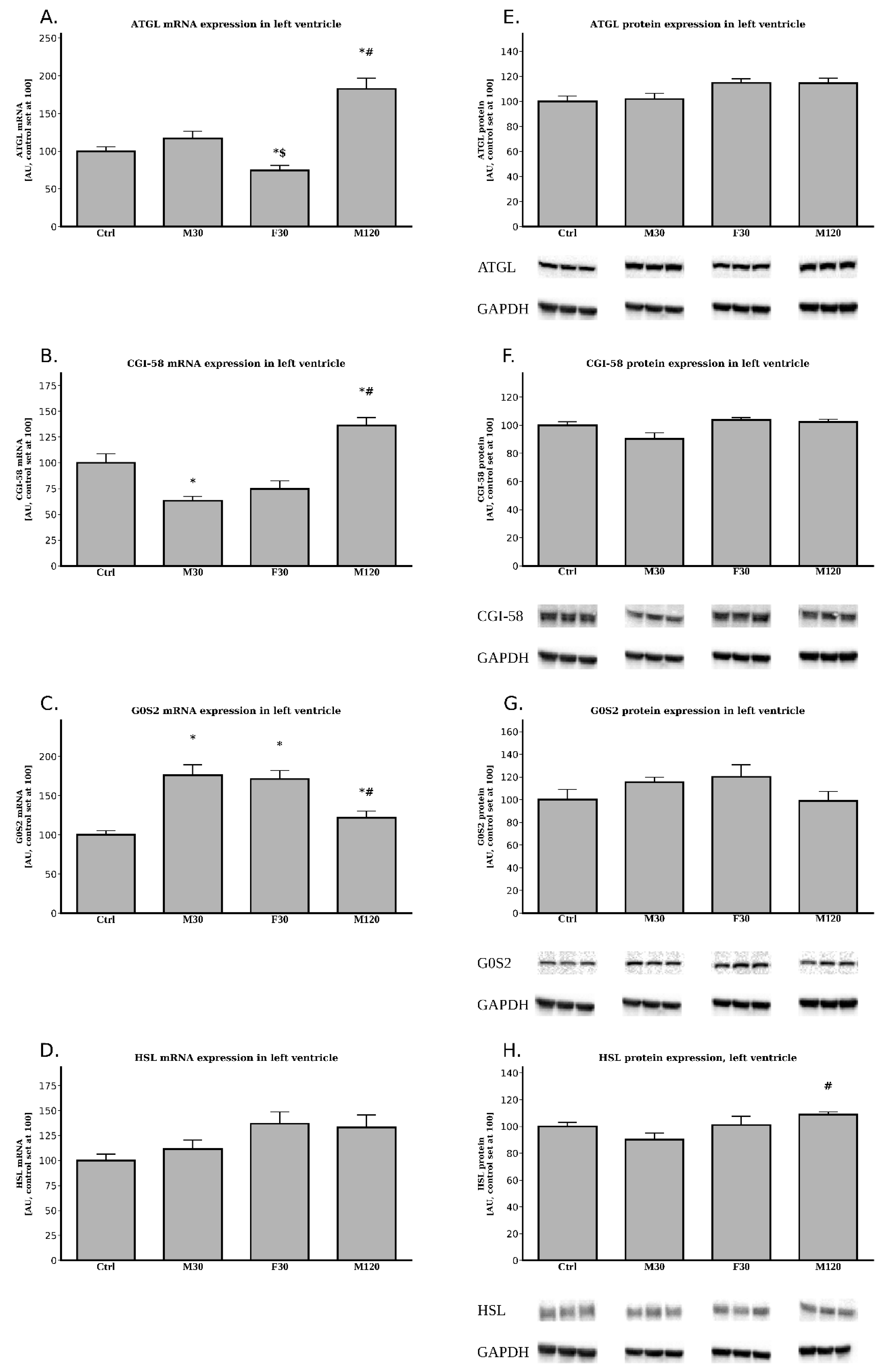
2.2.1. The Expression of ATGL, CGI-58, G0S2 and HSL at the Post-Transcript (mRNA) Level
2.2.2. The Expression of ATGL, CGI-58, G0S2 and HSL at the Protein Level
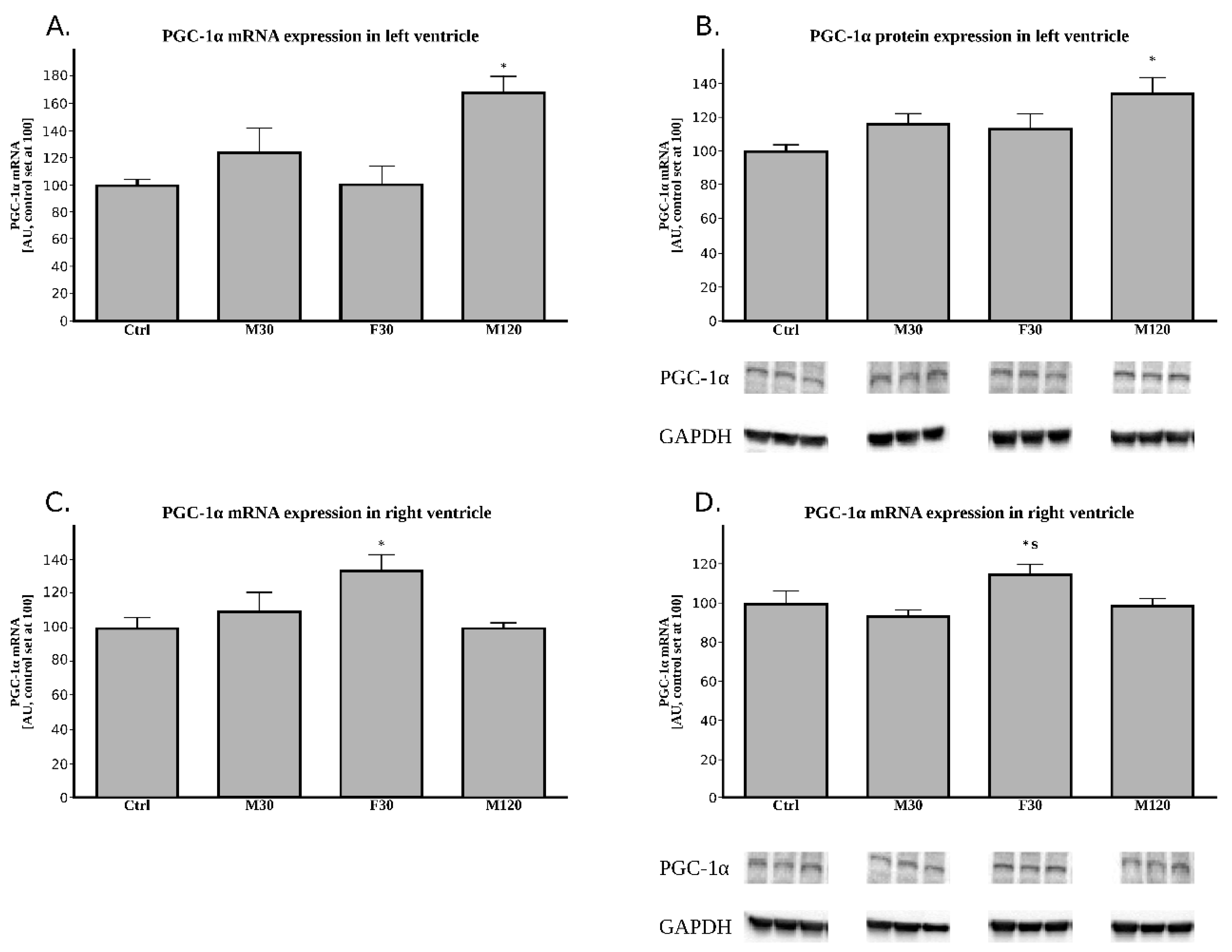
2.2.3. The Expression of PGC-1α (Peroxisome Proliferator-Activated Receptor Gamma Coactivator 1-Alpha) at the Post-Transcript (mRNA) Level
2.2.4. The Expression of PGC-1α (Peroxisome Proliferator-Activated Receptor Gamma Coactivator 1-Alpha) at the Protein Level
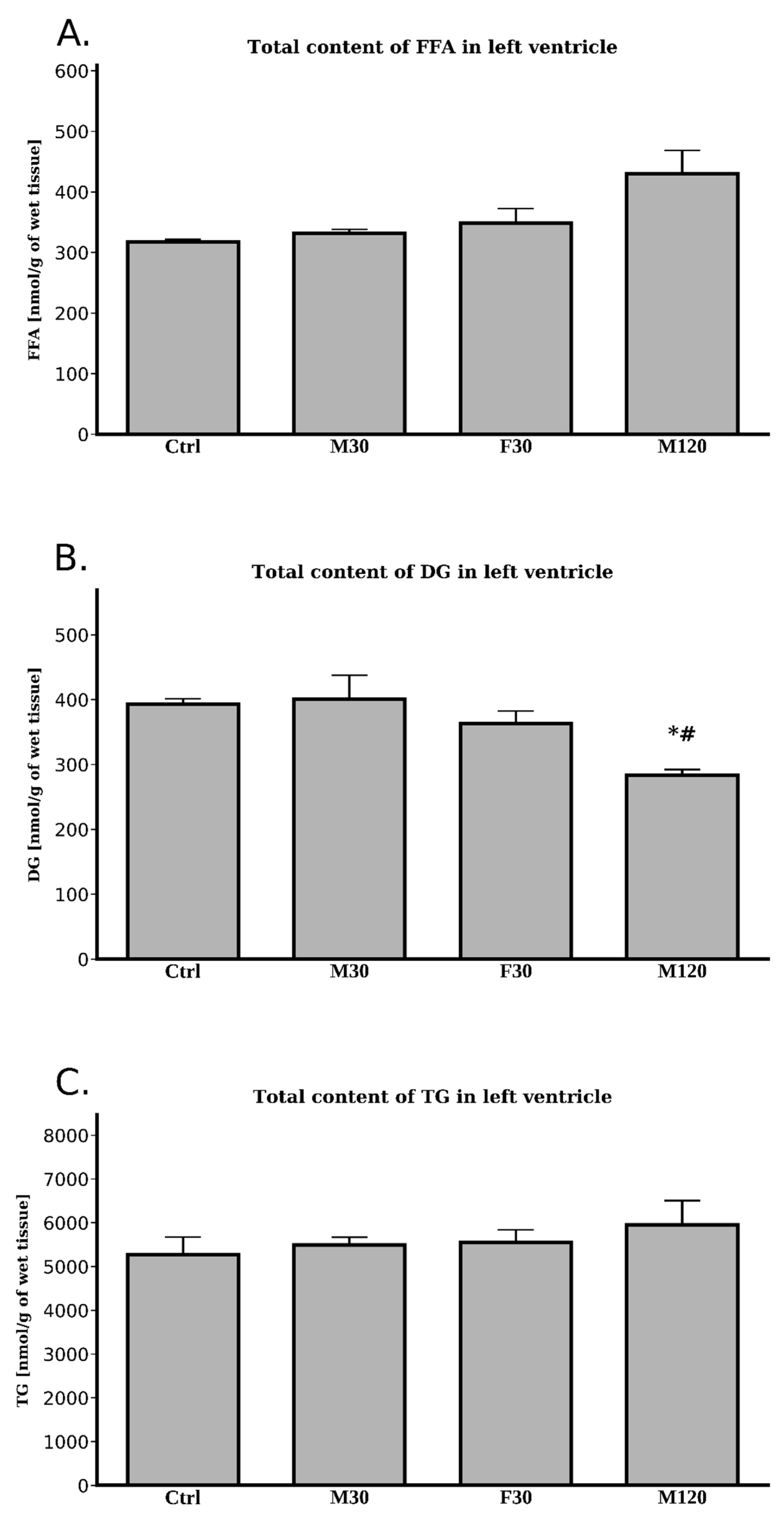
2.2.5. Lipids Content
Free Fatty Acids (FFA)
Diacylglycerols (DG)
Triacylglycerols (TG)
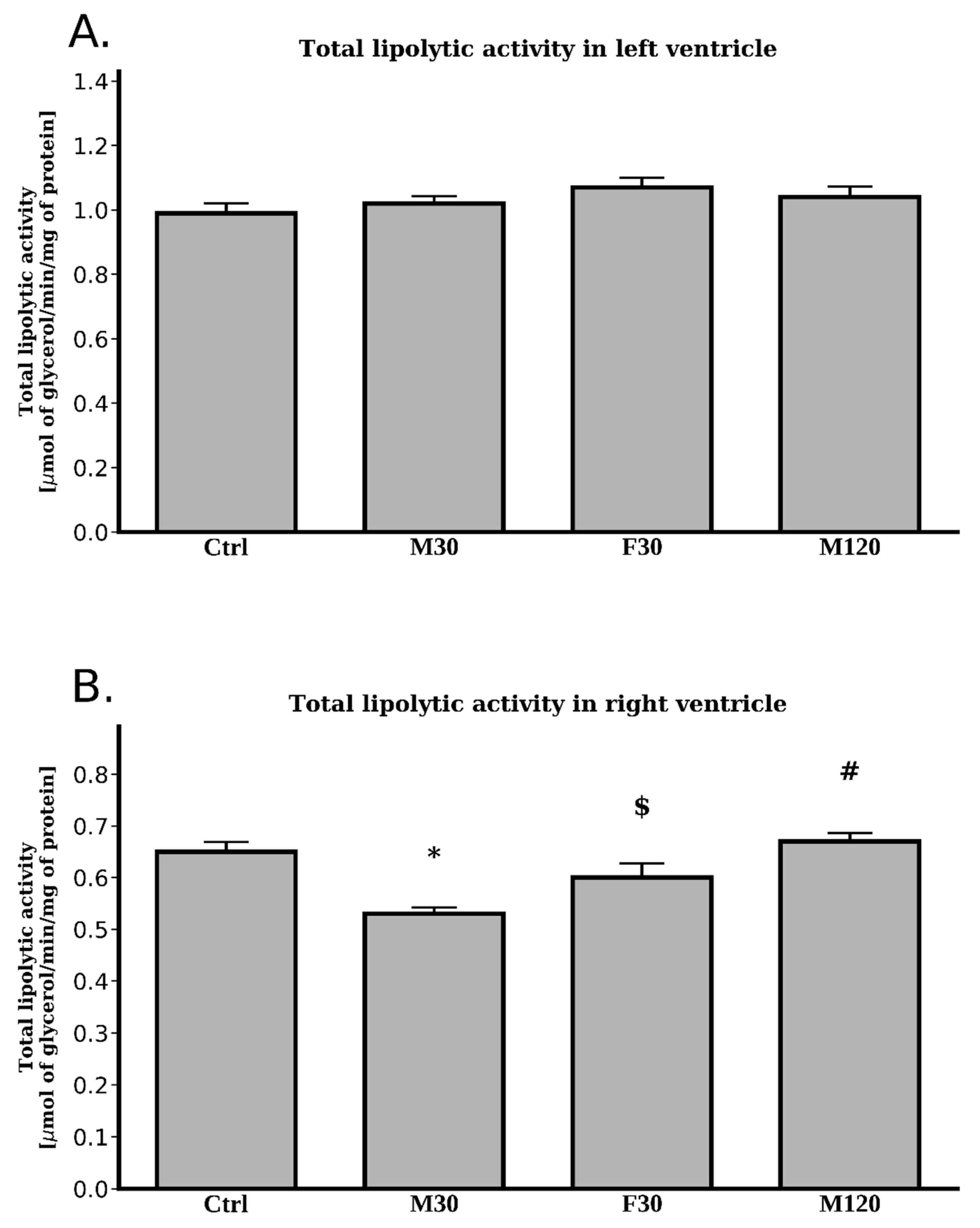
2.2.6. Total Lipolytic Activity
2.3. The Right Ventricle
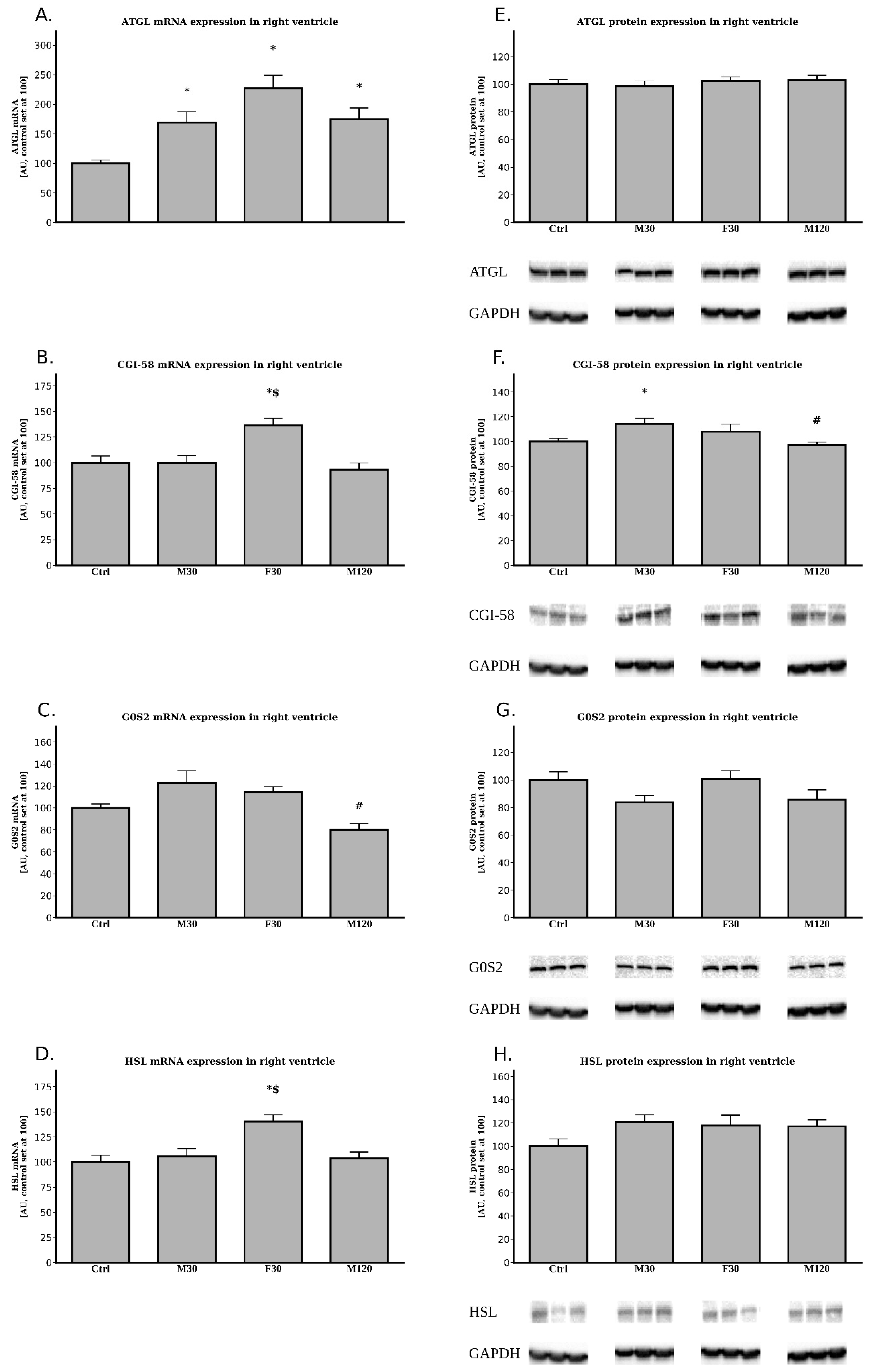
2.3.1. The Expression of ATGL, CGI-58, G0S2 and HSL at the Post-Transcript (mRNA) Level
2.3.2. The Expression of ATGL, CGI-58, G0S2 and HSL at the Protein Level
2.3.3. The Expression of PGC-1α at the Post-Transcript (mRNA) Level
2.3.4. The Expression of PGC-1α at the Protein Level
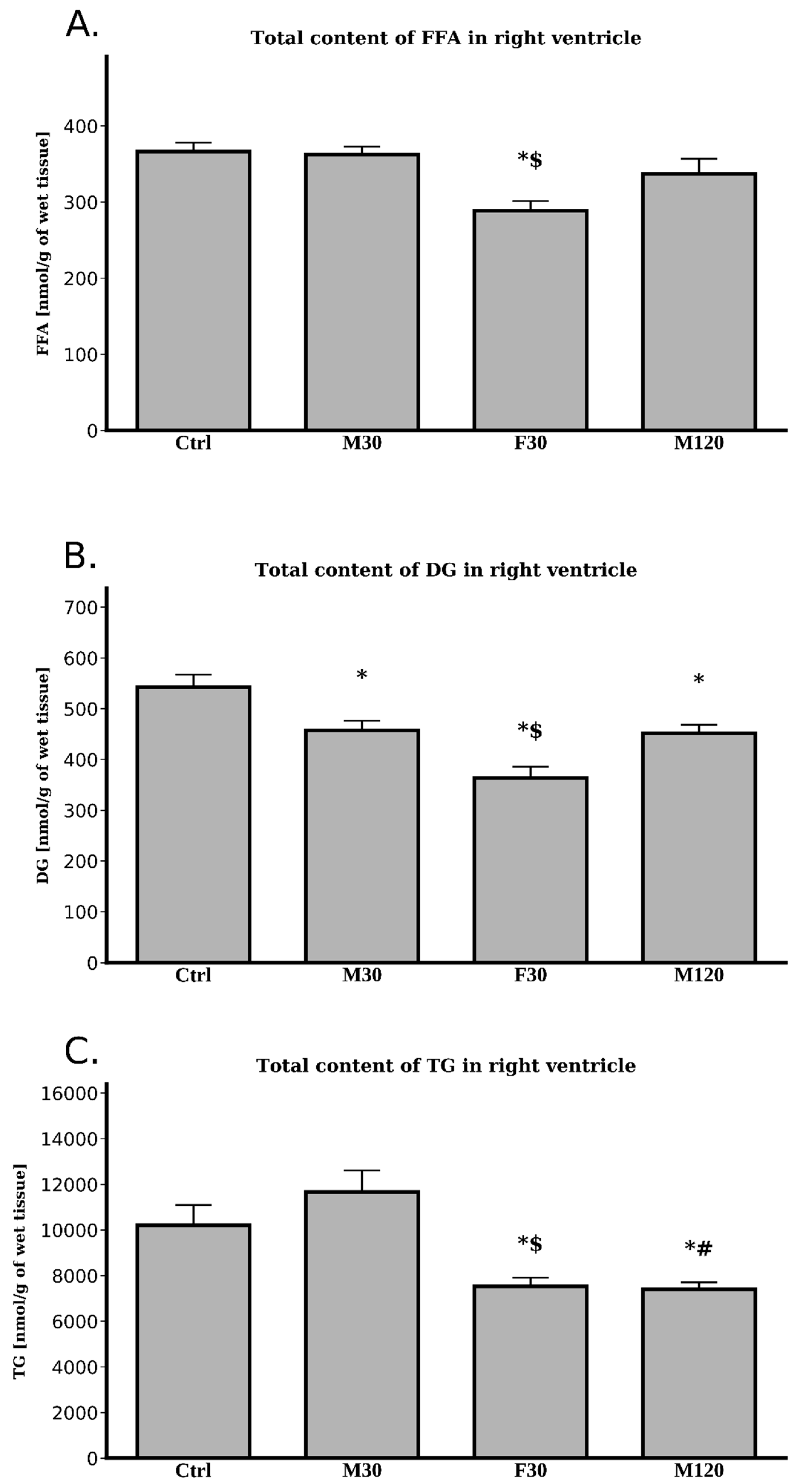
2.3.5. Lipids Content
Free Fatty Acids (FFA)
Diacylglycerols (DG)
Triacylglycerols (TG)
2.3.6. Total Lipolytic Activity
3. Discussion
3.1. The Left Ventricle
3.2. The Right Ventricle
4. Material and Methods
4.1. Animal Experiments
- Control (group designation: Ctrl),
- Moderately intense run on an electrically driven treadmill with the speed of 18 m/min for 30 min (group designation: M30),
- Fast (high intensity) run on an electrically driven treadmill with the speed of 28 m/min for 30 min (group designation: F30),
- Moderately intense run on an electrically driven treadmill with the speed of 18 m/min for 120 min (group designation: M120).
4.2. Lipid Content
4.3. Total Lipolytic Activity
4.4. Quantitative Real-Time PCR Analysis
- Cyclophilin A—F:5′-TGTCTCTTTTCGCCGCTTGCTG-3′; R:5′-CACCACCCTGGCACATGAATCC-3′;
- ATGL—F:5′-CCCTGACTCGAGTTTCGGAT-3′; R:5′-CACATAGCGCACCCCTTGAA-3′;
- G0S2—F:5′-TGACCTCCTTCAGCGAGTG-3′; R:5′-TCGGGACTTCTGCGTCATC-3′;
- CGI-58—F:5′-AACCCCAAGTGGTGAGACAG-3′; R:5′-GCGCCGAAGATGACTGAAAC-3′;
- HSL –F:5′-AATGACACAGTCGCTGGTGGCG-3′; R:5′-TGCCACACCCAAGAGCTGACCT-3′
- PGC1α—F: 5′-ACAGACACCGCACACATCGC-3′; R: 5′-GCTTCATAGCTGTCATACCTGGGC-3′;
4.5. Immunoblotting
4.6. Statistics
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| FFA | Free fatty acids |
| DG | Diacylglycerol |
| TG | Triacylglycerol |
| ATGL | Adipose triglyceride lipase |
| CGI-58 | Comparative gene identification-58 |
| G0S2 | G0/G1 switch gene 2 |
| HSL | Hormone sensitive lipase |
| Ctrl | Control group |
| M30 | Moderately intense run on an electrically driven treadmill with the speed of 18 m/min for 30 min |
| F30 | Fast (high intensity) run on an electrically driven treadmill with the speed of 28 m/min for 30 min |
| M120 | Moderately intense run on an electrically driven treadmill with the speed of 18 m/min for 120 min |
| PGC-1α | Peroxisome proliferator-activated receptor gamma coactivator 1-alpha |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid | Ctrl | M30 | F30 | M120 |
|---|---|---|---|---|
| Myristic acid (14:0) | 73.73 ± 3.207 | 88.98 ± 3.537 * | 72.93 ± 3.716 $ | 150.81 ± 17.922 *# |
| Palmitic acid (16:0) | 110.58 ± 2.295 | 117.76 ± 3.104 | 143.85 ± 12.956 | 132.99 ± 8.145 |
| Palmitooleic acid (16:1) | 6.93 ± 2.453 | 4.24 ± 0.176 | 5.62 ± 0.656 | 5.82 ± 0.559 |
| Stearic acid (18:0) | 63.12 ± 3.463 | 79.13 ± 2.096 * | 75.44 ± 8.094 | 84.84 ± 6.071 * |
| Oleic acid (18:1n9c) | 20.16 ± 0.793 | 16.82 ± 1.119 * | 24.32 ± 3.253 | 14.45 ± 1.145 * |
| Linoleic acid (18:2n6c) | 16.32 ± 1.148 | 6.07 ± 0.664 * | 9.59 ± 0.449 *$ | 11.65 ± 3.085 |
| Arachidic acid (20:0) | 5.01 ± 1.437 | 4.33 ± 0.203 | 2.65 ± 0.382 *$ | 5.97 ± 0.593 |
| Linolenic acid (18:9n3) | 2.7 ± 0.877 | 1.81 ± 0.103 | 1.45 ± 0.069 *$ | 2.81 ± 0.357 # |
| Behenic acid (22:0) | 2.52 ± 0.555 | 2.93 ± 0.108 * | 2.23 ± 0.775 $ | 4.03 ± 0.439 *# |
| Arachidonic acid (20:4n6) | 7.09 ± 0.616 | 3.83 ± 0.393 * | 5.82 ± 0.367 $ | 6.82 ± 1.692 |
| Lignoceric acid (24:0) | 3.83 ± 1.188 | 2.96 ± 0.128 | 2.96 ± 1.089 | 4.31 ± 0.419 |
| Eicosapentaenoic acid (20:5n3) | 0.57 ± 0.1 | 1.08 ± 0.116 * | 0.69 ± 0.127 | 1.59 ± 0.181 * |
| Nervonic acid (24:1) | 1.68 ± 0.736 | 0.73 ± 0.131 | 0.36 ± 0.133 * | 1.33 ± 0.168 # |
| Docosahexaenoic acid (22:6n3) | 3.03 ± 1.28 | 0.69 ± 0.199 * | 0.88 ± 0.168 * | 2.6 ± 0.433 # |
| UNSAT | 58.49 ± 6.877 | 35.26 ± 1.964 * | 48.74 ± 4.071 | 47.08 ± 7.16 *# |
| SAT | 258.79 ± 5.729 | 296.1 ± 5.113 * | 300.06 ± 20.175 | 382.95 ± 31.451 |
| Total | 317.28 ± 4.574 | 331.37 ± 6.756 | 348.8 ± 23.786 | 430.03 ± 38.524 |
| Fatty Acid | Ctrl | M30 | F30 | M120 |
|---|---|---|---|---|
| Myristic acid (14:0) | 52.85 ± 2.471 | 71.97 ± 19.535 | 67.05 ± 7.065 | 34.53 ± 1.926 * |
| Palmitic acid (16:0) | 163.62 ± 6.159 | 147.61 ± 12.37 | 157.98 ± 9.273 | 121.31 ± 4.949 * |
| Palmitooleic acid (16:1) | 3.98 ± 0.224 | 3.55 ± 0.284 | 4.28 ± 0.357 | 3.03 ± 0.09 * |
| Stearic acid (18:0) | 76.08 ± 1.331 | 93.12 ± 7.598 | 63.43 ± 4.014 *$ | 58.64 ± 2.026 *# |
| Oleic acid (18:1n9c) | 22.02 ± 1.693 | 16.03 ± 0.338 * | 18.0 ± 1.62 | 13.02 ± 0.941 *# |
| Linoleic acid (18:2n6c) | 41.44 ± 2.42 | 32.64 ± 1.918 | 26.3 ± 1.027 *$ | 26.94 ± 2.769 * |
| Arachidic acid (20:0) | 2.63 ± 0.357 | 5.39 ± 0.25 * | 3.14 ± 0.328 $ | 1.83 ± 0.147 *# |
| Linolenic acid (18:9n3) | 3.03 ± 0.317 | 2.5 ± 0.074 | 2.27 ± 0.082 | 2.02 ± 0.143 *# |
| Behenic acid (22:0) | 1.76 ± 0.109 | 3.19 ± 0.17 * | 2.3 ± 0.22 *$ | 1.26 ± 0.092 *# |
| Arachidonic acid (20:4n6) | 16.36 ± 0.468 | 17.14 ± 0.729 | 12.08 ± 0.845 *$ | 13.86 ± 1.332 |
| Lignoceric acid (24:0) | 2.73 ± 0.204 | 2.06 ± 0.186 | 2.27 ± 0.235 | 1.59 ± 0.204 * |
| Eicosapentaenoic acid (20:5n3) | 1.31 ± 0.117 | 0.91 ± 0.104 | 0.94 ± 0.144 | 0.9 ± 0.045 |
| Nervonic acid (24:1) | 0.91 ± 0.077 | 0.73 ± 0.087 | 0.61 ± 0.13 | 0.71 ± 0.058 |
| Docosahexaenoic acid (22:6n3) | 4.13 ± 0.412 | 3.64 ± 0.437 | 2.75 ± 0.378 | 3.91 ± 0.356 |
| UNSAT | 93.19 ± 4.909 | 77.13 ± 3.0 * | 67.23 ± 2.302 *$ | 64.38 ± 4.712 *# |
| SAT | 299.68 ± 7.128 | 323.33 ± 39.034 | 296.17 ± 18.268 | 219.15 ± 8.204 *# |
| Total | 392.87 ± 8.607 | 400.46 ± 37.197 | 363.41 ± 19.126 | 283.54 ± 8.619 *# |
| Fatty Acid | Ctrl | M30 | F30 | M120 |
|---|---|---|---|---|
| Myristic acid (14:0) | 91.25 ± 9.6 | 101.23 ± 11.452 | 93.31 ± 8.865 | 90.14 ± 5.451 |
| Palmitic acid (16:0) | 1808.44 ± 142.466 | 1887.52 ± 31.117 | 1928.92 ± 106.223 | 2046.47 ± 188.593 |
| Palmitooleic acid (16:1) | 109.65 ± 22.486 | 130.03 ± 9.572 | 144.11 ± 13.316 | 134.5 ± 16.395 |
| Stearic acid (18:0) | 331.59 ± 18.134 | 363.98 ± 15.939 | 369.92 ± 17.223 | 406.06 ± 26.889 |
| Oleic acid (18:1n9c) | 994.35 ± 102.385 | 972.33 ± 36.711 | 1014.15 ± 75.673 | 988.29 ± 109.766 |
| Linoleic acid (18:2n6c) | 1600.33 ± 115.5 | 1523.26 ± 74.325 | 1525.85 ± 71.725 | 1712.54 ± 179.137 |
| Arachidic acid (20:0) | 12.0 ± 0.482 | 27.75 ± 6.178 * | 14.92 ± 2.395 $ | 9.49 ± 0.898 # |
| Linolenic acid (18:9n3) | 116.35 ± 9.016 | 133.55 ± 8.768 | 133.32 ± 7.231 | 143.92 ± 15.362 |
| Behenic acid (22:0) | 16.29 ± 0.82 | 20.92 ± 1.407 * | 24.63 ± 2.671 * | 21.36 ± 2.017 |
| Arachidonic acid (20:4n6) | 103.77 ± 7.942 | 173.8 ± 11.261 * | 166.24 ± 6.223 * | 242.79 ± 17.27 *# |
| Lignoceric acid (24:0) | 7.7 ± 0.816 | 8.29 ± 0.698 | 8.37 ± 0.611 | 7.25 ± 0.806 |
| Eicosapentaenoic acid (20:5n3) | 6.65 ± 0.589 | 9.53 ± 0.789 * | 9.67 ± 0.398 * | 10.63 ± 0.658 * |
| Nervonic acid (24:1) | 5.11 ± 0.514 | 4.95 ± 0.403 | 5.43 ± 0.343 | 4.54 ± 0.444 |
| Docosahexaenoic acid (22:6n3) | 70.94 ± 10.027 | 135.54 ± 13.015 * | 109.92 ± 8.28 * | 135.52 ± 6.002 * |
| UNSAT | 3007.15 ± 235.749 | 3082.98 ± 133.149 | 3108.68 ± 159.705 | 3372.73 ± 335.539 |
| SAT | 2267.27 ± 167.302 | 2409.69 ± 56.829 | 2440.06 ± 131.837 | 2580.77 ± 222.949 |
| Total | 5274.42 ± 397.857 | 5492.68 ± 177.461 | 5548.75 ± 287.944 | 5953.5 ± 553.351 |
| Fatty Acid | Ctrl | M30 | F30 | M120 |
|---|---|---|---|---|
| Myristic acid (14:0) | 75.4 ± 3.026 | 77.91 ± 3.399 | 64.87 ± 2.872 #$ | 77.45 ± 2.974 |
| Palmitic acid (16:0) | 147.82 ± 9.791 | 143.17 ± 8.684 | 118.88 ± 6.792 | 139.47 ± 9.687 |
| Palmitooleic acid (16:1) | 5.99 ± 0.284 | 5.61 ± 0.369 | 4.64 ± 0.209 *# | 5.98 ± 0.228 |
| Stearic acid (18:0) | 80.51 ± 4.04 | 88.86 ± 4.604 | 62.28 ± 2.188 *$ | 66.03 ± 4.741 # |
| Oleic acid (18:1n9c) | 21.55 ± 2.086 | 19.3 ± 1.605 | 15.52 ± 0.652 | 16.69 ± 1.196 |
| Linoleic acid (18:2n6c) | 15.97 ± 2.466 | 10.93 ± 0.936 | 9.35 ± 0.792 | 13.04 ± 1.453 |
| Arachidic acid (20:0) | 2.94 ± 0.124 | 3.0 ± 0.27 | 2.24 ± 0.154 * | 2.54 ± 0.215 |
| Linolenic acid (18:9n3) | 1.87 ± 0.167 | 1.68 ± 0.104 | 1.33 ± 0.082 *#$ | 1.7 ± 0.116 |
| Behenic acid (22:0) | 1.93 ± 0.082 | 2.01 ± 0.204 | 1.39 ± 0.087 *$ | 1.61 ± 0.046 * |
| Arachidonic acid (20:4n6) | 6.63 ± 0.871 | 4.7 ± 0.384 | 4.0 ± 0.362 *# | 7.12 ± 0.932 |
| Lignoceric acid (24:0) | 2.36 ± 0.12 | 2.47 ± 0.201 | 2.02 ± 0.164 | 2.21 ± 0.104 |
| Eicosapentaenoic acid (20:5n3) | 1.11 ± 0.116 | 0.83 ± 0.048 | 0.72 ± 0.043 *# | 1.02 ± 0.083 |
| Nervonic acid (24:1) | 0.87 ± 0.096 | 0.66 ± 0.123 | 0.6 ± 0.044 | 0.75 ± 0.063 |
| Docosahexaenoic acid (22:6n3) | 1.62 ± 0.143 | 1.18 ± 0.105 | 0.97 ± 0.083 * | 1.34 ± 0.117 |
| UNSAT | 55.62 ± 4.481 | 44.89 ± 2.936 | 37.13 ± 1.83 * | 47.64 ± 3.515 |
| SAT | 310.96 ± 9.654 | 317.41 ± 9.761 | 251.68 ± 11.806 *$ | 289.31 ± 17.013 |
| Total | 366.57 ± 11.443 | 362.3 ± 10.663 | 288.81 ± 12.596 *$ | 336.94 ± 20.093 |
| Fatty Acid | Ctrl | M30 | F30 | M120 |
|---|---|---|---|---|
| Myristic acid (14:0) | 81.94 ± 3.76 | 46.84 ± 3.859 * | 29.95 ± 3.777 *$ | 63.62 ± 4.748 *# |
| Palmitic acid (16:0) | 239.47 ± 9.473 | 230.9 ± 16.085 | 180.91 ± 14.625 * | 216.21 ± 14.375 |
| Palmitooleic acid (16:1) | 10.24 ± 1.008 | 7.47 ± 0.495 * | 5.54 ± 0.536 *$ | 7.06 ± 0.415 * |
| Stearic acid (18:0) | 84.13 ± 5.611 | 85.6 ± 1.934 | 67.43 ± 4.071 $ | 76.48 ± 4.709 |
| Oleic acid (18:1n9c) | 34.42 ± 2.822 | 20.26 ± 1.72 * | 19.08 ± 1.145 * | 19.16 ± 1.056 * |
| Linoleic acid (18:2n6c) | 47.3 ± 4.943 | 30.07 ± 2.753 * | 29.29 ± 2.026 * | 31.04 ± 3.235 * |
| Arachidic acid (20:0) | 6.02 ± 1.024 | 5.45 ± 0.71 | 4.11 ± 0.326 | 5.17 ± 0.358 |
| Linolenic acid (18:9n3) | 3.49 ± 0.204 | 2.64 ± 0.349 | 2.42 ± 0.157 * | 2.63 ± 0.078 * |
| Behenic acid (22:0) | 4.25 ± 0.778 | 3.65 ± 0.427 | 2.84 ± 0.218 | 3.8 ± 0.249 |
| Arachidonic acid (20:4n6) | 20.72 ± 2.198 | 15.25 ± 1.269 | 12.67 ± 1.213 * | 15.64 ± 1.746 |
| Lignoceric acid (24:0) | 4.48 ± 0.272 | 4.84 ± 0.84 | 3.69 ± 0.443 | 4.34 ± 0.717 |
| Eicosapentaenoic acid (20:5n3) | 1.39 ± 0.185 | 0.89 ± 0.184 | 0.86 ± 0.068 | 1.17 ± 0.129 |
| Nervonic acid (24:1) | 1.18 ± 0.081 | 0.91 ± 0.098 | 0.94 ± 0.088 | 1.08 ± 0.085 |
| Docosahexaenoic acid (22:6n3) | 3.89 ± 0.712 | 2.54 ± 0.325 | 3.26 ± 0.4 | 4.23 ± 0.317 # |
| UNSAT | 122.63 ± 8.393 | 80.03 ± 5.398 * | 74.06 ± 4.37 * | 82.01 ± 4.825 * |
| SAT | 420.29 ± 18.549 | 377.28 ± 19.805 | 288.92 ± 20.804 *$ | 369.62 ± 20.365 |
| Total | 542.92 ± 24.155 | 457.31 ± 18.775 * | 362.98 ± 22.621 *$ | 451.63 ± 16.829 * |
| Fatty Acid | Ctrl | M30 | F30 | M120 |
|---|---|---|---|---|
| Myristic acid (14:0) | 233.01 ± 31.196 | 257.8 ± 36.272 | 158.17 ± 12.991 $ | 136.74 ± 14.038 *# |
| Palmitic acid (16:0) | 3288.24 ± 258.05 | 3676.25 ± 321.464 | 2578.81 ± 153.119 $ | 2481.14 ± 135.093 *# |
| Palmitooleic acid (16:1) | 386.33 ± 83.149 | 582.23 ± 113.805 | 188.27 ± 24.718 *$ | 203.71 ± 18.59 # |
| Stearic acid (18:0) | 580.98 ± 45.522 | 633.8 ± 36.805 | 453.99 ± 17.257 *$ | 513.46 ± 27.152 # |
| Oleic acid (18:1n9c) | 2206.31 ± 256.998 | 2597.64 ± 265.536 | 1505.47 ± 108.684 *$ | 1279.92 ± 74.866 *# |
| Linoleic acid (18:2n6c) | 2934.12 ± 298.753 | 3193.91 ± 211.725 | 2077.96 ± 86.963 *$ | 2091.18 ± 78.179 *# |
| Arachidic acid (20:0) | 26.44 ± 0.99 | 27.59 ± 1.28 | 24.55 ± 0.945 | 22.19 ± 0.747 *# |
| Linolenic acid (18:9n3) | 250.26 ± 28.214 | 261.84 ± 28.113 | 178.56 ± 9.399 | 166.05 ± 5.742 * |
| Behenic acid (22:0) | 33.63 ± 1.373 | 36.53 ± 2.833 | 35.24 ± 1.501 | 37.26 ± 1.163 |
| Arachidonic acid (20:4n6) | 156.97 ± 13.47 | 213.54 ± 9.911 * | 178.9 ± 5.941 $ | 269.42 ± 12.816 *# |
| Lignoceric acid (24:0) | 12.82 ± 0.678 | 11.16 ± 0.653 | 11.82 ± 1.071 | 12.44 ± 1.156 |
| Eicosapentaenoic acid (20:5n3) | 8.43 ± 0.696 | 12.92 ± 0.956 * | 10.79 ± 0.614 * | 14.59 ± 1.05 * |
| Nervonic acid (24:1) | 5.18 ± 0.193 | 6.06 ± 0.232 * | 5.9 ± 0.52 | 4.86 ± 0.349 # |
| Docosahexaenoic acid (22:6n3) | 90.19 ± 13.549 | 162.94 ± 17.452 * | 126.1 ± 9.079 | 174.16 ± 5.853 * |
| UNSAT | 6037.78 ± 563.3 | 7031.1 ± 556.859 | 4271.95 ± 198.241 *$ | 4203.89 ± 160.521 *# |
| SAT | 4175.12 ± 326.71 | 4643.13 ± 391.547 | 3262.58 ± 184.053 *$ | 3203.22 ± 158.731 *# |
| Total | 10212.9 ± 881.065 | 11674.23 ± 935.1 | 7534.53 ± 371.363 *$ | 7407.11 ± 298.955 *# |
References
- Choromańska, B.; Myśliwiec, P.; Razak Hady, H.; Dadan, J.; Myśliwiec, H.; Chabowski, A.; Mikłosz, A. Metabolic syndrome is associated with ceramide accumulation in visceral adipose tissue of women with morbid obesity. Obesity 2019, 27, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Mikłosz, A.; Łukaszuk, B.; Żendzian-Piotrowska, M.; Brańska-Januszewska, J.; Ostrowska, H.; Chabowski, A. Challenging of as160/tbc1d4 alters intracellular lipid milieu in l6 myotubes incubated with palmitate. J. Cell. Physiol. 2017, 232, 2373–2386. [Google Scholar] [CrossRef]
- Meex, R.C.R.; Hoy, A.J.; Mason, R.M.; Martin, S.D.; McGee, S.L.; Bruce, C.R.; Watt, M.J. ATGL-mediated triglyceride turnover and the regulation of mitochondrial capacity in skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2015, 308, E960–E970. [Google Scholar] [CrossRef] [Green Version]
- Banke, N.H.; Wende, A.R.; Leone, T.C.; O’Donnell, J.M.; Abel, E.D.; Kelly, D.P.; Lewandowski, E.D. Preferential oxidation of triacylglyceride-derived fatty acids in heart is augmented by the nuclear receptor pparalpha. Circ. Res. 2010, 107, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Haemmerle, G.; Lass, A.; Zimmermann, R.; Gorkiewicz, G.; Meyer, C.; Rozman, J.; Heldmaier, G.; Maier, R.; Theussl, C.; Eder, S.; et al. Defective lipolysis and altered energy metabolism in mice lacking adipose triglyceride lipase. Science 2006, 312, 734–737. [Google Scholar] [CrossRef]
- Paul, A.; Chan, L.; Bickel, P.E. The pat family of lipid droplet proteins in heart and vascular cells. Curr. Hypertens. Rep. 2008, 10, 461–466. [Google Scholar] [CrossRef]
- Kienesberger, P.C.; Pulinilkunnil, T.; Sung, M.M.Y.; Nagendran, J.; Haemmerle, G.; Kershaw, E.E.; Young, M.E.; Light, P.E.; Oudit, G.Y.; Zechner, R.; et al. Myocardial atgl overexpression decreases the reliance on fatty acid oxidation and protects against pressure overload-induced cardiac dysfunction. Mol. Cell. Biol. 2012, 32, 740–750. [Google Scholar] [CrossRef]
- Nielsen, T.S.; Jessen, N.; Jørgensen, J.O.L.; Møller, N.; Lund, S. Dissecting adipose tissue lipolysis: Molecular regulation and implications for metabolic disease. J. Mol. Endocrinol. 2014, 52, R199–R222. [Google Scholar] [CrossRef]
- Lass, A.; Zimmermann, R.; Haemmerle, G.; Riederer, M.; Schoiswohl, G.; Schweiger, M.; Kienesberger, P.; Strauss, J.G.; Gorkiewicz, G.; Zechner, R. Adipose triglyceride lipase-mediated lipolysis of cellular fat stores is activated by cgi-58 and defective in chanarin-dorfman syndrome. Cell Metab. 2006, 3, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Watt, M.J.; Hoy, A.J. Lipid metabolism in skeletal muscle: Generation of adaptive and maladaptive intracellular signals for cellular function. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E1315–E1328. [Google Scholar] [CrossRef]
- Ahmadian, M.; Duncan, R.E.; Varady, K.A.; Frasson, D.; Hellerstein, M.K.; Birkenfeld, A.L.; Samuel, V.T.; Shulman, G.I.; Wang, Y.; Kang, C.; et al. Adipose overexpression of desnutrin promotes fatty acid use and attenuates diet-induced obesity. Diabetes 2009, 58, 855–866. [Google Scholar] [CrossRef] [PubMed]
- Fischer, J.; Lefèvre, C.; Morava, E.; Mussini, J.-M.; Laforêt, P.; Negre-Salvayre, A.; Lathrop, M.; Salvayre, R. The gene encoding adipose triglyceride lipase (pnpla2) is mutated in neutral lipid storage disease with myopathy. Nat. Genet. 2007, 39, 28–30. [Google Scholar] [CrossRef] [PubMed]
- Ghanbari-Niaki, A.; Ghanbari-Abarghooi, S.; Rahbarizadeh, F.; Zare-Kookandeh, N.; Gholizadeh, M.; Roudbari, F.; Zare-Kookandeh, A. Heart abca1 and ppar- α genes expression responses in male rats: Effects of high intensity treadmill running training and aqueous extraction of black crataegus-pentaegyna. Res. Cardiovasc. Med. 2013, 2, 153–159. [Google Scholar] [CrossRef]
- Fröberg, S.O. Effect of acute exercise on tissue lipids in rats. Metab. Clin. Exp. 1971, 20, 714–720. [Google Scholar] [CrossRef]
- Schrauwen-Hinderling, V.B.; Meex, R.C.R.; Hesselink, M.K.C.; van de Weijer, T.; Leiner, T.; Schär, M.; Lamb, H.J.; Wildberger, J.E.; Glatz, J.F.C.; Schrauwen, P.; et al. Cardiac lipid content is unresponsive to a physical activity training intervention in type 2 diabetic patients, despite improved ejection fraction. Cardiovasc. Diabetol. 2011, 10, 47. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Yu, S.; Khan, R.S.; Ables, G.P.; Bharadwaj, K.G.; Hu, Y.; Huggins, L.A.; Eriksson, J.W.; Buckett, L.K.; Turnbull, A.V.; et al. DGAT1 deficiency decreases ppar expression and does not lead to lipotoxicity in cardiac and skeletal muscle. J. Lipid Res. 2011, 52, 732–744. [Google Scholar] [CrossRef] [PubMed]
- Dobrzyn, P.; Pyrkowska, A.; Duda, M.K.; Bednarski, T.; Maczewski, M.; Langfort, J.; Dobrzyn, A. Expression of lipogenic genes is upregulated in the heart with exercise training-induced but not pressure overload-induced left ventricular hypertrophy. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E1348–E1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sai, E.; Shimada, K.; Yokoyama, T.; Sato, S.; Miyazaki, T.; Hiki, M.; Tamura, Y.; Aoki, S.; Watada, H.; Kawamori, R.; et al. Association between myocardial triglyceride content and cardiac function in healthy subjects and endurance athletes. PLoS ONE 2013, 8, e61604. [Google Scholar] [CrossRef]
- Wojcik, B.; Miklosz, A.; Zabielski, P.; Chabowski, A.; Gorski, J. Effect of tachycardia on mRNA and protein expression of the principal components of the lipolytic system in the rat’s heart ventricles. J. Physiol. Pharmacol. 2017, 68, 731–736. [Google Scholar]
- Shepherd, R.E.; Gollnick, P.D. Oxygen uptake of rats at different work intensities. Pflug. Arch. 1976, 362, 219–222. [Google Scholar] [CrossRef]
- Foryst-Ludwig, A.; Kreissl, M.C.; Benz, V.; Brix, S.; Smeir, E.; Ban, Z.; Januszewicz, E.; Salatzki, J.; Grune, J.; Schwanstecher, A.-K.; et al. Adipose tissue lipolysis promotes exercise-induced cardiac hypertrophy involving the lipokine c16:1n7-palmitoleate. J. Biol. Chem. 2015, 290, 23603–23615. [Google Scholar] [CrossRef]
- Gertz, E.W.; Wisneski, J.A.; Stanley, W.C.; Neese, R.A. Myocardial substrate utilization during exercise in humans. dual carbon-labeled carbohydrate isotope experiments. J. Clin. Investig. 1988, 82, 2017–2025. [Google Scholar] [CrossRef]
- Kolwicz, S.C. An “Exercise” in cardiac metabolism. Front. Cardiovasc. Med. 2018, 5, 66. [Google Scholar] [CrossRef] [PubMed]
- Lawler, J.M.; Powers, S.K.; Hammeren, J.; Martin, A.D. Oxygen cost of treadmill running in 24-month-old fischer-344 rats. Med. Sci. Sports Exerc. 1993, 25, 1259–1264. [Google Scholar] [CrossRef]
- Stankiewicz-Choroszucha, B.; Górski, J. Effect of substrate supply and beta-adrenergic blockade on heart glycogen and triglyceride utilization during exercise in the rat. Eur. J. Appl. Physiol. Occup. Physiol. 1980, 43, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Nagy, H.M.; Paar, M.; Heier, C.; Moustafa, T.; Hofer, P.; Haemmerle, G.; Lass, A.; Zechner, R.; Oberer, M.; Zimmermann, R. Adipose triglyceride lipase activity is inhibited by long-chain acyl-coenzyme a. Biochim. Biophys. Acta 2014, 1841, 588–594. [Google Scholar] [CrossRef]
- Saddik, M.; Lopaschuk, G.D. Myocardial triglyceride turnover and contribution to energy substrate utilization in isolated working rat hearts. J. Biol. Chem. 1991, 266, 8162–8170. [Google Scholar]
- Boden, G.; Lebed, B.; Schatz, M.; Homko, C.; Lemieux, S. Effects of acute changes of plasma free fatty acids on intramyocellular fat content and insulin resistance in healthy subjects. Diabetes 2001, 50, 1612–1617. [Google Scholar] [CrossRef] [PubMed]
- Riehle, C.; Wende, A.R.; Zhu, Y.; Oliveira, K.J.; Pereira, R.O.; Jaishy, B.P.; Bevins, J.; Valdez, S.; Noh, J.; Kim, B.J.; et al. Insulin receptor substrates are essential for the bioenergetic and hypertrophic response of the heart to exercise training. Mol. Cell. Biol. 2014, 34, 3450–3460. [Google Scholar] [CrossRef]
- Monleon, D.; Garcia-Valles, R.; Morales, J.M.; Brioche, T.; Olaso-Gonzalez, G.; Lopez-Grueso, R.; Gomez-Cabrera, M.C.; Viña, J. Metabolomic analysis of long-term spontaneous exercise in mice suggests increased lipolysis and altered glucose metabolism when animals are at rest. J. Appl. Physiol. 2014, 117, 1110–1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huijsman, E.; van de Par, C.; Economou, C.; van der Poel, C.; Lynch, G.S.; Schoiswohl, G.; Haemmerle, G.; Zechner, R.; Watt, M.J. Adipose triacylglycerol lipase deletion alters whole body energy metabolism and impairs exercise performance in mice. Am. J. Physiol. Endocrinol. Metab. 2009, 297, E505–E513. [Google Scholar] [CrossRef] [Green Version]
- Schoiswohl, G.; Schweiger, M.; Schreiber, R.; Gorkiewicz, G.; Preiss-Landl, K.; Taschler, U.; Zierler, K.A.; Radner, F.P.W.; Eichmann, T.O.; Kienesberger, P.C.; et al. Adipose triglyceride lipase plays a key role in the supply of the working muscle with fatty acids. J. Lipid Res. 2010, 51, 490–499. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, R.; Strauss, J.G.; Haemmerle, G.; Schoiswohl, G.; Birner-Gruenberger, R.; Riederer, M.; Lass, A.; Neuberger, G.; Eisenhaber, F.; Hermetter, A.; et al. Fat mobilization in adipose tissue is promoted by adipose triglyceride lipase. Science 2004, 306, 1383–1386. [Google Scholar] [CrossRef]
- Taschler, U.; Radner, F.P.W.; Heier, C.; Schreiber, R.; Schweiger, M.; Schoiswohl, G.; Preiss-Landl, K.; Jaeger, D.; Reiter, B.; Koefeler, H.C.; et al. Monoglyceride lipase deficiency in mice impairs lipolysis and attenuates diet-induced insulin resistance. J. Biol. Chem. 2011, 286, 17467–17477. [Google Scholar] [CrossRef]
- Haemmerle, G.; Zimmermann, R.; Strauss, J.G.; Kratky, D.; Riederer, M.; Knipping, G.; Zechner, R. Hormone-sensitive lipase deficiency in mice changes the plasma lipid profile by affecting the tissue-specific expression pattern of lipoprotein lipase in adipose tissue and muscle. J. Biol. Chem. 2002, 277, 12946–12952. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Lu, X.; Lombès, M.; Rha, G.B.; Chi, Y.-I.; Guerin, T.M.; Smart, E.J.; Liu, J. The g(0)/g(1) switch gene 2 regulates adipose lipolysis through association with adipose triglyceride lipase. Cell Metab. 2010, 11, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Gruber, A.; Cornaciu, I.; Lass, A.; Schweiger, M.; Poeschl, M.; Eder, C.; Kumari, M.; Schoiswohl, G.; Wolinski, H.; Kohlwein, S.D.; et al. The n-terminal region of comparative gene identification-58 (cgi-58) is important for lipid droplet binding and activation of adipose triglyceride lipase. J. Biol. Chem. 2010, 285, 12289–12298. [Google Scholar] [CrossRef]
- McArdle, W.D.; Katch, F.E.; Katch, V.L. Exercise Physiology: Nutrition, Energy, and Human Performance, 8th ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2014; Chapter 22; pp. 467–503. [Google Scholar]
- Crystal, G.J.; Pagel, P.S. Right ventricular perfusion: Physiology and clinical implications. Anesthesiology 2018, 128, 202–218. [Google Scholar] [CrossRef]
- Dai, Z.; Wu, F.; Yeung, E.W.; Li, Y. IGF-iec expression, regulation and biological function in different tissues. Growth Horm. Igf Res. 2010, 20, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Nawrocki, A.; Górski, J. Effect of plasma free fatty acid concentration on the content and composition of the free fatty acid fraction in rat skeletal muscles. Horm. Metab. Res. 2004, 36, 601–606. [Google Scholar] [CrossRef]
- Supruniuk, E.; Mikłosz, A.; Chabowski, A.; Łukaszuk, B. Dose- and time-dependent alterations in lipid metabolism after pharmacological pgc-1α activation in l6 myotubes. J. Cell. Physiol. 2019, 234, 11923–11941. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time rt-pcr. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Mikłosz, A.; Łukaszuk, B.; Żendzian-Piotrowska, M.; Kurek, K.; Chabowski, A. The effects of as160 modulation on fatty acid transporters expression and lipid profile in l6 myotubes. Cell. Physiol. Biochem. 2016, 38, 267–282. [Google Scholar] [CrossRef] [PubMed]






| Fatty Acid | Ctrl | M30 | F30 | M120 |
|---|---|---|---|---|
| Myristic acid (14:0) | 4.69 ± 0.51 | 2.92 ± 0.3 * | 7.15 ± 0.717 *$ | 10.02 ± 0.52 *# |
| Palmitic acid (16:0) | 52.06 ± 4.233 | 81.94 ± 6.083 * | 111.14 ± 3.421 *$ | 138.12 ± 5.074 *# |
| Palmitooleic acid (16:1) | 4.38 ± 0.717 | 9.75 ± 1.163 * | 16.41 ± 1.233 *$ | 19.49 ± 1.505 *# |
| Stearic acid (18:0) | 13.65 ± 0.596 | 18.18 ± 1.063 * | 19.42 ± 0.522* | 24.59 ± 0.906 *# |
| Oleic acid (18:1n9c) | 19.34 ± 1.98 | 34.08 ± 2.999 * | 44.35 ± 1.308 *$ | 44.79 ± 3.132 *# |
| Linoleic acid (18:2n6c) | 42.57 ± 4.44 | 67.27 ± 5.322 * | 83.77 ± 3.798 *$ | 99.7 ± 5.476 *# |
| Arachidic acid (20:0) | 0.37 ± 0.017 | 0.33 ± 0.015 | 0.32 ± 0.02 | 44.5 ± 9.838 |
| Linolenic acid (18:9n3) | 4.87 ± 0.617 | 9.14 ± 0.737 * | 11.38 ± 0.578 *$ | 10.33 ± 1.167 * |
| Behenic acid (22:0) | 0.31 ± 0.019 | 0.32 ± 0.019 | 0.33 ± 0.019 | 5.96 ± 1.25 *# |
| Arachidonic acid (20:4n6) | 6.28 ± 0.474 | 10.01 ± 0.644 * | 10.03 ± 0.267 * | 9.07 ± 0.965 * |
| Lignoceric acid (24:0) | 0.44 ± 0.026 | 0.44 ± 0.031 | 0.45 ± 0.051 | 4.72 ± 1.012 |
| Eicosapentaenoic acid (20:5n3) | 0.26 ± 0.026 | 0.64 ± 0.051 * | 0.66 ± 0.044 * | 0.72 ± 0.068 * |
| Nervonic acid (24:1) | 0.15 ± 0.011 | 0.16 ± 0.009 | 0.14 ± 0.009 | 0.44 ± 0.061 *# |
| Docosahexaenoic acid (22:6n3) | 1.46 ± 0.156 | 2.42 ± 0.2 * | 2.2 ± 0.133 * | 1.6 ± 0.192 # |
| SAT | 71.51 ± 5.299 | 104.14 ± 7.298 * | 138.81 ± 4.176 *$ | 227.92 ± 17.043 *# |
| UNSAT | 79.32 ± 8.008 | 133.47 ± 10.703 * | 168.95 ± 6.138 *$ | 186.14 ± 9.822 *# |
| Total | 150.83 ± 13.165 | 237.61 ± 17.776 * | 307.77 ± 9.524 *$ | 414.06 ± 14.435 *# |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mikłosz, A.; Łukaszuk, B.; Baranowski, M.; Chabowski, A.; Górski, J. Assessment of the Main Compounds of the Lipolytic System in Treadmill Running Rats: Different Response Patterns between the Right and Left Ventricle. Int. J. Mol. Sci. 2019, 20, 2556. https://doi.org/10.3390/ijms20102556
Mikłosz A, Łukaszuk B, Baranowski M, Chabowski A, Górski J. Assessment of the Main Compounds of the Lipolytic System in Treadmill Running Rats: Different Response Patterns between the Right and Left Ventricle. International Journal of Molecular Sciences. 2019; 20(10):2556. https://doi.org/10.3390/ijms20102556
Chicago/Turabian StyleMikłosz, Agnieszka, Bartłomiej Łukaszuk, Marcin Baranowski, Adrian Chabowski, and Jan Górski. 2019. "Assessment of the Main Compounds of the Lipolytic System in Treadmill Running Rats: Different Response Patterns between the Right and Left Ventricle" International Journal of Molecular Sciences 20, no. 10: 2556. https://doi.org/10.3390/ijms20102556