Genome-Wide Identification and Analysis of Apple NITRATE TRANSPORTER 1/PEPTIDE TRANSPORTER Family (NPF) Genes Reveals MdNPF6.5 Confers High Capacity for Nitrogen Uptake under Low-Nitrogen Conditions


Abstract
:1. Introduction
2. Results
2.1. Identification and Annotation of NPF Genes in Apple
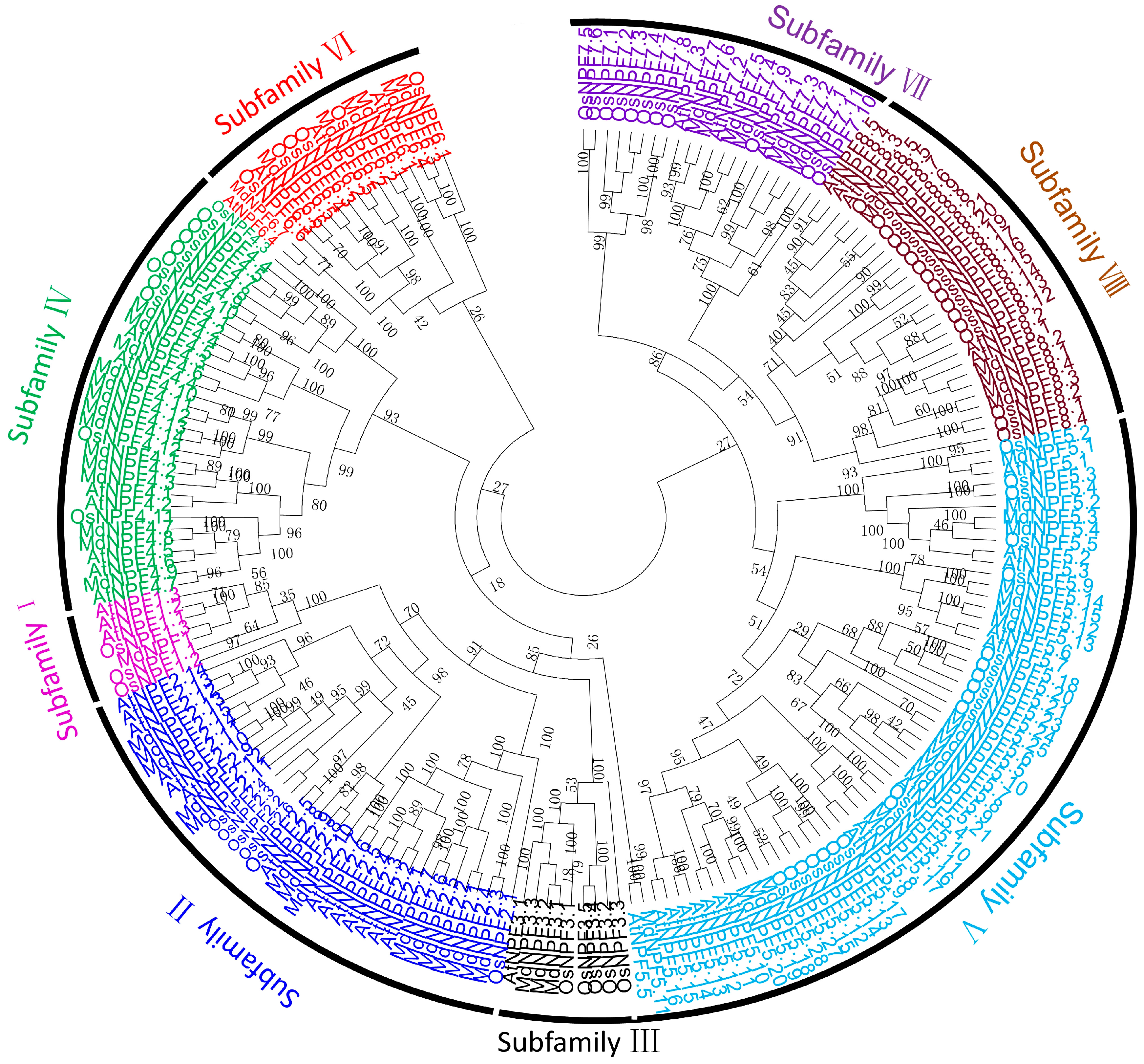
2.2. Phylogenetic Tree of NPF in Apple
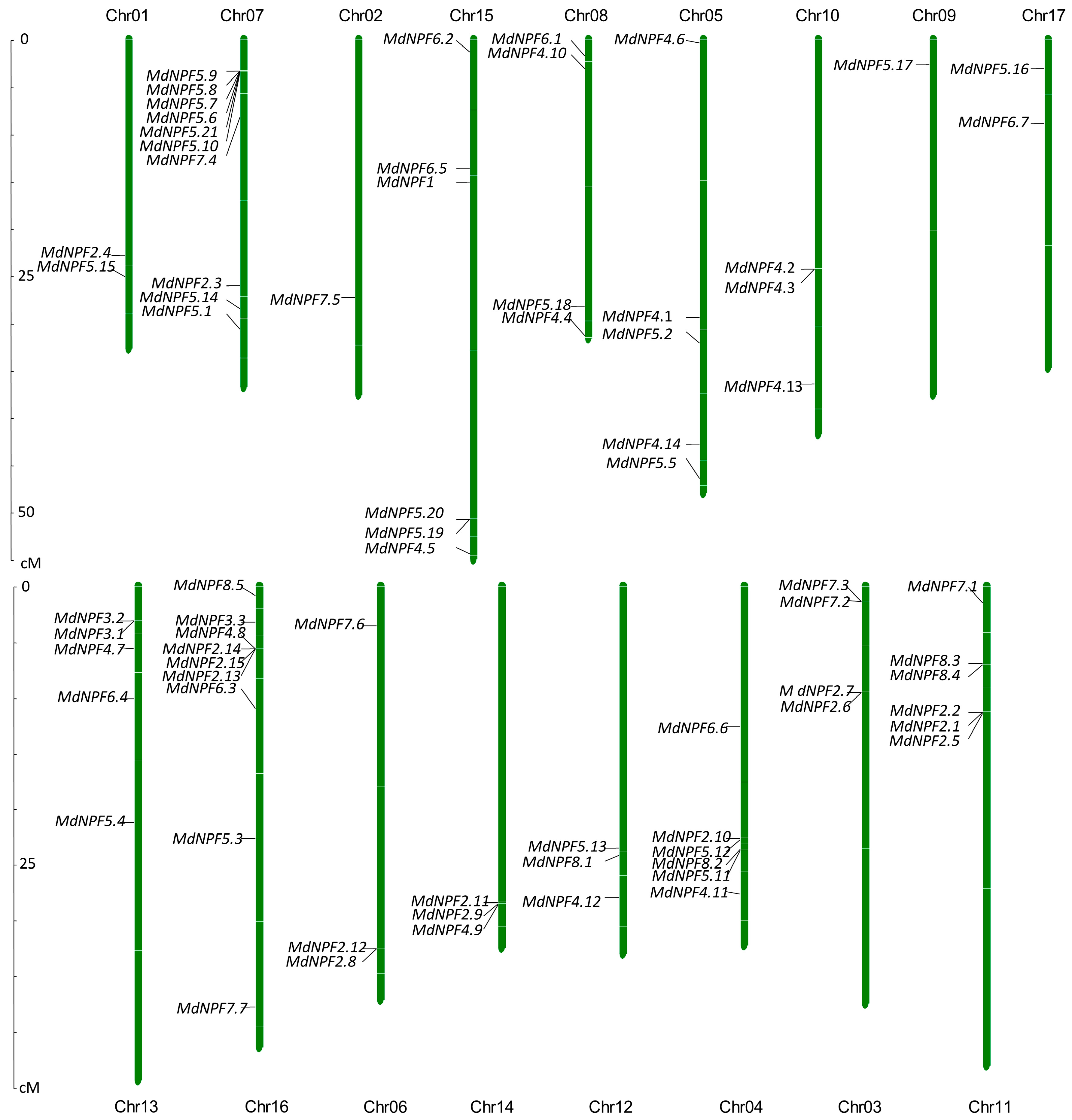
2.3. Chromosomal Localization Analysis of NPFs in Apple
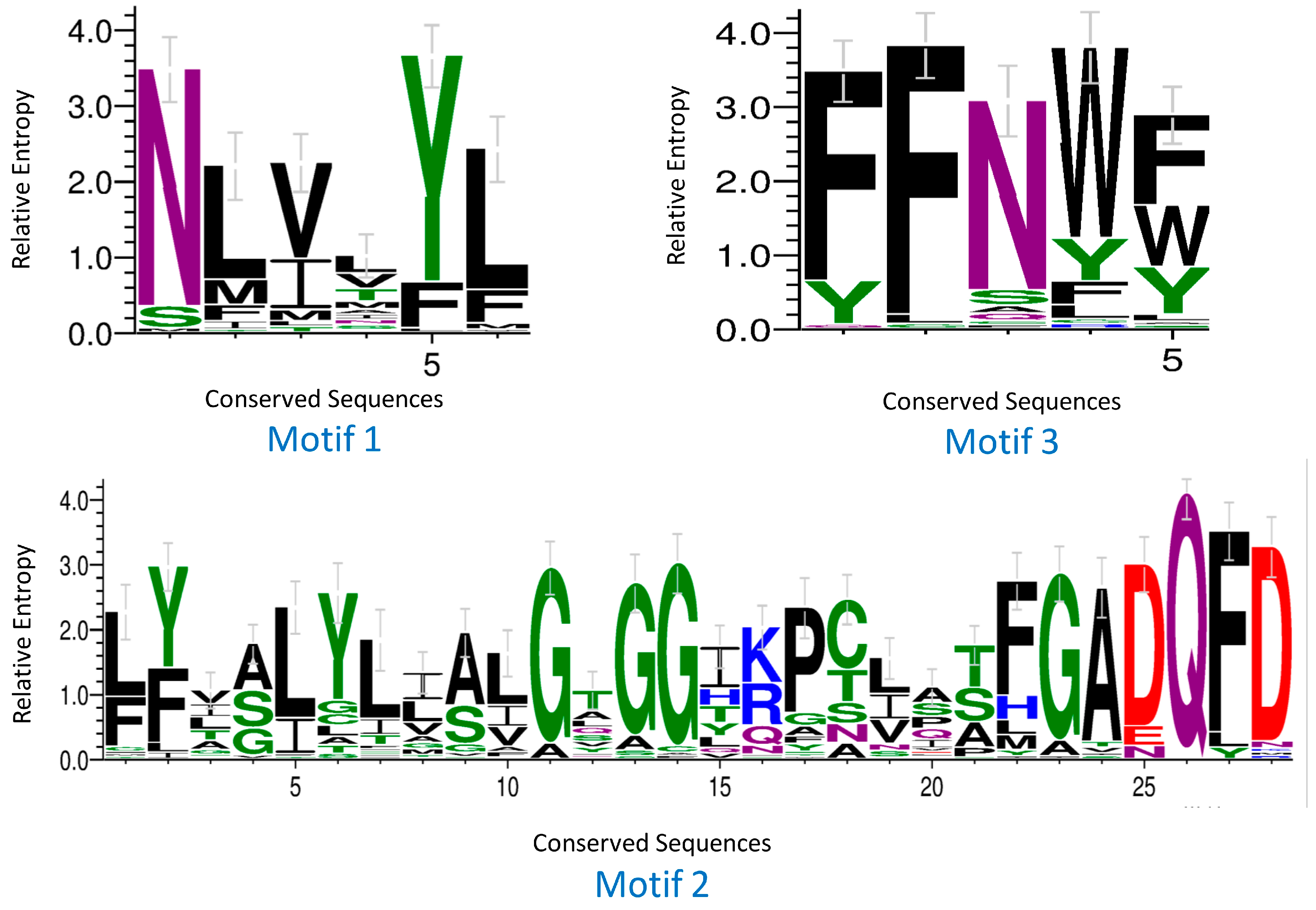
2.4. Analyses of Conserved Domains and TM Regions
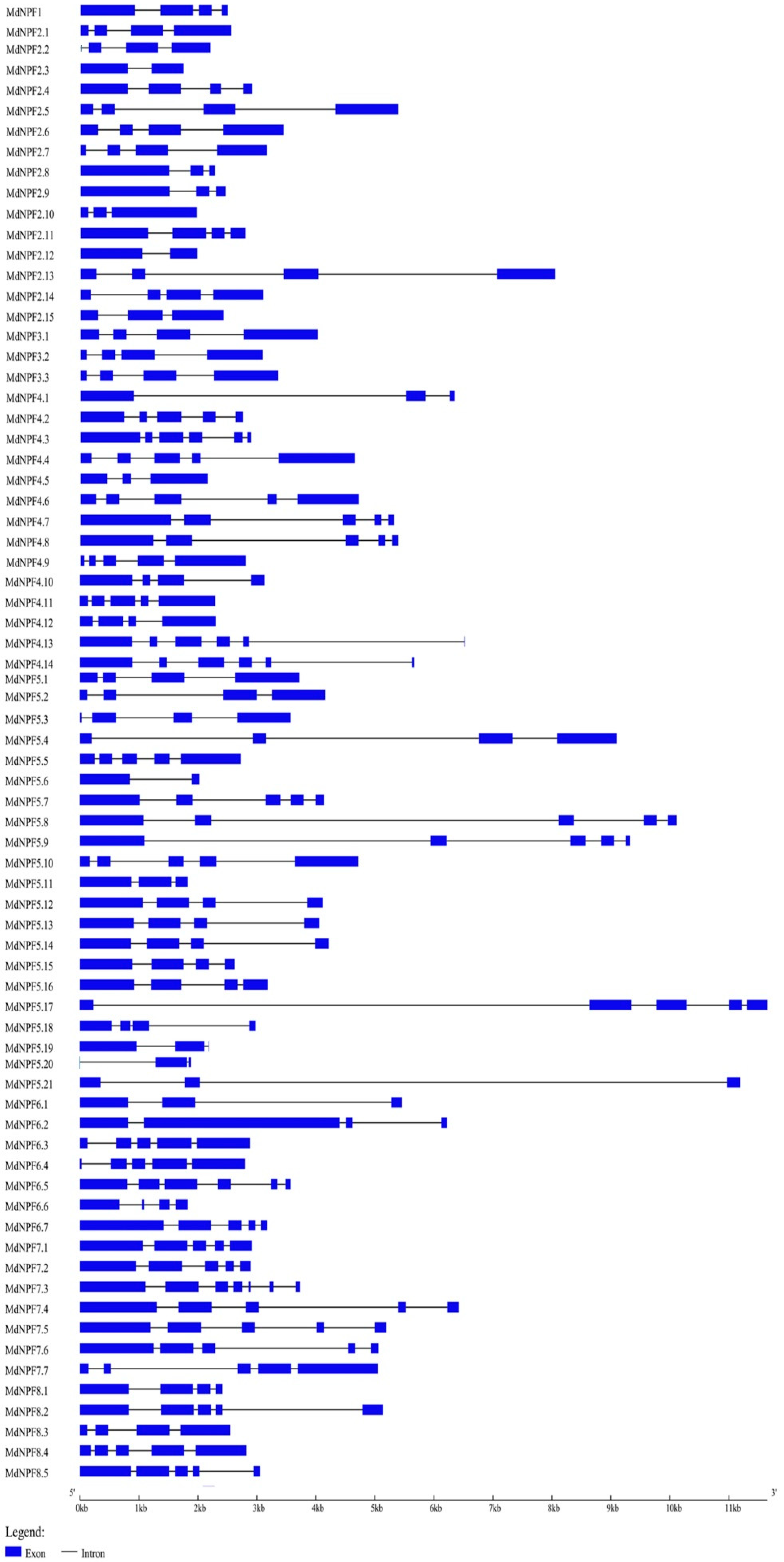
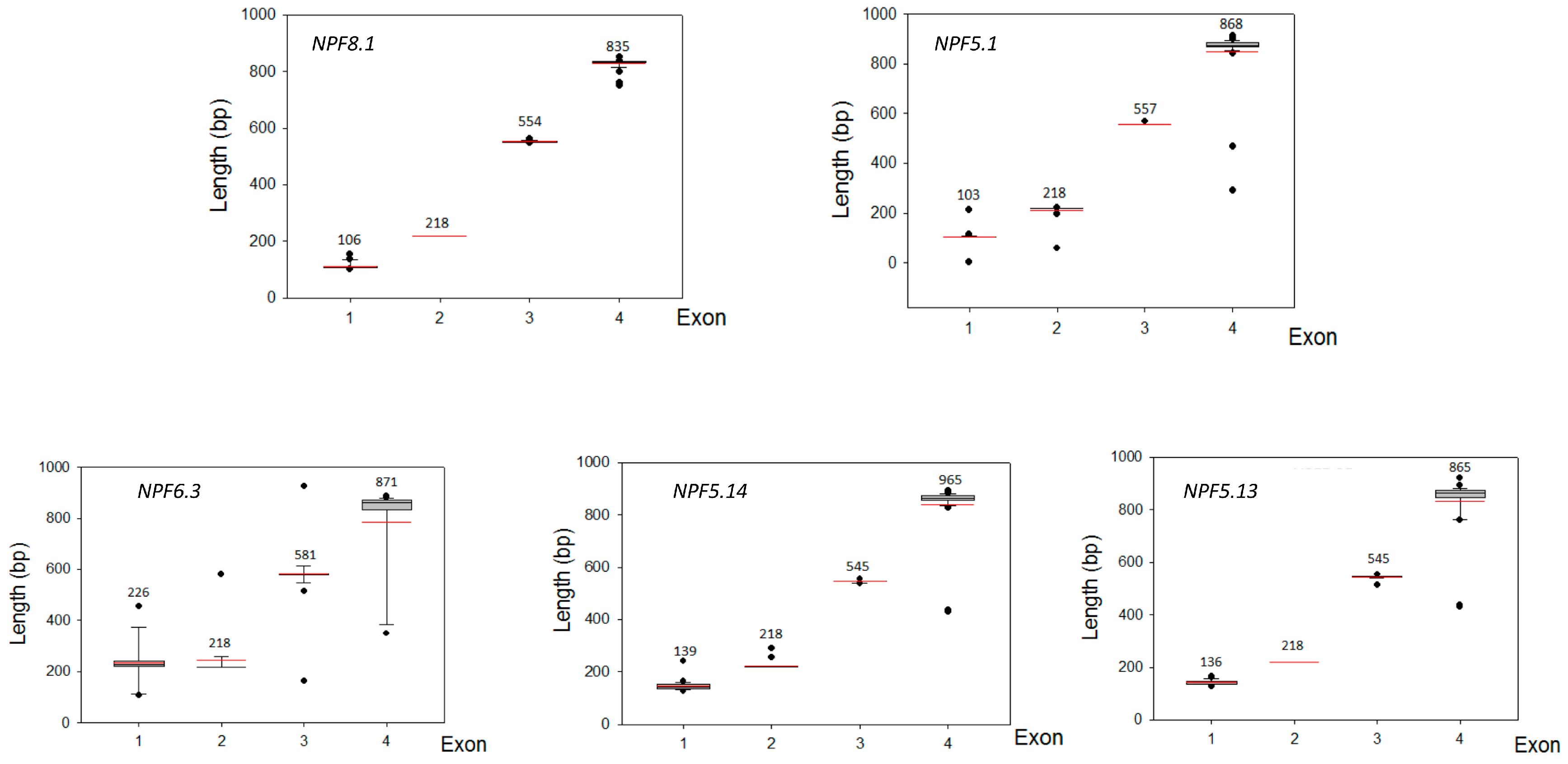
2.5. Comparison of Exon‒Intron Structures for NPF Genes in Apple and Other Species
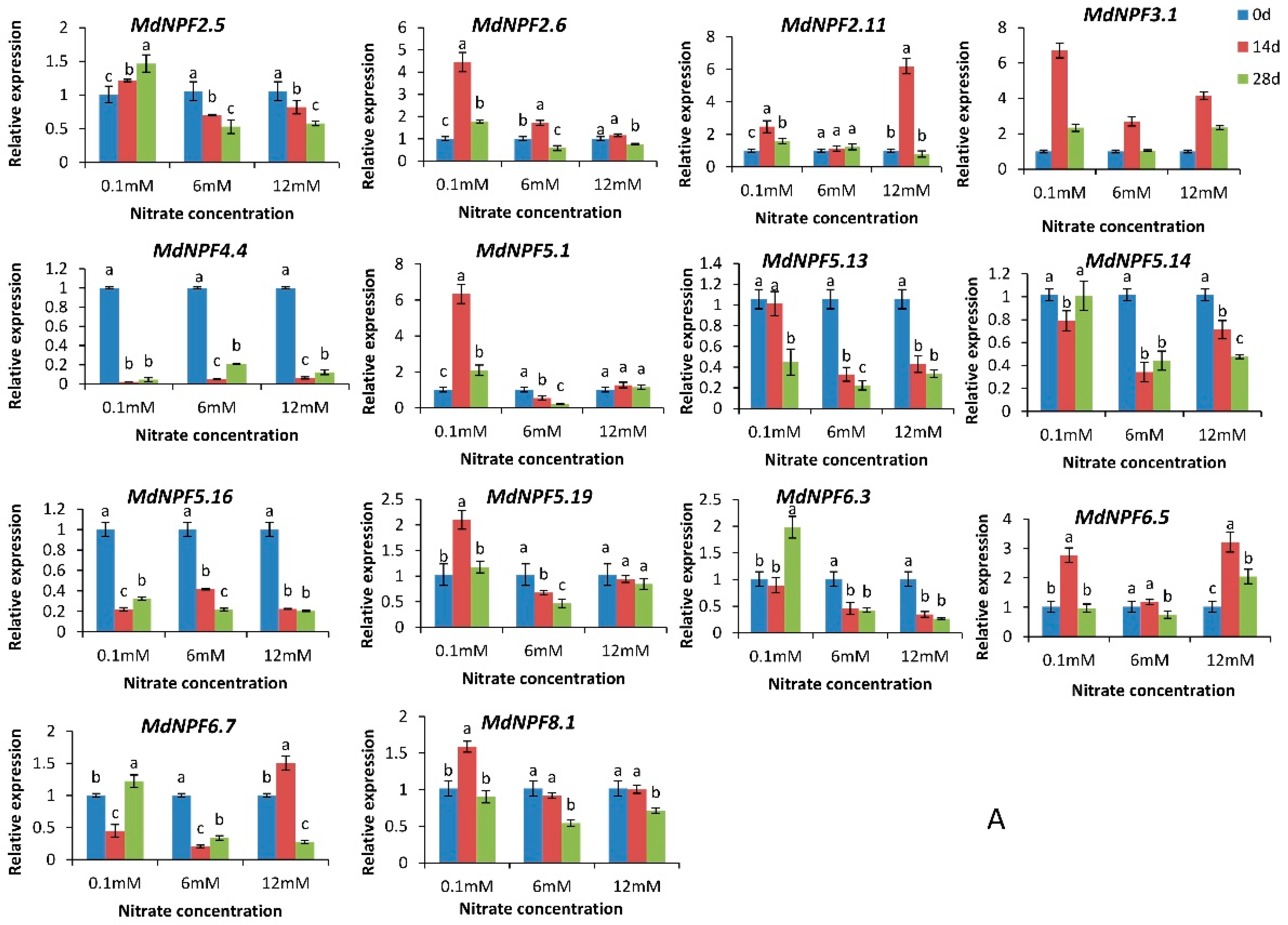
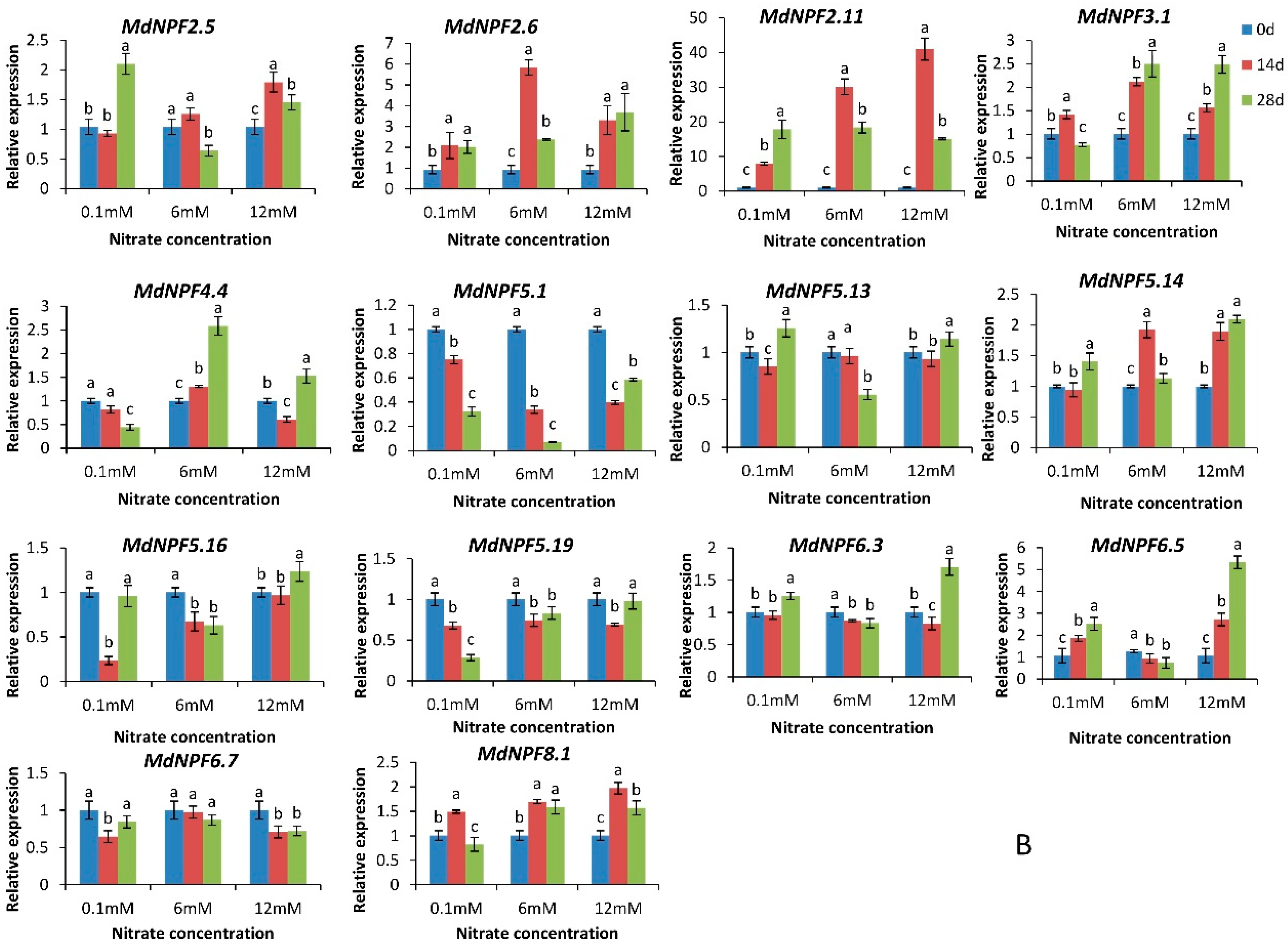
2.6. Analysis of Expression for 14 MdNPFs in Response to Different Nitrate Concentrations
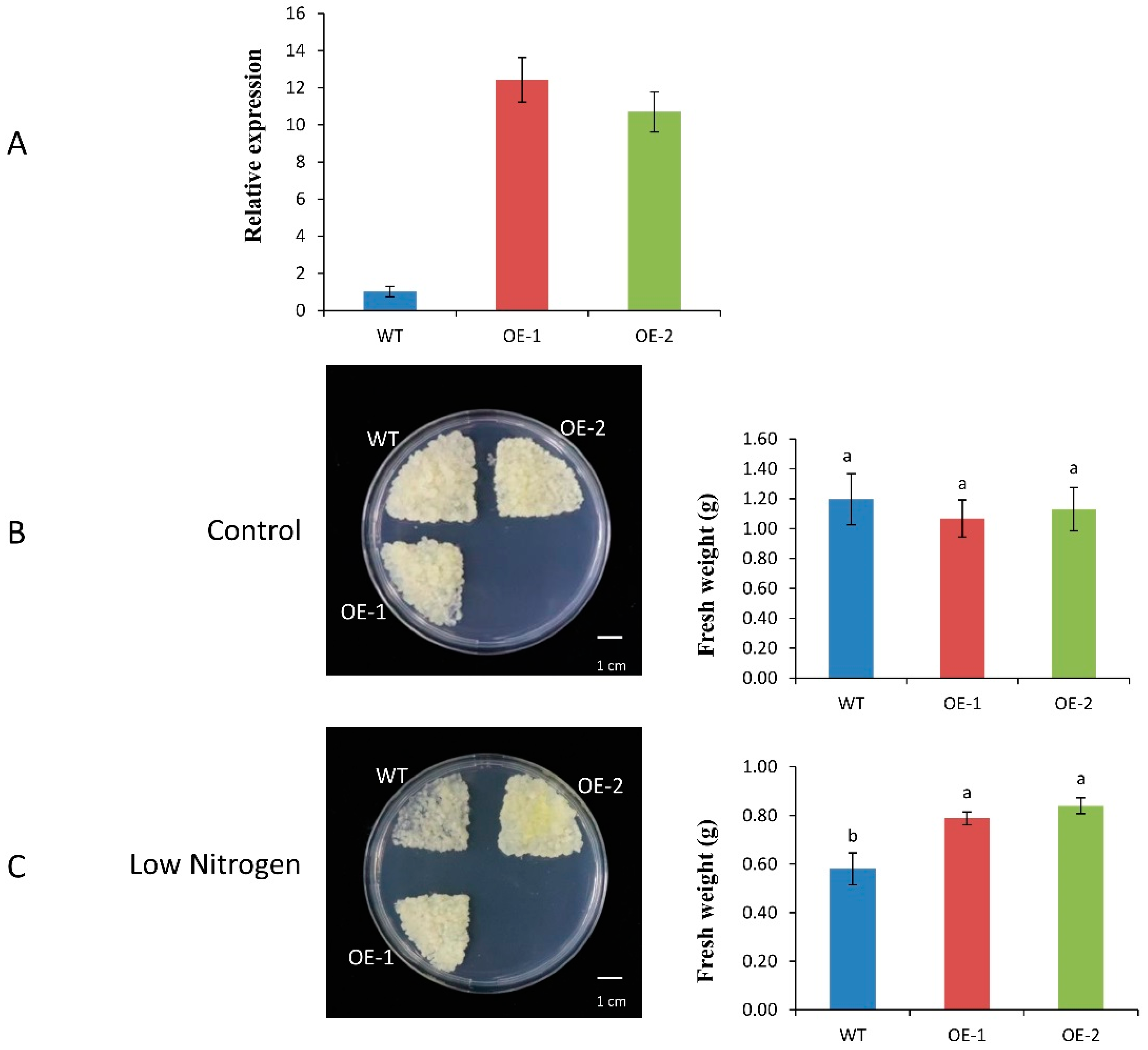
2.7. Effect of Low-Nitrogen Treatment on Growth by Apple Calli Tissue
3. Discussion
4. Materials and Methods
4.1. Identification of Apple NPF Genes
4.2. Sequence Alignment and Phylogenetic Analysis
4.3. Analyses of Exon‒Intron Structure and Genome Distribution
4.4. Sequence Logo and Prediction of TM Regions
4.5. Plant Materials and Nitrogen Treatments
4.6. Quantitative Real Time RT-PCR (qRT-PCR) and Gene-Cloning
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- O’Brien, J.A.; Vega, A.; Bouguyon, E.; Krouk, G.; Gojon, A.; Coruzzi, G.; Gutiérrez, R.A. Nitrate transport, sensing, and responses in plants. Mol. Plant. 2016, 9, 837–856. [Google Scholar] [CrossRef] [PubMed]
- Krapp, A. Plant nitrogen assimilation and its regulation: A complex puzzle with missing pieces. Curr. Opin. Plant Biol. 2015, 25, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Willmann, A.; Thomfohrde, S.; Haensch, R.; Nehls, U. The poplar NRT2 gene family of high affinity nitrate importers: Impact of nitrogen nutrition and ectomycorrhiza formation. Environ. Exp. Bot. 2014, 108, 79–88. [Google Scholar] [CrossRef]
- Hu, B.; Wang, W.; Ou, S.J.; Tang, J.Y.; Li, H.; Che, R.H.; Zhang, Z.H.; Chai, X.Y.; Wang, H.R.; Wang, Y.Q.; et al. Variation in NRT1.1B contributes to nitrate-use divergence between rice subspecies. Nat. Genet. 2015, 47, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Hu, B.; Yuan, D.Y. Expression of the nitrate transporter gene OsNRT1.1A/OsNPF6.3 confers high yield and early maturation in rice. Plant Cell. 2018, 30, 638–651. [Google Scholar] [CrossRef] [PubMed]
- Claussen, W. Growth, water use efficiency, and proline content of hydroponically grown tomato plants as affected by nitrogen source and nutrient concentration. Plant Soil. 2002, 247, 199–209. [Google Scholar] [CrossRef]
- Huang, L.L.; Li, M.J.; Zhou, K.; Sun, T.T.; Hu, L.Y.; Li, C.Y.; Ma, F.W. Uptake and metabolism of ammonium and nitrate in response to drought stress in Malus prunifolia. Plant Physiol. Biochem. 2018, 127, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Feldmann, K.A. The herbicide sensitivity gene CHL1 of Arabidopsis encodes a nitrate-inducible nitrate transporter. Cell 1993, 72, 705–713. [Google Scholar]
- Mounier, E.; Pervent, M.; Ljung, K.; Gojon, A.; Nacry, P. Auxin-mediated nitrate signalling by NRT1.1 participates in the adaptive response of Arabidopsis root architecture to the spatial heterogeneity of nitrate availability. Plant Cell Environ. 2014, 37, 162–174. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.H.; Kuo, H.F.; Canivenc, G.; Lin, C.S.; Lepetit, M.; Hsu, P.K.; Tillard, P.; Lin, H.L.; Wang, Y.Y.; Tsai, C.B.; et al. Mutation of the Arabidopsis NRT1.5 nitrate transporter causes defective root to-shoot nitrate transport. Plant Cell 2008, 20, 2514–2528. [Google Scholar] [CrossRef] [PubMed]
- Almagro, A.; Lin, S.H.; Tsay, Y.F. Characterization of the Arabidopsis nitrate transporter NRT1.6 reveals a role of nitrate in early embryo development. Plant Cell 2008, 20, 3289–3299. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, I.T.; Skurray, R.A. The POT family of transport proteins. Trends Biochem. Sci. 1994, 19, 404. [Google Scholar] [CrossRef]
- Steiner, H.Y.; Naider, F.; Becker, J.M. The PTR family: A new group of peptide transporters. Mol. Microbiol. 1995, 16, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.B.; Huang, J.Y.; Yu, H.H. Genomic survey, characterization and expression profile analysis of the peptide transporter family in rice (Oryzasativa L.). BMC Plant Biol. 2010, 10, 92. [Google Scholar] [CrossRef] [PubMed]
- Tsay, Y.F.; Chiu, C.C.; Tsai, C.B.; Ho, C.H.; Hsu, P.K. Nitrate transporters and peptide transporters. FEBS Lett. 2007, 581, 2290–2300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karim, S.; Lundh, D.; Holmström, K.O.; Mandal, A.; Pirhonen, M. Structural and functional characterization of AtPTR3, a stress-induced peptide transporter of Arabidopsis. J. Mol. Model. 2005, 11, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Karim, S.; Holmström, K.O.; Mandal, A.; Dahl, P.; Hohmann, S.; Brader, G.; Palva, E.T.; Pirhonen, M. AtPTR3 a wound-induced peptide transporter needed for defence against virulent bacterial pathogens in Arabidopsis. Planta 2007, 225, 1431–1445. [Google Scholar] [CrossRef] [PubMed]
- Komarova, N.Y.; Thor, K.; Gubler, A.; Meier, S.; Dietrich, D.; Weichert, A.; Grotemeyer, M.S.; Tegeder, M.; Rentsch, D. AtPTR1 and AtPTR5 transport dipeptides in planta. Plant Physiol. 2008, 148, 856–869. [Google Scholar] [CrossRef] [PubMed]
- Léran, S.; Varala, K.; Boyer, J.C.; Chiurazzi, M.; Crawford, N.; Daniel-Vedele, F.; David, L.; Dickstein, R.; Fernandez, E.; Forde, B.; et al. A unified nomenclature of NITRATE TRANSPORTER 1/PEPTIDE TRANSPORTER family members in plants. Trends Plant Sci. 2014, 19, 5–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jørgensen, M.E.; Xu, D.Y.; Crocoll, C.; Ramírez, D.; Motawia, M.S.; Olsen, C.E.; Nour-Eldin, H.H.; Kiba, T.; Feria-Bourrellier, A.B.; Lafouge, F.; et al. The Arabidopsis nitrate transporter NRT2.4 plays a double role in roots and shoots of nitrogen-starved plants. Plant Cell 2012, 24, 245–258. [Google Scholar]
- Nour-Eldin, H.H.; Andersen, T.G.; Burow, M.; Madsen, S.R.; Jørgensen, M.E.; Olsen, C.E.; Dreyer, I.; Hedrich, R.; Geiger, D.; Halkier, B.A. NRT/PTR transporters are essential for translocation of glucosinolate defence compounds to seeds. Nature 2012, 488, 531–534. [Google Scholar] [CrossRef] [PubMed]
- Boursiac, Y.; Léran, S.; Corratgé-Faillie, C.; Gojon, A.; Krouk, G.; Lacombe, B. ABA transport and transporters. Trends Plant Sci. 2013, 18, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Tal, I.; Zhang, Y.; Jorgensen, M.E.; Pisanty, O.; Barbosa, I.C.R.; Zourelidou, M.; Regnault, T.; Crocoll, C.; Olsen, C.E.; Weinstain, R.; et al. The Arabidopsis NPF3 protein is a GA transporter. Nat. Commun. 2016, 7, 11486–11496. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.K.; Tsay, Y.F. Two phloem nitrate transporters, NRT1.11 and NRT1.12, are important for redistributing xylem-borne nitrate to enhance plant growth. Plant Physiol. 2013, 163, 844–856. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.B.; Gong, J.M. The Arabidopsis ethylene/jasmonic acid-NRT signaling module coordinates nitrate reallocation and the trade-off between growth and environmental adaptation. Plant Cell 2014, 26, 3984–3998. [Google Scholar] [CrossRef] [PubMed]
- Léran, S.; Garg, B.; Boursiac, Y.; Corratgé-Faillie, C.; Brachet, C.; Tillard, P.; Gojon, A.; Lacombe, B. AtNPF5.5, a nitrate transporter affecting nitrogen accumulation in Arabidopsis embryo. Sci. Rep. 2015, 5, 7962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pike, S.; Gao, F.; Kim, M.J.; Kim, S.H.; Schachtman, D.P.; Gassmann, W. Members of the NPF3 transporter subfamily encode pathogen-inducible nitrate/nitrite transporters in grapevine and Arabidopsis. Plant Cell Physiol. 2014, 55, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Li, F.X.; Liu, D.; Jiang, C.H.; Cui, L.J.; Shen, L.L.; Liu, G.S.; Yang, A.G. Dynamic expression analysis of early response genes induced by potato virus Y in PVY-resistant Nicotiana tabacum. Plant Cell Rep. 2017, 36, 297–311. [Google Scholar] [CrossRef] [PubMed]
- Chopin, F.; Orsel, M.; Dorbe, M.F.; Chardon, F.; Truong, H.N.; Miller, A.J.; Krapp, A.; Daniel-Vedele, F. The Arabidopsis AtNRT2.7 nitrate transporter controls nitrate content in seeds. Plant Cell 2007, 19, 1590–1602. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.C.; Lin, C.S.; Hsu, P.K.; Lin, S.H.; Tsay, Y.F. The Arabidopsis nitrate transporter NRT1.7, expressed in phloem, is responsible for source-to-sink remobilization of nitrate. Plant Cell 2009, 21, 2750–2761. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.D.; Fan, X.R.; Wei, J. Rice nitrate transporter OsNPF2.4 functions in low-affinity acquisition and long-distance transport. J. Exp. Bot. 2015, 66, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.R.; Tang, Z.; Tan, Y.; Zhang, Y.; Luo, B.; Yang, M.; Lian, X.M.; Shen, Q.R.; Miller, A.J.; Xu, G.H. Overexpression of a pH-sensitive nitrate transporter in rice increases crop yields. Proc. Natl. Acad. Sci. USA 2016, 113, 7118–7123. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.Y.; Sun, J.Y.; Tian, Z.W.; Hu, H.; Michel, E.J.S.; Gao, J.W.; Jiang, D.; Cao, W.X.; Dai, T.B. Root extension and nitrate transporter up-regulation induced by nitrogen deficiency improves nitrogen status and plant growth at the seedling stage of winter wheat (Triticum aestivum L.). Environ. Exp. Bot. 2017, 141, 28–40. [Google Scholar] [CrossRef]
- Luo, J.; Li, H.; Liu, T.X.; Polle, A.; Peng, C.H.; Luo, Z.B. Nitrogen metabolism of two contrasting poplar species during acclimation to limiting nitrogen availability. J. Exp. Bot. 2013, 64, 4207–4224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valkov, V.T.; Rogato, A.; Alves, L.M.; Sol, S.; Noguero, M.; Léran, S.; Lacombe, B.; Chiurazzi, M. The nitrate transporter family protein LjNPF8.6 controls the N-fixing nodule activity. Plant Physiol. 2017, 175, 1269–1282. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.L.; Yi, H.Y.; Bao, J.; Gong, J.M. LeNRT2.3 functions in nitrate acquisition and long-distance transport in tomato. FEBS Lett. 2015, 589, 1072–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.L.; Ma, W.J.; Song, J.Y.; Lu, M.; Rahman, S.U.; Bui, T.T.X.; Vu, D.D.; Zheng, H.F.; Wang, J.H.; Zhang, Y. Physiological and transcriptional responses of Catalpa bungei to drought stress under sufficient- and deficient-nitrogen conditions. Tree Physiol. 2017, 247, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Saier, M.H., Jr. Families of transmembrane transporters selective for amino acids and their derivatives. Microbiology 2000, 146, 1775–1795. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Liang, D.; Wang, P.; Liu, J.Y.; Ma, F.W. Genome-wide analysis and expression profiling of the DREB transcription factor gene family in Malus under abiotic stress. Mol. Genet. Genom. 2012, 287, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.H.; Zhang, S.Z.; Wang, R.K.; Zhang, R.F.; Hao, Y.J. Genome wide analysis of the apple MYB transcription factor family allows the identification of MdoMYB121 gene conferring abiotic stress tolerance in plants. PLoS ONE 2013, 8, e69955. [Google Scholar]
- Tian, Y.; Dong, Q.L.; Ji, Z.R.; Chi, F.M.; Cong, P.H.; Zhou, Z.S. Genome-wide identification and analysis of the MADS-box gene family in apple. Gene 2015, 555, 277–290. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.T.; Li, M.J.; Shao, Y.; Yu, L.Y.; Ma, F.W. Comprehensive genomic identification and expression analysis of the phosphate transporter (PHT) gene family in apple. Front. Plant Sci. 2017, 8, 426. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Gong, X.Q.; Ma, F.W. Genome-wide identification of the radiation sensitivity protein-23 (RAD23) family members in apple (Malus×domestica Borkh.) and expression analysis of their stress responsiveness. J. Integr. Agric. 2017, 16, 820–827. [Google Scholar] [CrossRef]
- Zhou, K.; Hu, L.Y.; Li, P.M.; Gong, X.Q.; Ma, F.W. Genome-wide identification of glycosyltransferases converting phloretin to phloridzin in Malus species. Plant Sci. 2017, 265, 131–145. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Qin, Y.; Zou, Y.J.; Ma, F.W. Genome-wide identification and expression profiling of the SnRK2 gene family in Malus prunifolia. Gene 2014, 552, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.B.; Ji, Z.R.; Chi, F.M.; Qiao, Z.; Xu, C.N.; Zhang, J.X.; Dong, Q.L.; Zhou, Z.S. Bioinformatics and expression analysis of the WRKY gene family in apple. Scient. Agric. Sin. 2015, 48, 3221–3238. [Google Scholar]
- Meng, D.; Li, Y.Y.; Bai, Y.; Li, M.J.; Cheng, L.L. Genome-wide identification and characterization of WRKY transcriptional factor family in apple and analysis of their responses to waterlogging and drought stress. Plant Physiol. Biochem. 2016, 103, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Euring, D.; Volmer, K.; Janz, D.; Polle, A. The nitrate transporter (NRT) gene family in poplar. PLoS ONE 2013, 8, e72126. [Google Scholar] [CrossRef] [PubMed]
- Pellizzaro, A.; Alibert, B.; Planchet, E.; Limami, A.M.; Morère-Le Paven, M.C. Nitrate transporters: An overview in legumes. Planta 2017, 246, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Plett, D.; Toubia, J.; Garnett, T.; Tester, M.; Kaiser, B.N.; Baumann, U. Dichotomy in the NRT gene families of dicots and grass species. PLoS ONE 2010, 5, e72126. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.B.; Krishnakumar, V.; Bidwell, S.; Rosen, B.; Chan, A.; Zhou, S.G.; Gentzbittel, L.; Childs, K.L.; Yandell, M.; Gundlach, H.; et al. An improved genome release (version Mt4.0) for the model legume Medicago truncatula. BMC Genom. 2014, 15, 312. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yu, M.; Du, X.Q.; Wang, Z.F.; Wu, W.H.; Quintero, F.J.; Jin, X.H.; Li, H.D.; Wang, Y. NRT1.5/NPF7.3 functions as a proton-coupled H+/K+ antiporter for K+ loading into the xylem in Arabidopsis. Plant Cell 2017, 29, 2016–2026. [Google Scholar] [CrossRef] [PubMed]
- Drechsler, N.; Courty, P.E.; Brulé, D.; Kunze, R. Identification of arbuscular mycorrhiza-inducible Nitrate Transporter 1/Peptide Transporter Family (NPF) genes in rice. Mycorrhiza 2018, 28, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.H.; Tsay, Y.F. Switching between the two action modes of the dual-affinity nitrate transporter CHL1 by phosphorylation. EMBO J. 2003, 22, 1005–1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, K.H.; Huang, C.Y.; Tsay, Y.F. CHL1 is a dual-affinity nitrate transporter of Arabidopsis involved in multiple phases of nitrate uptake. Plant Cell 1999, 11, 865–874. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Li, L.; Luan, S. Immunophilins and parvulins. Superfamily of peptidyl prolyl isomerases in Arabidopsis. Plant Physiol. 2004, 134, 1248–1267. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING v10: Protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015, 43, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Bailly, A.; Sovero, V.; Vincenzetti, V.; Santelia, D.; Bartnik, D.; Koening, B.W.; Mancuso, S.; Martinoia, E.; Geisler, M. Modulation of P-glycoproteins by auxin transport inhibitors is mediated by interaction with immunophilins. J. Biol. Chem. 2008, 283, 21817–21826. [Google Scholar] [CrossRef] [PubMed]
- Crooks, G.E.; Hon, G.; Chandonia, J.M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.G.; Li, M.; Luo, H.; Dong, Q.L.; Yao, Y.X.; You, C.X.; Hao, Y.J. Molecular cloning and functional characterization of MdSOS2 reveals its involvement in salt tolerance in apple callus and Arabidopsis. Plant Cell Rep. 2012, 31, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Dong, Q.L.; Zhao, S.; Duan, D.Y.; Tian, Y.; Wang, Y.P.; Mao, K.; Zhou, Z.S.; Ma, F.W. Structural and functional analyses of genes encoding VQ proteins in apple. Plant Sci. 2018, 272, 208–219. [Google Scholar] [CrossRef] [PubMed]
- Perini, P.; Pasquali, G.; Margis-Pinheiro, M.; Oliviera, P.R.D.; Revers, L.F. Reference genes for transcriptional analysis of flowering and fruit ripening stages in apple (Malus × domestica Borkh.). Mol. Breed. 2014, 34, 829–842. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID a | Chromosome Location | ORF | Protein Length (aa) | MW (kDa) | Theoretical Isoeletrical Point (pI) | GenBank Accession Numbers of Cloned Gene |
|---|---|---|---|---|---|---|---|
| MdNPF1 | MD15G1190800 | Chr15:15008548-15011039 | 1779 | 593 | 65.21 | 8.90 | |
| MdNPF2.1 | MD11G1122200 | Chr11:11243604-11246153 | 1656 | 552 | 60.36 | 8.14 | |
| MdNPF2.2 | MD11G1121600 | Chr11:11225429-11227624 | 1404 | 468 | 51.09 | 8.99 | |
| MdNPF2.3 | MD07G1180500 | Chr07:25970318-25972596 | 1539 | 513 | 56.69 | 9.27 | |
| MdNPF2.4 | MD01G1112600 | Chr01:22682711-22685617 | 1662 | 554 | 60.53 | 9.25 | |
| MdNPF2.5 | MD11G1122500 | Chr11:11284715-11290091 | 1698 | 566 | 61.85 | 8.86 | MG021338 |
| MdNPF2.6 | MD03G1108900 | Chr03:9496734-9500175 | 1812 | 604 | 66.37 | 8.73 | MG021345 |
| MdNPF2.7 | MD03G1108700 | Chr03:9467766-9470916 | 1683 | 561 | 61.53 | 8.96 | |
| MdNPF2.8 | MD06G1186800 | Chr06:32435102-32437373 | 1809 | 603 | 67.17 | 9.61 | |
| MdNPF2.9 | MD14G1193200 | Chr14:28393718-28396166 | 1866 | 622 | 69.34 | 8.88 | |
| MdNPF2.10 | MD04G1137500 | Chr04:22509802-22511767 | 1764 | 588 | 64.55 | 8.76 | |
| MdNPF2.11 | MD14G1193100 | Chr14:28376948-28379737 | 1746 | 482 | 64.43 | 9.00 | MG021332 |
| MdNPF2.12 | MD06G1186500 | Chr06:32427308-32429277 | 1299 | 433 | 48.04 | 8.86 | |
| MdNPF2.13 | MD16G1080100 | Chr16:5611393-5619434 | 1845 | 615 | 68.20 | 8.44 | |
| MdNPF2.14 | MD16G1079900 | Chr16:5602664-5605755 | 1818 | 606 | 66.73 | 8.14 | |
| MdNPF2.15 | MD16G1080000 | Chr16:5608051-5610469 | 1737 | 579 | 63.71 | 7.82 | |
| MdNPF3.1 | MD13G1043200 | Chr13:3030935-3034949 | 1740 | 580 | 64.10 | 8.61 | MG021337 |
| MdNPF3.2 | MD13G1043100 | Chr13:3013976-3017053 | 1776 | 592 | 65.47 | 8.61 | |
| MdNPF3.3 | MD16G1044000 | Chr16:3127680-3131016 | 1743 | 581 | 64.24 | 8.34 | |
| MdNPF4.1 | MD05G1164400 | Chr05:29316397-29322734 | 1308 | 436 | 48.37 | 7.39 | |
| MdNPF4.2 | MD10G1153900 | Chr10:24150970-24153718 | 1605 | 535 | 59.31 | 9.33 | |
| MdNPF4.3 | MD10G1154000 | Chr10:24171779-24174663 | 1602 | 534 | 59.02 | 8.26 | |
| MdNPF4.4 | MD08G1248200 | Chr08:31226687-31231330 | 1842 | 614 | 67.84 | 8.30 | MG021342 |
| MdNPF4.5 | MD15G1443100 | Chr15:54322869-54325021 | 1482 | 494 | 54.88 | 8.96 | |
| MdNPF4.6 | MD05G1000900 | Chr05:293868-298577 | 1845 | 615 | 68.08 | 8.54 | |
| MdNPF4.7 | MD13G1079300 | Chr13:5562578-5567885 | 1761 | 587 | 64.77 | 8.58 | |
| MdNPF4.8 | MD16G1079100 | Chr16:5547983-5553360 | 1761 | 587 | 64.85 | 8.70 | |
| MdNPF4.9 | MD14G1194100 | Chr14:28528146-28530941 | 1758 | 586 | 64.70 | 8.26 | |
| MdNPF4.10 | MD08G1040500 | Chr08:2990042-2993173 | 1668 | 556 | 61.30 | 8.22 | |
| MdNPF4.11 | MD04G1184500 | Chr04:27539356-27541647 | 1836 | 612 | 68.06 | 9.15 | |
| MdNPF4.12 | MD12G1197700 | Chr12:27888187-27890493 | 1653 | 551 | 61.38 | 8.67 | |
| MdNPF4.13 | MD10G1271800 | Chr10:36356975-36363499 | 1779 | 593 | 66.11 | 8.08 | |
| MdNPF4.14 | MD05G1293900 | Chr05:42659455-42665121 | 1809 | 603 | 67.05 | 8.58 | |
| MdNPF5.1 | MD07G1230600 | Chr07:30498119-30501840 | 1776 | 592 | 66.07 | 8.76 | MG021340 |
| MdNPF5.2 | MD05G1192100 | Chr05:32036536-32040692 | 1809 | 603 | 67.23 | 9.30 | |
| MdNPF5.3 | MD16G1224200 | Chr16:22556239-22559806 | 1644 | 548 | 61.45 | 8.57 | |
| MdNPF5.4 | MD13G1218900 | Chr13:21148526-21157623 | 1806 | 602 | 67.12 | 9.36 | |
| MdNPF5.5 | MD05G1342600 | Chr05:46297101-46299829 | 1779 | 593 | 66.26 | 9.22 | |
| MdNPF5.6 | MD07G1039100 | Chr07:3293256-3295279 | 873 | 291 | 32.36 | 9.39 | |
| MdNPF5.7 | MD07G1038900 | Chr07:3272734-3276872 | 1695 | 565 | 62.35 | 9.08 | |
| MdNPF5.8 | MD07G1038800 | Chr07:3237655-3247764 | 1731 | 577 | 63.70 | 8.82 | |
| MdNPF5.9 | MD07G1038600 | Chr07:3176293-3185619 | 1695 | 565 | 62.63 | 9.01 | |
| MdNPF5.10 | MD07G1039600 | Chr07:3353966-3358684 | 1731 | 577 | 63.97 | 8.71 | |
| MdNPF5.11 | MD04G1148300 | Chr04:23666705-23668537 | 1635 | 545 | 60.03 | 6.25 | |
| MdNPF5.12 | MD04G1138500 | Chr04:22623404-22627516 | 1770 | 590 | 65.59 | 8.62 | |
| MdNPF5.13 | MD12G1153900 | Chr12:23405312-23409371 | 1770 | 590 | 65.19 | 8.93 | MG021339 |
| MdNPF5.14 | MD07G1205700 | Chr07:28355985-28360202 | 1794 | 598 | 67.03 | 8.77 | MG021344 |
| MdNPF5.15 | MD01G1141500 | Chr01:25051572-25054193 | 1785 | 595 | 66.39 | 9.24 | |
| MdNPF5.16 | MD17G1041000 | Chr17:2979487-2982673 | 1632 | 544 | 60.48 | 8.35 | MG021336 |
| MdNPF5.17 | MD09G1040700 | Chr09:2607173-2618823 | 1581 | 527 | 58.64 | 8.79 | |
| MdNPF5.18 | MD08G1218300 | Chr08:28077601-28080578 | 1080 | 360 | 40.06 | 5.25 | |
| MdNPF5.19 | MD15G1406700 | Chr15:50693334-50695520 | 1317 | 439 | 48.70 | 6.36 | MG021331 |
| MdNPF5.20 | MD15G1406500 | Chr15:50682947-50684829 | 1377 | 459 | 50.92 | 5.76 | |
| MdNPF5.21 | MD07G1039200 | Chr07:3295281-3306861 | 825 | 275 | 29.88 | 8.94 | |
| MdNPF6.1 | MD08G1022500 | Chr08:1648693-1654146 | 1551 | 517 | 57.45 | 9.00 | |
| MdNPF6.2 | MD15G1019900 | Chr15:1155213-1159411 | 1869 | 623 | 69.53 | 8.46 | |
| MdNPF6.3 | MD16G1142100 | Chr16:10938991-10941873 | 1914 | 638 | 70.17 | 8.56 | MG021341 |
| MdNPF6.4 | MD13G1131800 | Chr13:10003867-10006664 | 1914 | 638 | 70.08 | 7.70 | |
| MdNPF6.5 | MD15G1173800 | Chr15:13572779-13576346 | 1746 | 582 | 63.62 | 9.30 | MG021346 |
| MdNPF6.6 | MD04G1086400 | Chr04:12553185-12555016 | 1011 | 337 | 36.83 | 8.33 | |
| MdNPF6.7 | MD17G1103000 | Chr17:8745481-8748650 | 1773 | 591 | 65.11 | 9.24 | MG021333 |
| MdNPF7.1 | MD11G1017300 | Chr11:1392309-1395225 | 1866 | 622 | 67.81 | 5.82 | |
| MdNPF7.2 | MD03G1016700 | Chr03:1321170-1324060 | 2022 | 674 | 73.81 | 7.30 | |
| MdNPF7.3 | MD03G1016400 | Chr03:1307373-1311107 | 1782 | 594 | 65.21 | 6.59 | |
| MdNPF7.4 | MD07G1082700 | Chr07:8103711-8110134 | 1815 | 605 | 67.04 | 7.80 | |
| MdNPF7.5 | MD02G1228800 | Chr02:27110155-27115346 | 1815 | 605 | 67.22 | 7.64 | |
| MdNPF7.6 | MD06G1029400 | Chr06:3504379-3509435 | 1791 | 597 | 66.37 | 7.89 | |
| MdNPF7.7 | MD16G1277800 | Chr16:37688341-37693389 | 1788 | 596 | 66.44 | 6.71 | |
| MdNPF8.1 | MD12G1160700 | Chr12:24039618-24042029 | 1707 | 569 | 63.32 | 8.60 | MG021334 |
| MdNPF8.2 | MD04G1147500 | Chr04:23563075-23568217 | 2061 | 687 | 76.60 | 8.76 | |
| MdNPF8.3 | MD11G1081100 | Chr11:6913320-6915864 | 1728 | 576 | 63.96 | 8.11 | |
| MdNPF8.4 | MD11G1081200 | Chr11:6931888-6934707 | 1749 | 583 | 64.77 | 8.62 | |
| MdNPF8.5 | MD16G1010600 | Chr16:814341-817396 | 1758 | 586 | 64.39 | 7.18 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Liu, C.; Dong, Q.; Huang, D.; Li, C.; Li, P.; Ma, F. Genome-Wide Identification and Analysis of Apple NITRATE TRANSPORTER 1/PEPTIDE TRANSPORTER Family (NPF) Genes Reveals MdNPF6.5 Confers High Capacity for Nitrogen Uptake under Low-Nitrogen Conditions. Int. J. Mol. Sci. 2018, 19, 2761. https://doi.org/10.3390/ijms19092761
Wang Q, Liu C, Dong Q, Huang D, Li C, Li P, Ma F. Genome-Wide Identification and Analysis of Apple NITRATE TRANSPORTER 1/PEPTIDE TRANSPORTER Family (NPF) Genes Reveals MdNPF6.5 Confers High Capacity for Nitrogen Uptake under Low-Nitrogen Conditions. International Journal of Molecular Sciences. 2018; 19(9):2761. https://doi.org/10.3390/ijms19092761
Chicago/Turabian StyleWang, Qian, Changhai Liu, Qinglong Dong, Dong Huang, Cuiying Li, Pengmin Li, and Fengwang Ma. 2018. "Genome-Wide Identification and Analysis of Apple NITRATE TRANSPORTER 1/PEPTIDE TRANSPORTER Family (NPF) Genes Reveals MdNPF6.5 Confers High Capacity for Nitrogen Uptake under Low-Nitrogen Conditions" International Journal of Molecular Sciences 19, no. 9: 2761. https://doi.org/10.3390/ijms19092761