Somatostatin and the “Small-For-Size” Liver

 ,
,
Abstract
:1. Introduction
2. Changes Occurring in the “Small-For-Size” Liver
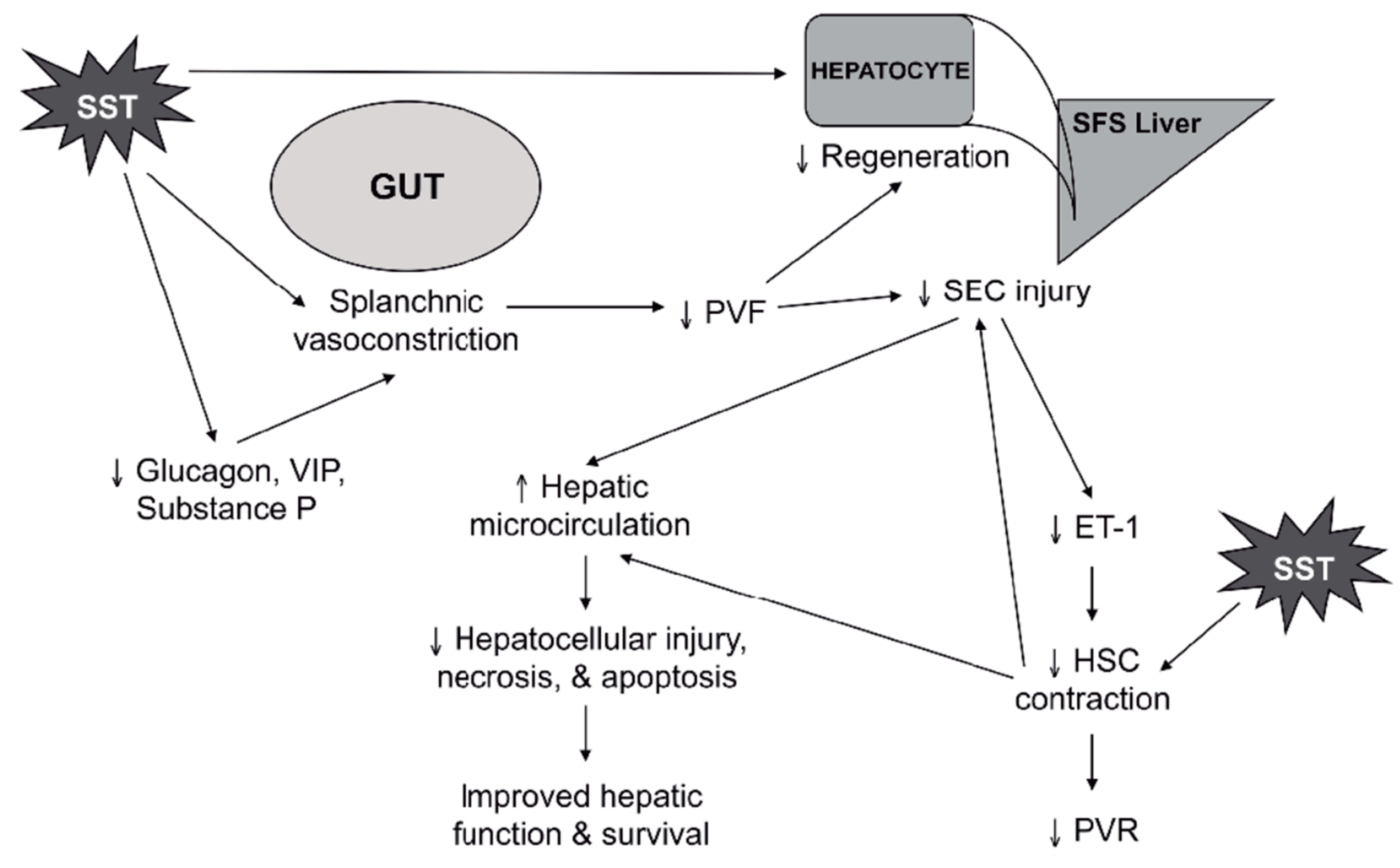
3. Somatostatin Mechanisms of Action
3.1. Splanchnic Vasculature
3.2. Liver
3.3. Potential Side Effects
4. Somatostatin and the “Small-For-Size” Liver In Vivo
4.1. Following Major Hepatectomy
4.2. Following Liver Transplantation
5. Summary and Future Directions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Cieslak, K.P.; Runge, J.H.; Heger, M.; Stoker, J.; Bennink, R.J.; van Gulik, T.M. New perspectives in the assessment of future remnant liver. Dig. Surg. 2014, 31, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Heaton, N. Small-for-size liver syndrome after auxiliary and split liver transplantation: Donor selection. Liver Transplant. 2003, 9, S26–S28. [Google Scholar] [CrossRef]
- Balzan, S.; Belghiti, J.; Farges, O.; Ogata, S.; Sauvanet, A.; Delefosse, D.; Durand, F. The “50-50 criteria” on postoperative day 5: An accurate predictor of liver failure and death after hepatectomy. Ann. Surg. 2005, 242, 824–828. [Google Scholar] [CrossRef]
- Van den Broek, M.A.; Olde Damink, S.W.; Dejong, C.H.; Lang, H.; Malago, M.; Jalan, R.; Saner, F.H. Liver failure after partial hepatic resection: Definition, pathophysiology, risk factors and treatment. Liver Int. 2008, 28, 767–780. [Google Scholar] [CrossRef] [PubMed]
- Rahbari, N.N.; Garden, O.J.; Padbury, R.; Brooke-Smith, M.; Crawford, M.; Adam, R.; Koch, M.; Makuuchi, M.; DeMatteo, R.P.; Christophi, C.; et al. Posthepatectomy liver failure: A definition and grading by the International Study Group of Liver Surgery (ISGLS). Surgery 2011, 149, 713–724. [Google Scholar] [CrossRef] [PubMed]
- Michalopoulos, G.K. Hepatostat: Liver regeneration and normal liver tissue maintenance. Hepatology 2017, 65, 1384–1392. [Google Scholar] [CrossRef]
- Álvarez-Sola, G.; Uriarte, I.; Latasa, M.U.; Jimenez, M.; Barcena-Varela, M.; Santamaria, E.; Urtasun, R.; Rodríguez-Ortigosa, C.; Prieto, J.; Berraondo, P.; et al. Bile acids, FGF15/19 and liver regeneration: From mechanisms to clinical applications. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 1326–1334. [Google Scholar] [CrossRef] [PubMed]
- Michalopoulos, G.K. Advances in liver regeneration. Expert Rev. Gastroenterol. Hepatol. 2014, 8, 897–907. [Google Scholar] [CrossRef]
- Taub, R. Liver regeneration: From myth to mechanism. Nat. Rev. Mol. Cell Biol. 2004, 5, 836–847. [Google Scholar] [CrossRef]
- Fausto, N.; Campbell, J.S.; Riehle, K.J. Liver regeneration. Hepatology 2006, 43 (Suppl. 1), S45–S53. [Google Scholar] [CrossRef]
- Michalopoulos, G.K. Liver regeneration. J. Cell. Physiol. 2007, 213, 286–300. [Google Scholar] [CrossRef]
- Matsubara, T.; Li, F.; Gonzalez, F.J. FXR signaling in the enterohepatic system. Mol. Cell. Endocrinol. 2013, 368, 17–19. [Google Scholar] [CrossRef]
- Uriarte, I.; Fernandez-Barrena, M.G.; Monte, M.J.; Latasa, M.U.; Chang, H.C.; Carotti, S.; Vespasiani-Gentilucci, U.; Morini, S.; Vicente, E.; Concepción, A.R.; et al. Identification of fibroblast growth factor 15 as a novel mediator of liver regeneration and its application in the prevention of post-resection liver failure in mice. Gut 2013, 62, 899–910. [Google Scholar] [CrossRef]
- Navasa, M.; Feu, F.; Garcia-Pagan, J.C.; Jimenez, W.; Llach, J.; Rimola, A.; Bosch, J.; Rodés, J. Hemodynamic and humoral changes after liver transplantation in patients with cirrhosis. Hepatology 1993, 17, 355–360. [Google Scholar] [CrossRef]
- Eguchi, S.; Yanaga, K.; Sugiyama, N.; Okudaira, S.; Furui, J.; Kanematsu, T. Relationship between portal venous flow and liver regeneration in patients after living donor right-lobe liver transplantation. Liver Transpl. 2003, 9, 547–551. [Google Scholar] [CrossRef]
- Maetani, Y.; Itoh, K.; Egawa, H.; Shibata, T.; Ametani, F.; Kubo, T.; Kiuchi, T.; Tanaka, K.; Konishi, J. Factors influencing liver regeneration following living-donor liver transplantation of the right hepatic lobe. Transplantation 2003, 75, 97–102. [Google Scholar] [CrossRef] [Green Version]
- Hessheimer, A.J.; Fondevila, C.; Taura, P.; Munoz, J.; Sanchez, O.; Fuster, J.; Rimola, A.; García-Valdecasas, J.C. Decompression of the portal bed and twice-baseline portal inflow are necessary for the functional recovery of a “small-for-size” graft. Ann. Surg. 2011, 253, 1201–1210. [Google Scholar] [CrossRef]
- Ku, Y.; Fukumoto, T.; Nishida, T.; Tominaga, M.; Maeda, I.; Kitagawa, T.; Takao, S.; Shiotani, M.; Tseng, A.; Kuroda, Y. Evidence that portal vein decompression improves survival of canine quarter orthotopic liver transplantation. Transplantation 1995, 59, 1388–1392. [Google Scholar] [CrossRef]
- Emond, J.C.; Renz, J.F.; Ferrell, L.D.; Rosenthal, P.; Lim, R.C.; Roberts, J.P.; Lake, J.R.; Ascher, N.L. Functional analysis of grafts from living donors. Implications for the treatment of older recipients. Ann. Surg. 1996, 224, 544–552. [Google Scholar] [CrossRef]
- Troisi, R.; Ricciardi, S.; Smeets, P.; Petrovic, M.; Van, M.G.; Colle, I.; van Vlierberghe, H.; de Hemptinne, B. Effects of hemi-portocaval shunts for inflow modulation on the outcome of small-for-size grafts in living donor liver transplantation. Am. J. Transplant. 2005, 5, 1397–1404. [Google Scholar] [CrossRef]
- Fondevila, C.; Hessheimer, A.J.; Taura, P.; Sanchez, O.; Calatayud, D.; de Riva, N.; Muñoz, J.; Fuster, J.; Rimola, A.; García-Valdecasas, J.C. Portal hyperperfusion: Mechanism of injury and stimulus for regeneration in porcine small-for-size transplantation. Liver Transplant. 2010, 16, 364–374. [Google Scholar] [CrossRef]
- Hessheimer, A.J.; Escobar, B.; Munoz, J.; Flores, E.; Gracia-Sancho, J.; Taura, P.; Taurá, P.; Fuster, J.; Rimola, A.; García-Valdecasas, J.C.; Fondevila, C. Somatostatin Therapy Protects Porcine Livers in Small-for-Size Liver Transplantation. Am. J. Transplant. 2014, 14, 1806–1816. [Google Scholar] [CrossRef]
- Demetris, A.J.; Kelly, D.M.; Eghtesad, B.; Fontes, P.; Wallis, M.J.; Tom, K.; Tan, H.P.; Shaw-Stiffel, T.; Boig, L.; Novelli, P.; et al. Pathophysiologic observations and histopathologic recognition of the portal hyperperfusion or small-for-size syndrome. Am. J. Surg. Pathol. 2006, 30, 986–993. [Google Scholar] [CrossRef]
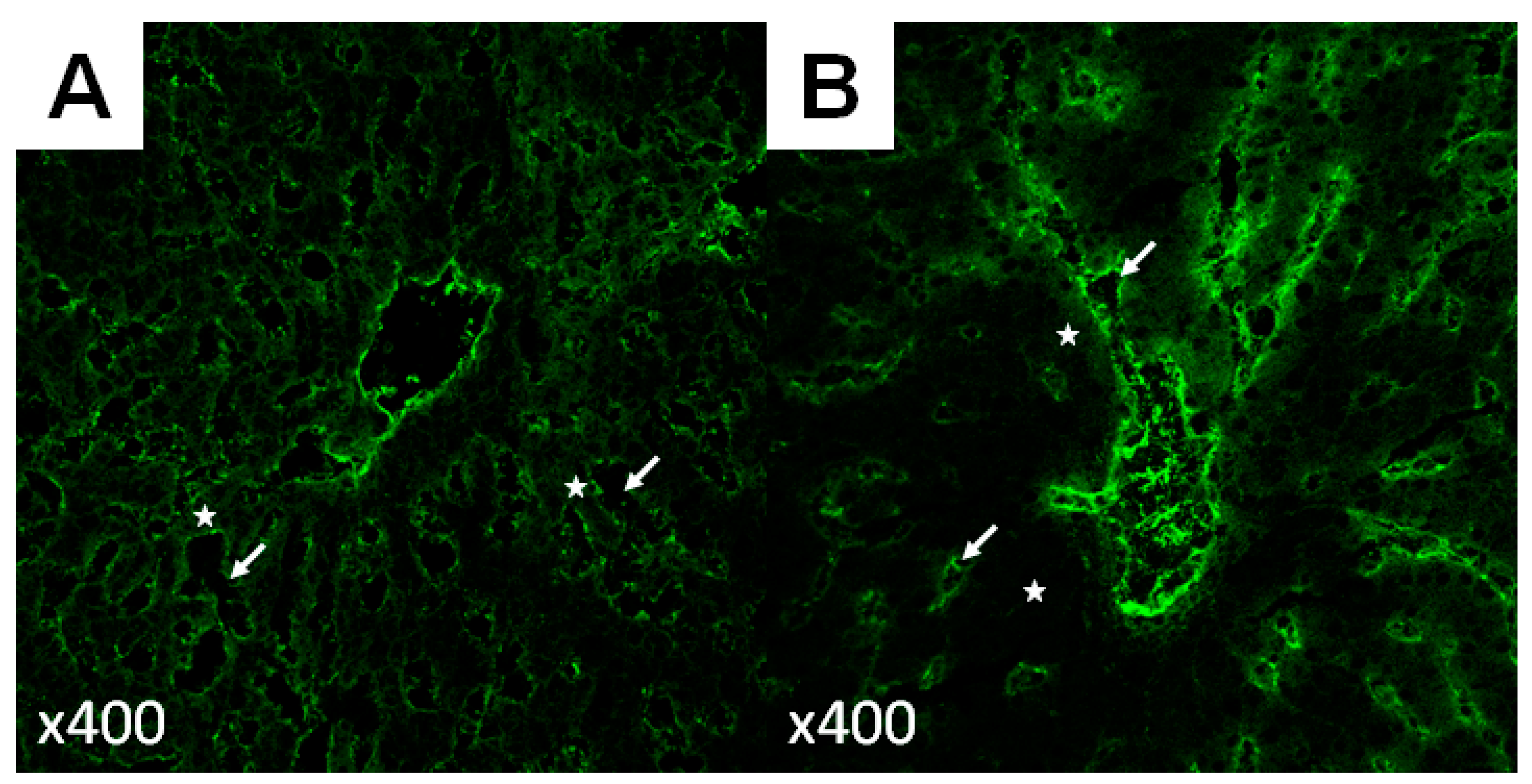
- Gruttadauria, S.; Pagano, D.; Liotta, R.; Tropea, A.; Tuzzolino, F.; Marrone, G.; Mamone, G.; Marsh, J.W.; Miraglia, R.; Luca, A.; et al. Liver Volume Restoration and Hepatic Microarchitecture in Small-for-Size Syndrome. Ann. Transplant. 2015, 20, 381–389. [Google Scholar]
- Michalopoulos, G.K.; DeFrances, M.C. Liver regeneration. Science 1997, 276, 60–66. [Google Scholar] [CrossRef]
- Ross, M.A.; Sander, C.M.; Kleeb, T.B.; Watkins, S.C.; Stolz, D.B. Spatiotemporal expression of angiogenesis growth factor receptors during the revascularization of regenerating rat liver. Hepatology 2001, 34, 1135–1148. [Google Scholar] [CrossRef]
- Belghiti, J.; Liddo, G.; Raut, V.; Zappa, M.; Dokmak, S.; Vilgrain, V.; Durand, F.; Dondero, F. “Inherent limitations” in donors: Control matched study of consequences following a right hepatectomy for living donation and benign liver lesions. Ann. Surg. 2012, 255, 528–533. [Google Scholar] [CrossRef]
- Tanaka, A.; Shinohara, H.; Hatano, E.; Sato, S.; Kanazawa, A.; Yamaoka, Y.; Torizuka, T.; Konishi, J.; Tamaki, N. Perioperative changes in hepatic function as assessed by asialoglycoprotein receptor indices by technetium 99 m galactosyl human serum albumin. Hepatogastroenterology 1999, 46, 369–375. [Google Scholar]
- Nadalin, S.; Testa, G.; Malago, M.; Beste, M.; Frilling, A.; Schroeder, T.; Jochum, C.; Gerken, G.; Broelsch, C.E. Volumetric and functional recovery of the liver after right hepatectomy for living donation. Liver Transpl. 2004, 10, 1024–1029. [Google Scholar] [CrossRef]
- Jochum, C.; Beste, M.; Penndorf, V.; Farahani, M.S.; Testa, G.; Nadalin, S.; Malago, M.; Broelsch, C.; Gerken, G. Quantitative liver function tests in donors and recipients of living donor liver transplantation. Liver Transpl. 2006, 12, 544–549. [Google Scholar] [CrossRef]
- De Graaf, W.; Bennink, R.J.; Heger, M.; Maas, A.; de Bruin, K.; van Gulik, T.M. Quantitative assessment of hepatic function during liver regeneration in a standardized rat model. J. Nucl. Med. 2011, 52, 294–302. [Google Scholar]
- Ninomiya, M.; Shirabe, K.; Terashi, T.; Ijichi, H.; Yonemura, Y.; Harada, N.; Soejima, Y.; Taketomi, A.; Shimada, M.; Maehara, Y. Deceleration of regenerative response improves the outcome of rat with massive hepatectomy. Am. J. Transplant. 2010, 10, 1580–1587. [Google Scholar] [CrossRef]
- Marbach, P.; Bauer, W.; Bodmer, D.; Briner, U.; Bruns, C.; Kay, A.; Lancranjan, I.; Pless, J.; Raulf, F.; Robinson, R.; et al. Discovery and development of somatostatin agonists. Pharm. Biotechnol. 1998, 11, 183–209. [Google Scholar]
- Hagemeister, A.L.; Sheridan, M.A. Somatostatin inhibits hepatic growth hormone receptor and insulin-like growth factor I mRNA expression by activating the ERK and PI3K signaling pathways. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 295, R490–R497. [Google Scholar] [CrossRef] [Green Version]
- Patel, Y.C. Somatostatin and its receptor family. Front. Neuroendocrinol. 1999, 20, 157–198. [Google Scholar] [CrossRef]
- Curtis, S.B.; Chen, J.C.; Winkelaar, G.; Turnbull, R.G.; Hewitt, J.; Buchan, A.M.; Hsiang, Y.N. Effect of endothelial and adventitial injury on somatostatin receptor expression. Surgery 2000, 127, 577–583. [Google Scholar] [CrossRef]
- Reynaert, H.; Vaeyens, F.; Qin, H.; Hellemans, K.; Chatterjee, N.; Winand, D.; Quartier, E.; Schuit, F.; Urbain, D.; Kumar, U.; et al. Somatostatin suppresses endothelin-1-induced rat hepatic stellate cell contraction via somatostatin receptor subtype 1. Gastroenterology 2001, 121, 915–930. [Google Scholar] [CrossRef]
- Nelson, L.E.; Sheridan, M.A. Insulin and growth hormone stimulate somatostatin receptor (SSTR) expression by inducing transcription of SSTR mRNAs and by upregulating cell surface SSTRs. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 291, R163–R169. [Google Scholar] [CrossRef] [Green Version]
- Strader, C.D.; Fong, T.M.; Graziano, M.P.; Tota, M.R. The family of G-protein-coupled receptors. FASEB J. 1995, 9, 745–754. [Google Scholar] [CrossRef]
- Oberg, K. Future aspects of somatostatin-receptor-mediated therapy. Neuroendocrinology 2004, 80 (Suppl. 1), 57–61. [Google Scholar] [CrossRef]
- Rai, U.; Thrimawithana, T.R.; Valery, C.; Young, S.A. Therapeutic uses of somatostatin and its analogues: Current view and potential applications. Pharmacol. Ther. 2015, 152, 98–110. [Google Scholar] [CrossRef]
- Bosch, J.; Kravetz, D.; Rodes, J. Effects of somatostatin on hepatic and systemic hemodynamics in patients with cirrhosis of the liver: Comparison with vasopressin. Gastroenterology 1981, 80, 518–525. [Google Scholar] [CrossRef]
- Cirera, I.; Feu, F.; Luca, A.; Garcia-Pagan, J.C.; Fernandez, M.; Escorsell, A.; Bosch, J.; Rodés, J. Effects of bolus injections and continuous infusions of somatostatin and placebo in patients with cirrhosis: A double-blind hemodynamic investigation. Hepatology 1995, 22, 106–111. [Google Scholar]
- Chatila, R.; Ferayorni, L.; Gupta, T.; Groszmann, R.J. Local arterial vasoconstriction induced by octreotide in patients with cirrhosis. Hepatology 2000, 31, 572–576. [Google Scholar] [CrossRef]
- Wiest, R.; Tsai, M.H.; Groszmann, R.J. Octreotide potentiates PKC-dependent vasoconstrictors in portal-hypertensive and control rats. Gastroenterology 2001, 120, 975–983. [Google Scholar] [CrossRef]
- Kravetz, D.; Bosch, J.; Arderiu, M.T.; Pizcueta, M.P.; Casamitjana, R.; Rivera, F.; Rodés, J. Effects of somatostatin on splanchnic hemodynamics and plasma glucagon in portal hypertensive rats. Am. J. Physiol. 1988, 254 Pt 1, G322–G328. [Google Scholar] [CrossRef]
- Pizcueta, M.P.; Garcia-Pagan, J.C.; Fernandez, M.; Casamitjana, R.; Bosch, J.; Rodes, J. Glucagon hinders the effects of somatostatin on portal hypertension. A study in rats with partial portal vein ligation. Gastroenterology 1991, 101, 1710–1715. [Google Scholar] [CrossRef]
- Sieber, C.C.; Mosca, P.G.; Groszmann, R.J. Effect of somatostatin on mesenteric vascular resistance in normal and portal hypertensive rats. Am. J. Physiol. 1992, 262 Pt 1, G274–G277. [Google Scholar] [CrossRef]
- Stoyanova, I.I.; Gulubova, M.V. Immunocytochemical study on the liver innervation in patients with cirrhosis. Acta Histochem. 2000, 102, 391–402. [Google Scholar] [CrossRef]
- Reynaert, H.; Rombouts, K.; Vandermonde, A.; Urbain, D.; Kumar, U.; Bioulac-Sage, P.; Pinzani, M.; Rosenbaum, J.; Geerts, A. Expression of somatostatin receptors in normal and cirrhotic human liver and in hepatocellular carcinoma. Gut 2004, 53, 1180–1189. [Google Scholar] [CrossRef] [Green Version]
- Reynaert, H.; Rombouts, K.; Jia, Y.; Urbain, D.; Chatterjee, N.; Uyama, N.; Geerts, A. Somatostatin at nanomolar concentration reduces collagen I and III synthesis by, but not proliferation of activated rat hepatic stellate cells. Br. J. Pharmacol. 2005, 146, 77–88. [Google Scholar] [CrossRef] [Green Version]
- Vanheule, E.; Geerts, A.M.; Reynaert, H.; Van Vlierberghe, H.; Geerts, A.; De Vos, M.; Colle, I. Influence of somatostatin and octreotide on liver microcirculation in an experimental mouse model of cirrhosis studied by intravital fluorescence microscopy. Liver Int. 2008, 28, 107–116. [Google Scholar] [CrossRef]
- Raper, S.E.; Kothary, P.C.; Kokudo, N. Somatostatin-14 blocks the hepatotrophic effects of insulin in the rat. J. Surg. Res. 1991, 50, 386–390. [Google Scholar] [CrossRef] [Green Version]
- Kokudo, N.; Kothary, P.C.; Eckhauser, F.E.; Raper, S.E. Inhibitory effects of somatostatin on rat hepatocyte proliferation are mediated by cyclic AMP. J. Surg. Res. 1991, 51, 113–118. [Google Scholar] [CrossRef] [Green Version]
- Kokudo, N.; Kothary, P.C.; Eckhauser, F.E.; Nakamura, T.; Raper, S.E. Inhibition of DNA synthesis by somatostatin in rat hepatocytes stimulated by hepatocyte growth factor or epidermal growth factor. Am. J. Surg. 1992, 163, 169–173. [Google Scholar] [CrossRef] [Green Version]
- Karalis, K.; Mastorakos, G.; Chrousos, G.P.; Tolis, G. Somatostatin analogues suppress the inflammatory reaction in vivo. J. Clin. Investig. 1994, 93, 2000–2006. [Google Scholar] [CrossRef]
- Bruno, J.F.; Xu, Y.; Song, J.; Berelowitz, M. Tissue distribution of somatostatin receptor subtype messenger ribonucleic acid in the rat. Endocrinology 1993, 133, 2561–2567. [Google Scholar] [CrossRef]
- Lang, A.; Sakhnini, E.; Fidder, H.H.; Maor, Y.; Bar-Meir, S.; Chowers, Y. Somatostatin inhibits pro-inflammatory cytokine secretion from rat hepatic stellate cells. Liver Int. 2005, 25, 808–816. [Google Scholar] [CrossRef]
- Kothary, P.C.; Kokudo, N.; Eckhauser, F.E.; DelValle, J.; Raper, S.E. Preferential suppression of insulin-stimulated proliferation of cultured hepatocytes by somatostatin: Evidence for receptor-mediated growth regulation. J. Cell Biochem. 1995, 59, 258–265. [Google Scholar] [CrossRef] [Green Version]
- Pinter, E.; Helyes, Z.; Szolcsanyi, J. Inhibitory effect of somatostatin on inflammation and nociception. Pharmacol. Ther. 2006, 112, 440–456. [Google Scholar] [CrossRef]
- Lamberts, S.W.; van der Lely, A.J.; de Herder, W.W.; Hofland, L.J. Octreotide. N. Engl. J. Med. 1996, 334, 246–254. [Google Scholar] [CrossRef] [Green Version]
- De Franchis, R. Somatostatin, somatostatin analogues and other vasoactive drugs in the treatment of bleeding oesophageal varices. Dig. Liver Dis. 2004, 36 (Suppl. 1), S93–S100. [Google Scholar]
- Chen, Z.B.; Hedner, T.; Hedner, J. Local application of somatostatin in the rat ventrolateral brain medulla induces apnea. J. Appl. Physiol. 1990, 69, 2233–2238. [Google Scholar] [CrossRef]
- Halma, C.; Jansen, J.B.; Janssens, A.R.; Griffioen, G.; Lamers, C.B. Life-threatening water intoxication during somatostatin therapy. Ann. Int. Med. 1987, 107, 518–520. [Google Scholar] [CrossRef]
- Davies, N.; Yates, J.; Kynaston, H.; Taylor, B.A.; Jenkins, S.A. Effects of octreotide on liver regeneration and tumour growth in the regenerating liver. J. Gastroenterol. Hepatol. 1997, 12, 47–53. [Google Scholar] [CrossRef]
- Papalampros, E.; Felekouras, E.S.; Filis, K.; Angelopoulou, R.; Kourelis, T.; Tsamandas, A.C.; Bastounis, E. Liver pathology and cell proliferation after octreotide administration following partial hepatectomy in rats: An experimental study. Dig. Dis. Sci. 2002, 47, 1953–1958. [Google Scholar] [CrossRef]
- Davies, N.; Kynaston, H.; Yates, J.; Nott, D.M.; Nash, J.; Taylor, B.A.; Jenkins, S.A. Octreotide inhibits the growth and development of three types of experimental liver metastases. Br. J. Surg. 1995, 82, 840–843. [Google Scholar] [CrossRef]
- Pyronnet, S.; Bousquet, C.; Najib, S.; Azar, R.; Laklai, H.; Susini, C. Antitumor effects of somatostatin. Mol. Cell. Endocrinol. 2008, 286, 230–237. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.S.; Liang, Q.M.; Li, H.S.; Yang, J.; Wang, S.; Long, J.W. Octreotide inhibits growth of colonic cancer SW480 cells by modulating the Wnt/P-catenin pathway. Pharmazie 2009, 64, 126–131. [Google Scholar]
- Wang, S.; Bao, Z.; Liang, Q.M.; Long, J.W.; Xiao, Z.S.; Jiang, Z.J.; Liu, B.; Yang, J.; Long, Z.X. Octreotide stimulates somatostatin receptor-induced apoptosis of SW480 colon cancer cells by activation of glycogen synthase kinase-3beta, A Wnt/beta-catenin pathway modulator. Hepatogastroenterology 2013, 60, 1639–1646. [Google Scholar]
- Modarai, S.R.; Opdenaker, L.M.; Viswanathan, V.; Fields, J.Z.; Boman, B.M. Somatostatin signaling via SSTR1 contributes to the quiescence of colon cancer stem cells. BMC Cancer 2016, 16, 941. [Google Scholar] [CrossRef]
- Di Bella, G.; Colori, B.; Scanferlato, R. The over-expression of GH/GHR in tumour tissues with respect to healthy ones confirms its oncogenic role and the consequent oncosuppressor role of its physiological inhibitor, somatostatin: A review of the literature. Neuro Endocrinol. Lett. 2018, 39, 179–188. [Google Scholar]
- Huang, C.Z.; Huang, A.M.; Liu, J.F.; Wang, B.; Lin, K.C.; Ye, Y.B. Somatostatin Octapeptide Inhibits Cell Invasion and Metastasis in Hepatocellular Carcinoma Through PEBP1. Cell. Physiol. Biochem. 2018, 47, 2340–2349. [Google Scholar] [CrossRef]
- Du, Z.; Zhou, Y.; Lu, X.; Li, L.; Lu, C.; Li, L.; Li, B.; Bu, H.; Yang, J.; Shi, Y. Octreotide prevents liver failure through upregulating 5′-methylthioadenosine in extended hepatectomized rats. Liver Int. 2016, 36, 212–222. [Google Scholar] [CrossRef]
- Brunmark, A.; Cadenas, E. Redox and addition chemistry of quinoid compounds and its biological implications. Free Radic. Biol. Med. 1989, 7, 435–477. [Google Scholar] [CrossRef]
- Corrales, F.; Gimenez, A.; Alvarez, L.; Caballeria, J.; Pajares, M.A.; Andreu, H.; Parés, A.; Mató, J.M.; Rodés, J. S-adenosylmethionine treatment prevents carbon tetrachloride-induced S-adenosylmethionine synthetase inactivation and attenuates liver injury. Hepatology 1992, 16, 1022–1027. [Google Scholar] [CrossRef]
- Frau, M.; Feo, F.; Pascale, R.M. Pleiotropic effects of methionine adenosyltransferases deregulation as determinants of liver cancer progression and prognosis. J. Hepatol. 2013, 59, 830–841. [Google Scholar] [CrossRef] [Green Version]
- Mohkam, K.; Darnis, B.; Schmitt, Z.; Duperret, S.; Ducerf, C.; Mabrut, J.Y. Successful modulation of portal inflow by somatostatin in a porcine model of small-for-size syndrome. Am. J. Surg. 2016, 212, 321–326. [Google Scholar] [CrossRef]
- Rhaiem, R.; Piardi, T.; Chetboun, M.; Pessaux, P.; Lestra, T.; Memeo, R.; Kianmanesh, R.; Sommacale, D. Portal Inflow Modulation by Somatostatin After Major Liver Resection: A Pilot Study. Ann. Surg. 2018, 267, e101–e103. [Google Scholar] [CrossRef]
- Kiuchi, T.; Onishi, Y.; Nakamura, T. Small-for-size graft: Not defined solely by being small for size. Liver Transpl. 2010, 16, 815–817. [Google Scholar] [CrossRef]
- Fondevila, C.; Busuttil, R.W.; Kupiec-Weglinski, J.W. Hepatic ischemia/reperfusion injury—A fresh look. Exp. Mol. Pathol. 2003, 74, 86–93. [Google Scholar] [CrossRef]
- Xu, X.; Man, K.; Zheng, S.S.; Liang, T.B.; Lee, T.K.; Ng, K.T.; Fan, S.T.; Lo, C.M. Attenuation of acute phase shear stress by somatostatin improves small-for-size liver graft survival. Liver Transpl. 2006, 12, 621–627. [Google Scholar] [CrossRef]
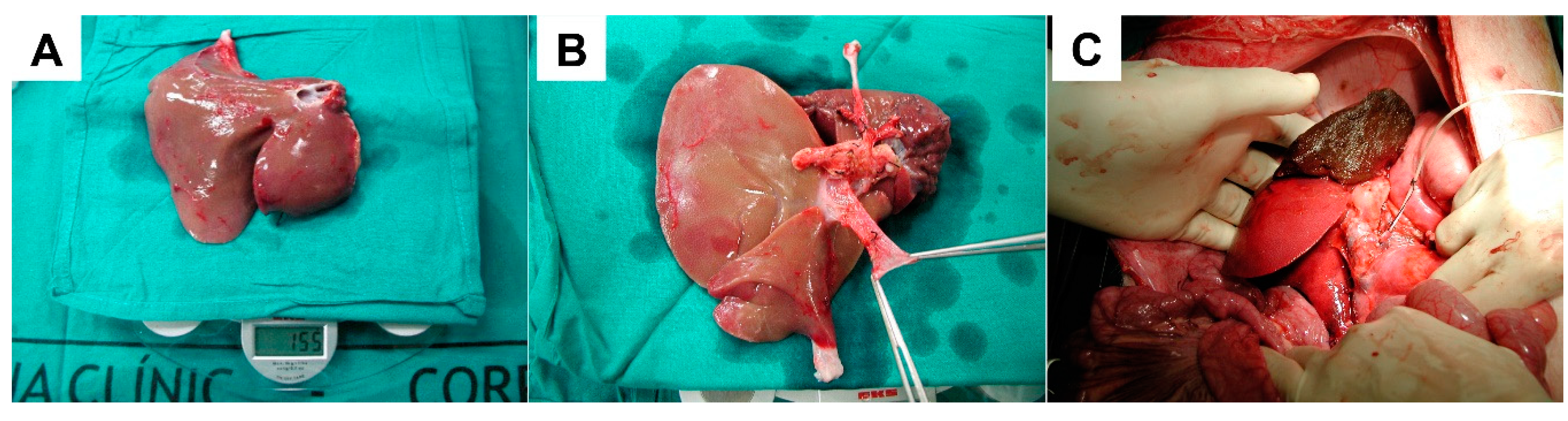
- Fondevila, C.; Hessheimer, A.J.; Flores, E.; Vendrell, M.; Munoz, J.; Escobar, B.; Calatayud, D.; Taurá, P.; Fuster, J.; García-Valdecasas, J.C. Step-by-step guide for a simplified model of porcine orthotopic liver transplant. J. Surg. Res. 2011, 167, e39–e45. [Google Scholar] [CrossRef]
- Ozden, I.; Kara, M.; Pinarbasi, B.; Salmaslioglu, A.; Yavru, A.; Kaymakoglu, S.; Emre, A.; Bilge, O.; Alper, A. Somatostatin and propranolol to treat small-for-size syndrome that occurred despite splenic artery ligation. Exp. Clin. Transplant. 2007, 5, 686–689. [Google Scholar]
- Feng, Z.Y.; Xu, X.; Wu, L.J.; Wu, J.; Zhu, S.M.; Zheng, S.S. Downregulation of endothelin-1 by somatostatin improves liver function of recipients undergoing adult-to-adult living donor liver transplantation. Chin. Med. J. (Engl.) 2010, 123, 1961–1966. [Google Scholar]
- Lee, T.Y.; Fan, H.L.; Wang, C.W.; Hsieh, C.B.; Chen, T.W. Somatostatin Therapy in Patients with Massive Ascites After Liver Transplantation. Ann. Transplant. 2019, 24, 1–8. [Google Scholar] [CrossRef]
- Troisi, R.I.; Vanlander, A.; Giglio, M.C.; Van, L.J.; Scudeller, L.; Heyse, B.; de Baerdemaeker, L.; Croo, A.; Voet, D.; Praet, M.; et al. Somatostatin as Inflow Modulator in Liver-transplant Recipients with Severe Portal Hypertension: A Randomized Trial. Ann. Surg. 2018. [Google Scholar] [CrossRef]
- Mohkam, K.; Rayar, M.; Adam, J.P.; Muscari, F.; Rode, A.; Merle, P.; Pradat, P.; Bauler, S.; Delfour, I.; Chiche, L.; et al. Evaluation of postoperative ascites after somatostatin infusion following hepatectomy for hepatocellular carcinoma by laparotomy: A multicenter randomized double-blind controlled trial (SOMAPROTECT). BMC Cancer 2018, 18, 844. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Reduces | Improves |
|---|---|
|
|
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hessheimer, A.J.; Martínez de la Maza, L.; Adel Al Shwely, F.; Espinoza, A.S.; Ausania, F.; Fondevila, C. Somatostatin and the “Small-For-Size” Liver. Int. J. Mol. Sci. 2019, 20, 2512. https://doi.org/10.3390/ijms20102512
Hessheimer AJ, Martínez de la Maza L, Adel Al Shwely F, Espinoza AS, Ausania F, Fondevila C. Somatostatin and the “Small-For-Size” Liver. International Journal of Molecular Sciences. 2019; 20(10):2512. https://doi.org/10.3390/ijms20102512
Chicago/Turabian StyleHessheimer, Amelia J., Lilia Martínez de la Maza, Farah Adel Al Shwely, Arlena Sofía Espinoza, Fabio Ausania, and Constantino Fondevila. 2019. "Somatostatin and the “Small-For-Size” Liver" International Journal of Molecular Sciences 20, no. 10: 2512. https://doi.org/10.3390/ijms20102512