High Expression of IL-1RI and EP2 Receptors in the IL-1β/COX-2 Pathway, and a New Alternative to Non-Steroidal Drugs—Osthole in Inhibition COX-2

 , ,
, ,
Abstract
:
1. Introduction
2. Results
2.1. Basal Expression of HRH-1, IL-1RI, COX-2, and EP2 Receptors
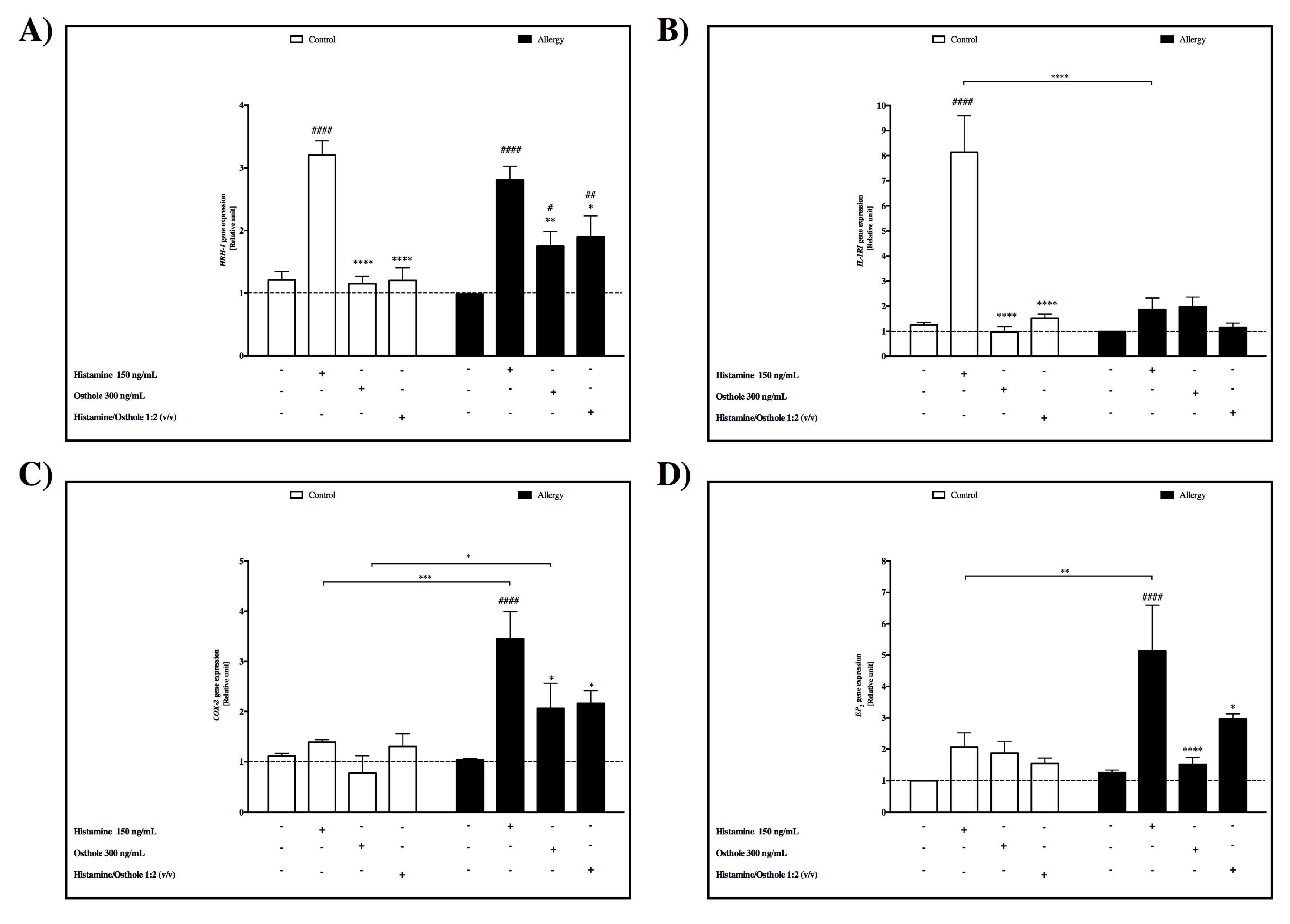
2.2. HRH-1 Gene Expression Induced by Histamine
- Osthole effect
2.3. IL-1RI Gene Expression
- Induced by histamine
- Osthole effect
2.4. COX-2 Gene Expression
- Induced by histamine
- Osthole effect
2.5. EP2 Gene Expression
- Induced by histamine
- Osthole effect
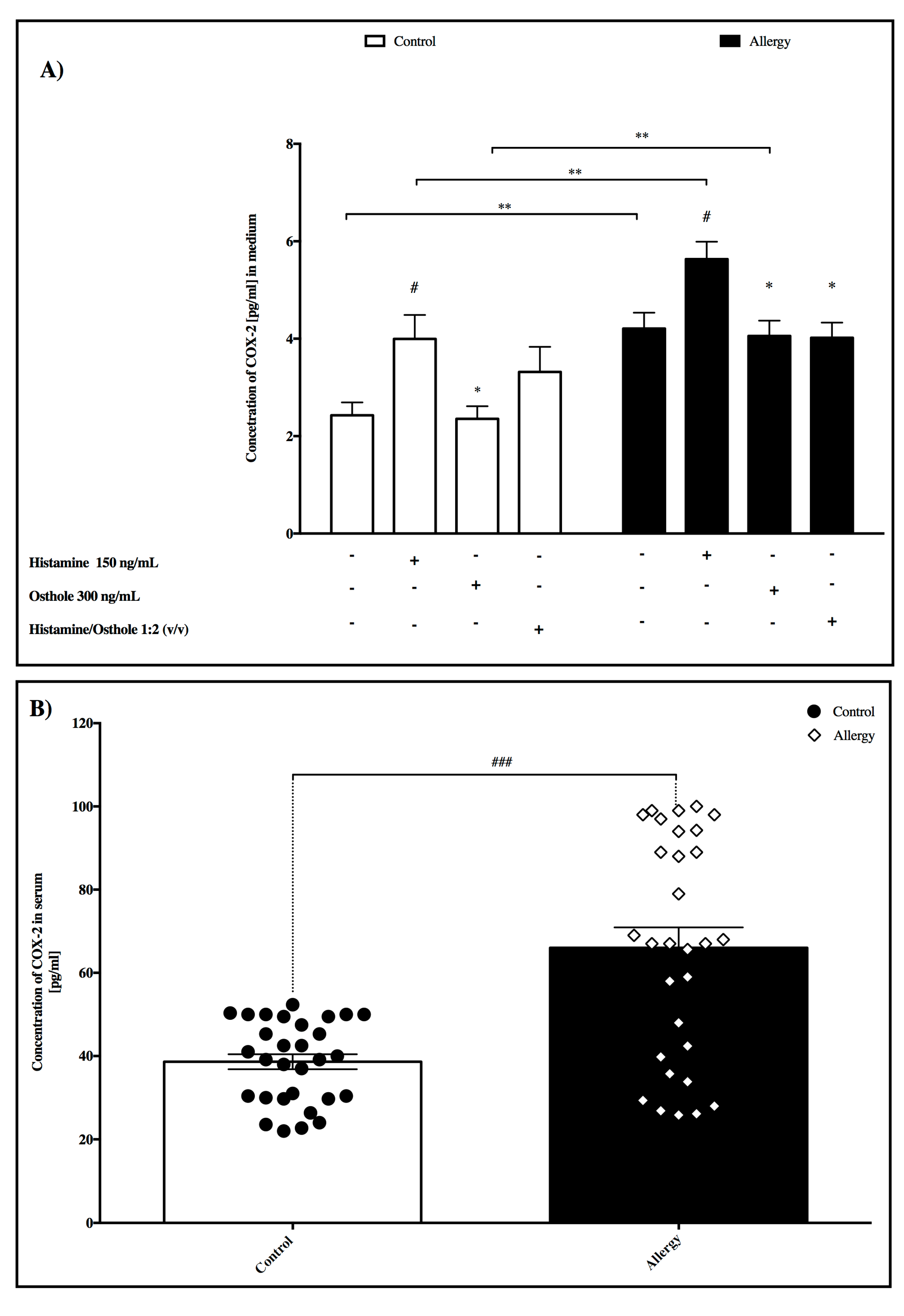
2.6. IL-1β Concentration
- In medium
- In serum
2.7. COX-2 Concentration
- In medium
- In serum
2.8. The Inhibitory Effects of COX-2—Activity of COX-2
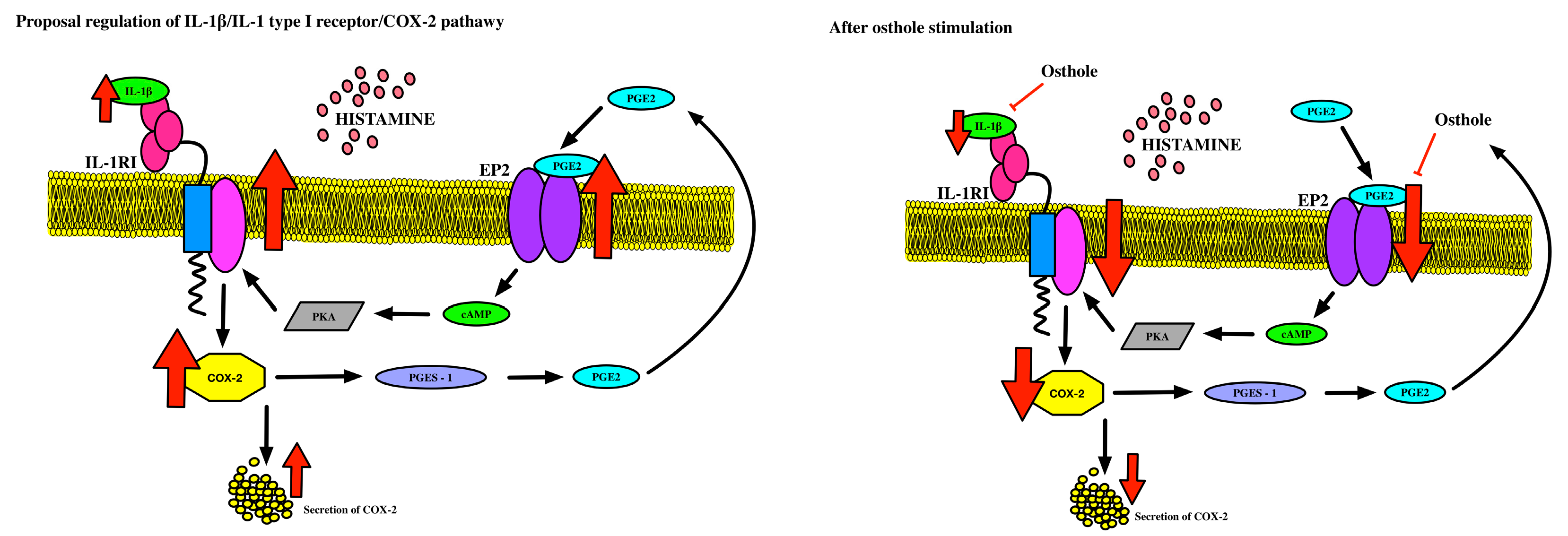
3. Discussion
4. Material and Methods
4.1. Participants
4.2. Biological Material
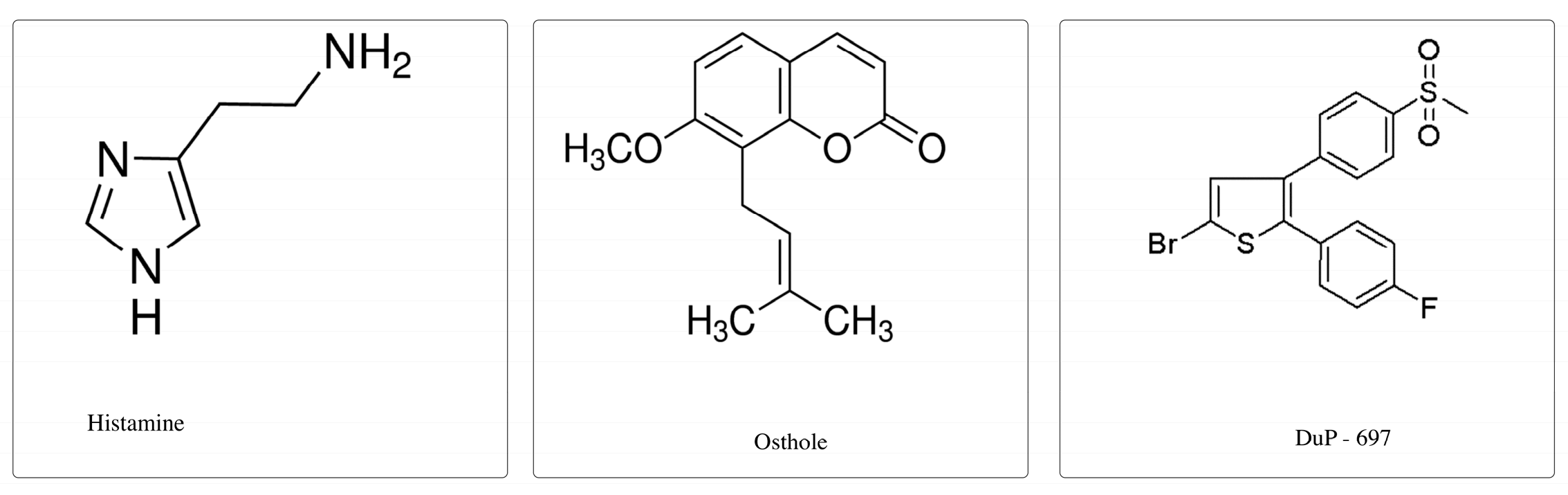
4.3. Examined Substances
4.4. PBMC Isolation
4.5. Analysis of IL-1RI, EP2, and COX-2 Gene Expression
4.6. Measurement of COX-2 Concentration
4.7. Assay of COX-2 Enzymatic Activity
4.8. Measurement of IL-1β
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kim, K.H.; Jahan, S.A.; Kabir, E. A review on human health perspective of air pollution with respect to allergies and asthma. Environ. Int. 2013, 59, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.A.; Burks, A.W. Food allergies: prevalence, molecular characterization, and treatment/prevention strategies. Ann. Rev. Nutr. 2006, 26, 539–565. [Google Scholar] [CrossRef] [PubMed]
- Thompson-Souza, G.A.; Gropillo, I.; Neves, J.S. Cysteinyl Leukotrienes in Eosinophil Biology: Functional Roles and Therapeutic Perspectives in Eosinophilic Disorders. Front. Med. 2017, 4, 106. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, Y.; Narumiya, S. Prostaglandin E receptors. J. Boil. Chem. 2007, 282, 11613–11617. [Google Scholar] [CrossRef] [PubMed]
- Meijer, H.; Reinecke, J.; Becker, C.; Tholen, G.; Wehling, P. The production of anti-inflammatory cytokines in whole blood by physic-chemical induction. Inflamm. Res. 2003, 52, 404–407. [Google Scholar] [CrossRef] [PubMed]
- Ren, K.; Torres, R. Role of interleukin-1β during pain and inflammation. Brain Res. Rev. 2009, 60, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Dinarello, C.A. The IL-1 family and inflammatory diseases. Clin. Exp. Rheumatol. 2002, 20, S1–S13. [Google Scholar]
- Weber, A.; Wasiliew, P.; Kracht, M. Interleukin-1 (IL-1) pathway. Sci. Signal. 2010, 3, cm1. [Google Scholar] [CrossRef]
- Roca-Ferrer, J.; Pérez-Gonzalez, M.; Garcia-Garcia, F.J.; Pereda, J.; Pujols, L.; Alobid, I.; Mullol, J.; Picado, C. Low prostaglandin E2 and cyclooxygenase expression in nasal mucosa fibroblasts of aspirin-intolerant asthmatics. Respirology 2013, 18, 711–717. [Google Scholar] [CrossRef]
- Mullol, J.; Fernàndez-Morata, J.C.; Roca-Ferrer, J.; Pujols, L.; Xaubet, A.; Benitez, P.; Picado, C. Cyclooxygenase 1 and cyclooxygenase 2 expression is abnormally regulated in human nasal polyps. J. Allergy Clin. Immunol. 2002, 109, 824–830. [Google Scholar] [CrossRef]
- Samad, T.A.; Moore, K.A.; Sapirstein, A.; Billet, S. Interleukin-1beta-mediated induction of Cox-2 in the CNS contributes to inflammatory pain hypersensitivity. Nature 2001, 410, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Takii, T.; Akahoshi, T.; Kato, K.; Hayashi, H.; Marunouchi, T.; Onozaki, K. Interleukin-1 up-regulates transcription of its own receptor in a human fibroblast cell line TIG-1: Role of endogenous PGE2 and cAMP. Eur. J. Immunol. 1992, 22, 1221–1227. [Google Scholar] [CrossRef]
- Klein, T.; Shephard, P.; Kleinert, H.; Kömhoff, M. Regulation of cyclooxygenase-2 expression by cyclic AMP. Biochim. Biophys. Acta Mol. Cell Res. 2007, 1773, 1605–1618. [Google Scholar] [CrossRef] [Green Version]
- Kordulewska, N.K.; Kostyra, E.; Cieślińska, A.; Fiedorowicz, E.; Jarmołowska, B. Cytokine production by PBMC and serum from allergic and non-allergic subjects following in vitro histamine stimulation to test fexofenadine and osthole anti-allergic properties. Eur. J. Pharmacol. 2016, 791, 763–772. [Google Scholar] [CrossRef] [PubMed]
- Kordulewska, N.K.; Kostyra, E.; Cieślińska, A.; Matysiewicz, M.; Fiedorowicz, E.; Sienkiewicz-Szłapka, E. Changes in gene expression induced by histamine, fexofenadine and osthole: Expression of histamine H1 receptor, COX-2, NF-κB, CCR1, chemokine CCL5/RANTES and interleukin-1β in PBMC allergic and non-allergic patients. Immunobiology 2017, 222, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Kordulewska, N.K.; Kostyra, E.; Matysiewicz, M.; Cieślińska, A.; Jarmołowska, B. Impact of fexofenadine, osthole and histamine on peripheral blood mononuclear cell proliferation and cytokine secretion. Eur. J. Pharmacol. 2015, 761, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Kunikata, T.; Yamane, H.; Segi, E.; Matsuoka, T.; Sugimoto, Y.; Tanaka, S.; Tanaka, H.; Nagai, H.; Ichikawa, A.; Narumiya, S. Suppression of allergic inflammation by the prostaglandin E receptor subtype EP3. Nat. Immunol. 2005, 6, 524–531. [Google Scholar] [CrossRef]
- Kordulewska, N.K.; Kostyra, E.; Chwała, B.; Moszyńska, M.; Cieślińska, A.; Fiedorowicz, E.; Jarmołowska, B. A novel concept of immunological and allergy interactions in Autism Spectrum Disorders: Molecular, anti-inflammatory effect of osthole. Int. Immunopharmacol. In review.
- Nagai, H. Prostaglandin as a target molecule for pharmacotherapy of allergic inflammatory diseases. Allergol. Int. 2008, 57, 187–196. [Google Scholar] [CrossRef]
- Honda, T.; Tokura, Y.; Miyachi, Y.; Kabashima, K. Prostanoid receptors as possible targets for anti-allergic drugs: recent advances in prostanoids on allergy and immunology. Curr. Drug Targets 2010, 11, 1605–1613. [Google Scholar] [CrossRef]
- Singh, G.; Bhatti, R.; Mannan, R.; Singh, D.; Kesavan, A.; Singh, P. Osthole ameliorates neurogenic and inflammatory hyperalgesia by modulation of iNOS, COX-2, and inflammatory cytokines in mice. Inflammopharmacology 2018, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Jutel, M.; Watanabe, T.; Akdis, M.; Blaser, K.; Akdis, C.A. Immune regulation by histamine. Curr. Opin. Immunol. 2002, 14, 735–740. [Google Scholar] [CrossRef]
- Dinarello, C.A. Biologic basis for interleukin-1 in disease. Blood 1996, 87, 2095–2147. [Google Scholar] [PubMed]
- Martin, M.U.; Wesche, H. Summary and comparison of the signaling mechanisms of the Toll/interleukin-1 receptor family. Biochim. Biophys. Acta Mol. Cell Res. 2002, 1592, 265–280. [Google Scholar] [CrossRef] [Green Version]
- Ariasnegrete, S.; Keller, K.; Chadee, K. Proinflammatory cytokines regulate cyclooxygenase-2 mRNA expression in human macrophages. Biochem. Biophys. Res. Commun. 1995, 208, 582–589. [Google Scholar] [CrossRef]
- Payvandi, F.; Wu, L.; Haley, M.; Schafer, P.H.; Zhang, L.H.; Chen, R.S.; Muller, G.W.; Stirling, D.I. Immunomodulatory drugs inhibit expression of cyclooxygenase-2 from TNF-α, IL-1β, and LPS-stimulated human PBMC in a partially IL-10-dependent manner. Cell. Immunol. 2004, 230, 81–88. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, G.P.; Ford-Hutchinson, A.W. Expression of mRNA for cyclooxygenase-1 and cyclooxygenase-2 in human tissues. FEBS Lett. 1993, 330, 157–160. [Google Scholar] [CrossRef] [Green Version]
- Cianchi, F.; Cortesini, C.; Schiavone, N.; Perna, F.; Magnelli, L.; Fanti, E.; Bani, D.; Messerini, L.; Fabbroni, V.; Capaccioli, S.; et al. The role of cyclooxygenase-2 in mediating the effects of histamine on cell proliferation and vascular endothelial growth factor production in colorectal cancer. Clin. Cancer Res. 2005, 11, 6807–6815. [Google Scholar] [CrossRef]
- Sousa, A.; Pfister, R.; Christie, P.E.; Lane, S.J.; Nasser, S.M.; Schmitz-Schumann, M.; Lee, T.H. Enhanced expression of cyclo-oxygenase isoenzyme 2 (COX-2) in asthmatic airways and its cellular distribution in aspirin-sensitive asthma. Thorax 1997, 52, 940–945. [Google Scholar] [CrossRef] [Green Version]
- Chung, K.F.; Barnes, P.J. Cytokines in asthma. Thorax 1999, 54, 825–857. [Google Scholar] [CrossRef] [Green Version]
- Narumiya, S.; Sugimoto, Y.; Ushikubi, F. Prostanoid receptors: structures, properties, and functions. Physiol. Rev. 1999, 79, 1193–1226. [Google Scholar] [CrossRef] [PubMed]
- Machado-Carvalho, L.; Martín, M.; Torres, R.; Gabasa, M.; Alobid, I.; Mullol, J.; Pujols, L.; Roca-Ferrer, J.; Picado, C. Low E-prostanoid 2 receptor levels and deficient induction of the IL-1β/IL-1 type I receptor/COX-2 pathway: Vicious circle in patients with aspirin-exacerbated respiratory disease. J. Allergy Clin. Immunol. 2016, 137, 99–107. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Control | Allergies |
|---|---|---|
| Patient’s samples, no. | 28 | 30 |
| Age (y), mean ± SD | 7.60 ± 1.93 | 4 ± 1.75 |
| Female sex, no. (%) | 10 (35,7%) | 8 (26,6%) |
| Diagnosed Allergy, no. (%) | 0 (0%) | 30 (100%) |
| Moderate/severe asthma, no. (%) | 0 (0%) | 15 (50%) |
| Skin prick test positivity, no. (%) | 0 (0%) | 30 (100%) |
| cIgE [IU/mL] mean ± SD | 90.13 ± 43.93 **** | 307.7 ± 130.0 |
| a-sIgE [IU/mL] class | Negative predictive value, class 0 | Positive predictive value, class 4–6 |
| Blood eosinophilia (mean % of eosinophils in the blood smear) mean ± SD | 3.668 ± 1.104 **** | 23.92 ± 5.587 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kordulewska, N.K.; Cieślińska, A.; Fiedorowicz, E.; Jarmołowska, B.; Kostyra, E. High Expression of IL-1RI and EP2 Receptors in the IL-1β/COX-2 Pathway, and a New Alternative to Non-Steroidal Drugs—Osthole in Inhibition COX-2. Int. J. Mol. Sci. 2019, 20, 186. https://doi.org/10.3390/ijms20010186
Kordulewska NK, Cieślińska A, Fiedorowicz E, Jarmołowska B, Kostyra E. High Expression of IL-1RI and EP2 Receptors in the IL-1β/COX-2 Pathway, and a New Alternative to Non-Steroidal Drugs—Osthole in Inhibition COX-2. International Journal of Molecular Sciences. 2019; 20(1):186. https://doi.org/10.3390/ijms20010186
Chicago/Turabian StyleKordulewska, Natalia Karolina, Anna Cieślińska, Ewa Fiedorowicz, Beata Jarmołowska, and Elżbieta Kostyra. 2019. "High Expression of IL-1RI and EP2 Receptors in the IL-1β/COX-2 Pathway, and a New Alternative to Non-Steroidal Drugs—Osthole in Inhibition COX-2" International Journal of Molecular Sciences 20, no. 1: 186. https://doi.org/10.3390/ijms20010186