Phosphatidylethanolamine Induces an Antifibrotic Phenotype in Normal Human Lung Fibroblasts and Ameliorates Bleomycin-Induced Lung Fibrosis in Mice

 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Effect of Different Phospholipid Components of Surfactant on Collagen mRNA Expression
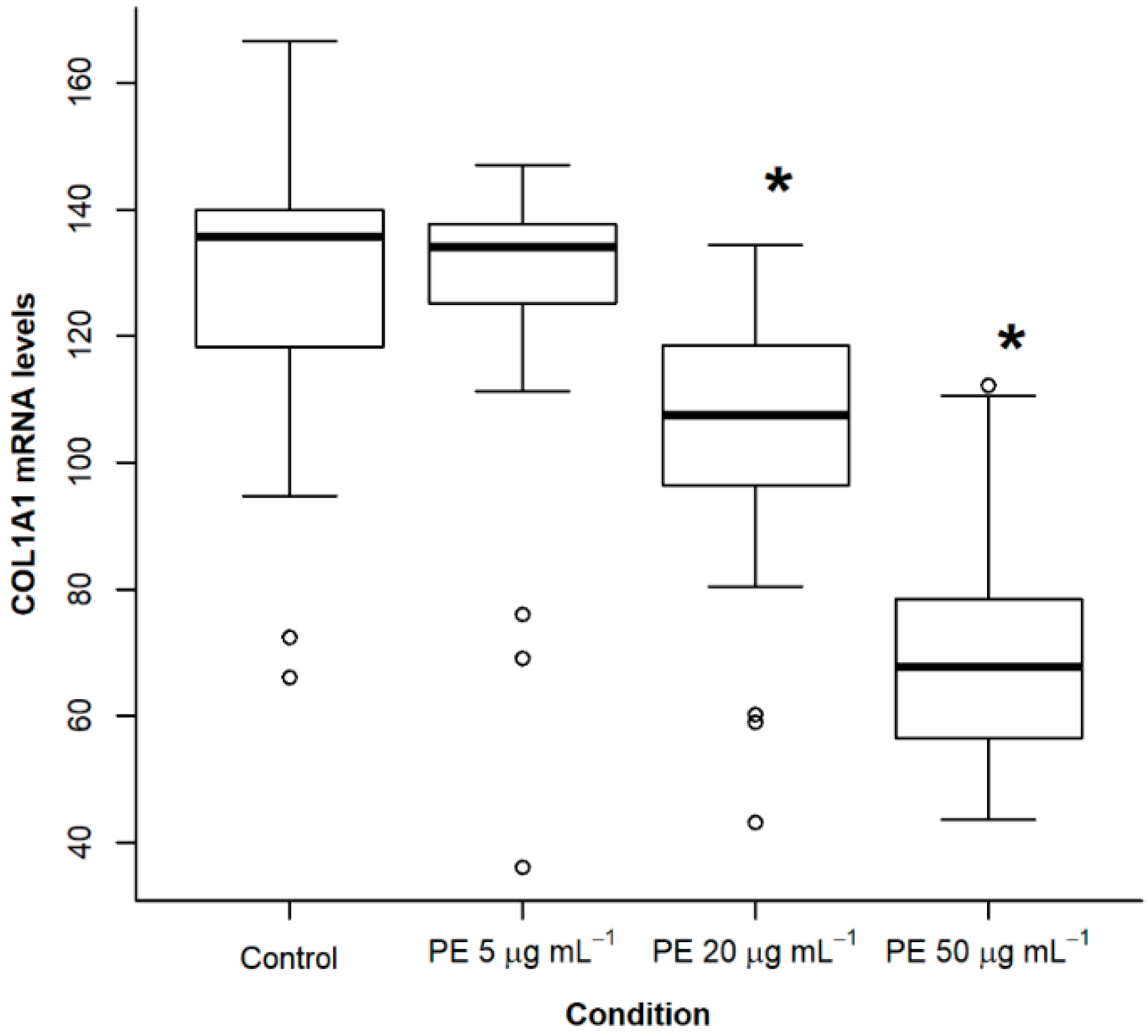
2.2. Treatment of NHLF with PE Causes a Dose- and Time-Dependent Decrease of Collagen Expression
2.3. Effect of PE on Fibroblast Apoptosis
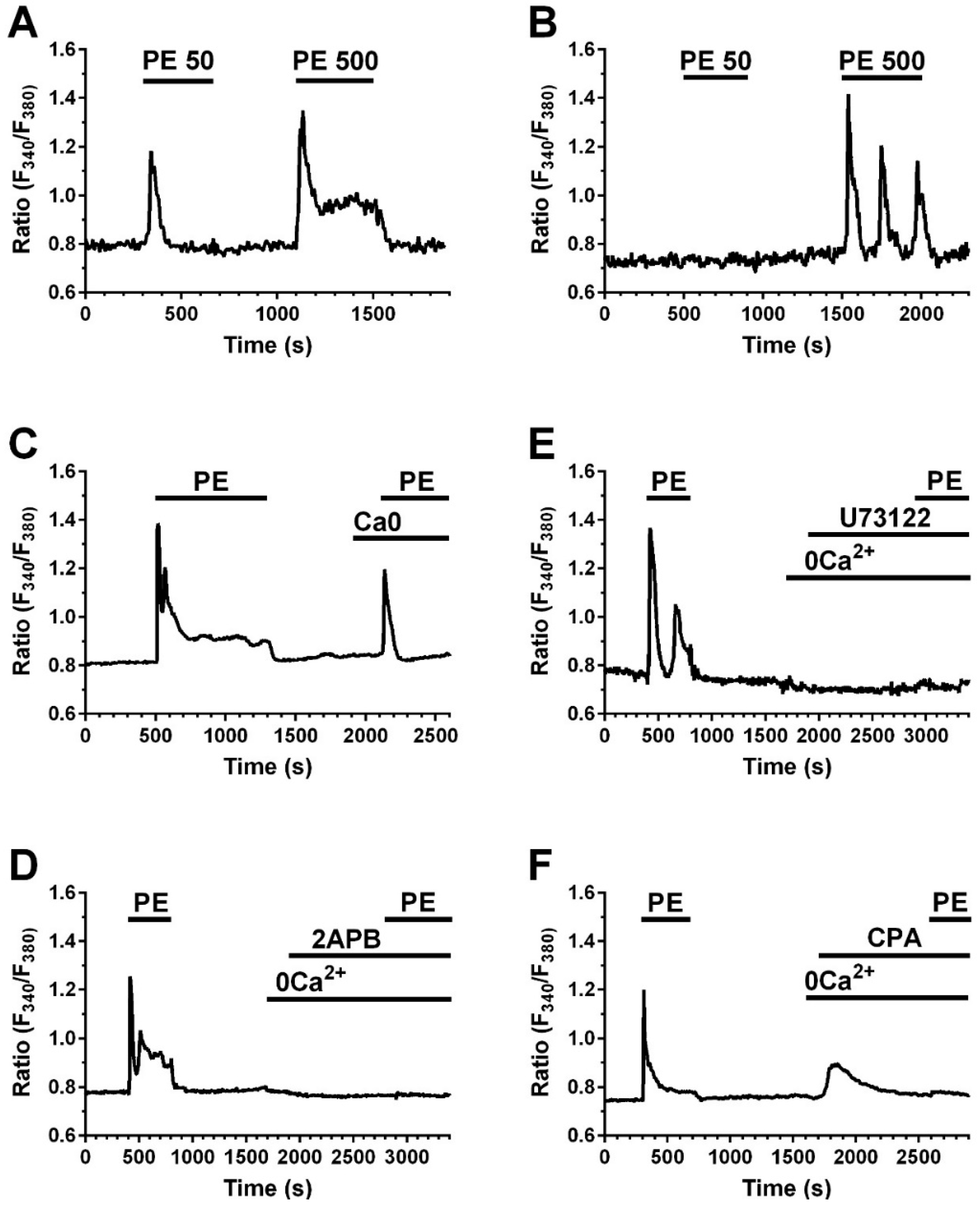
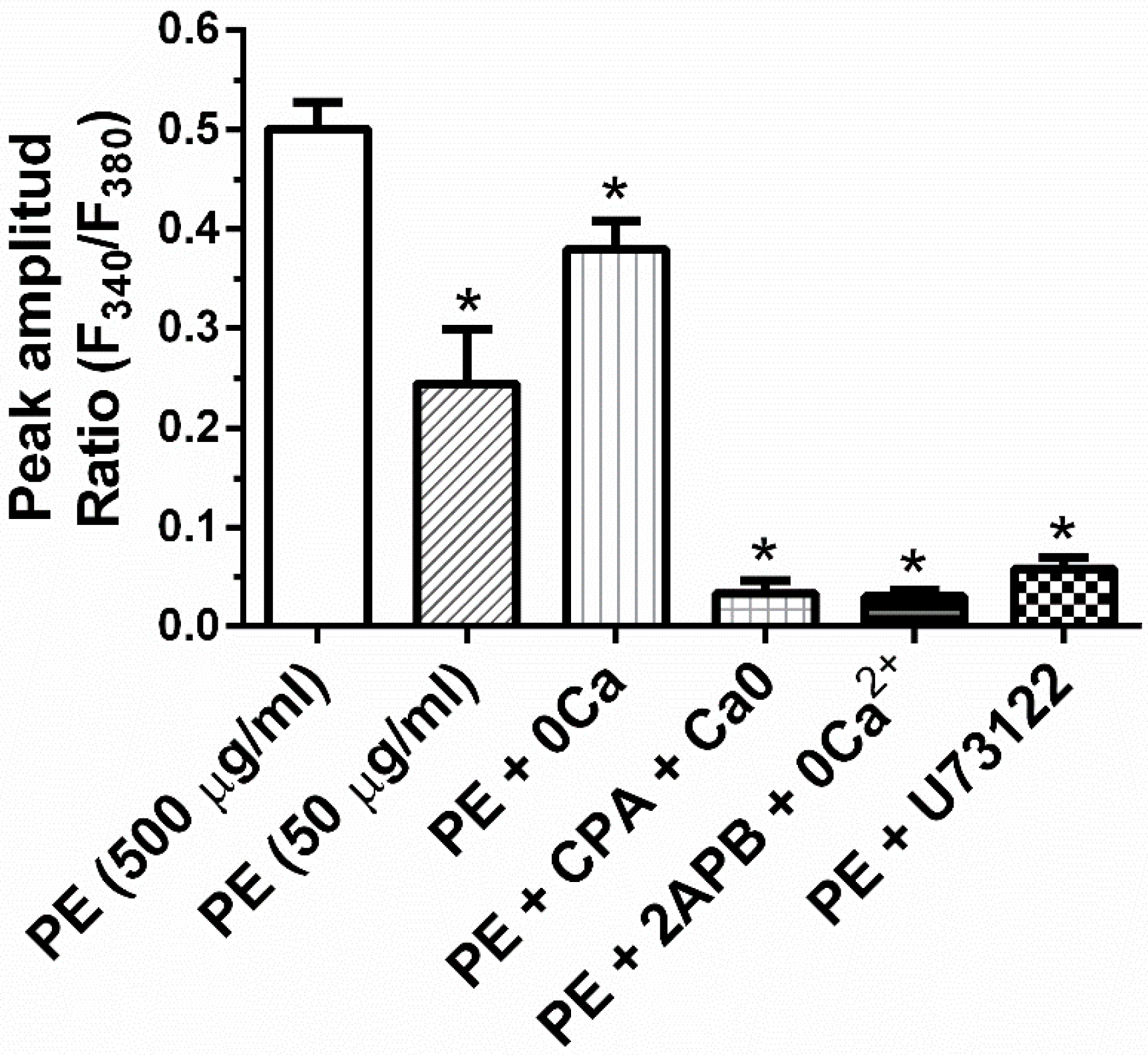
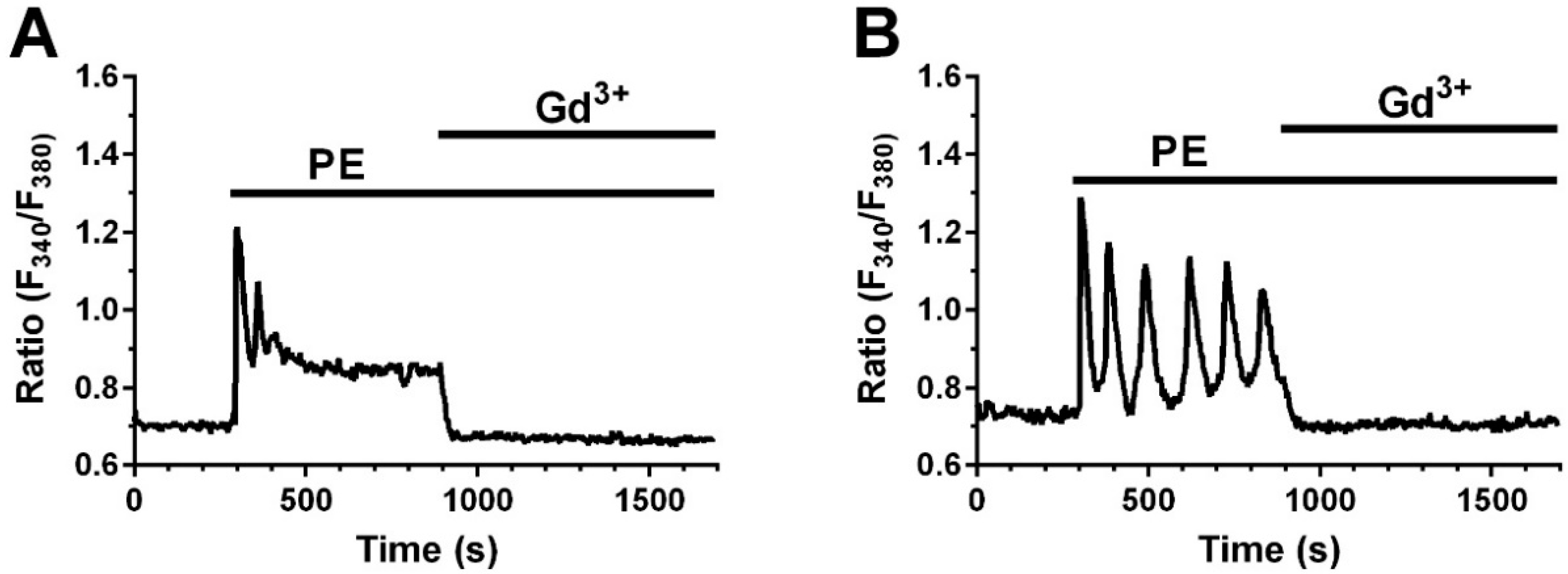
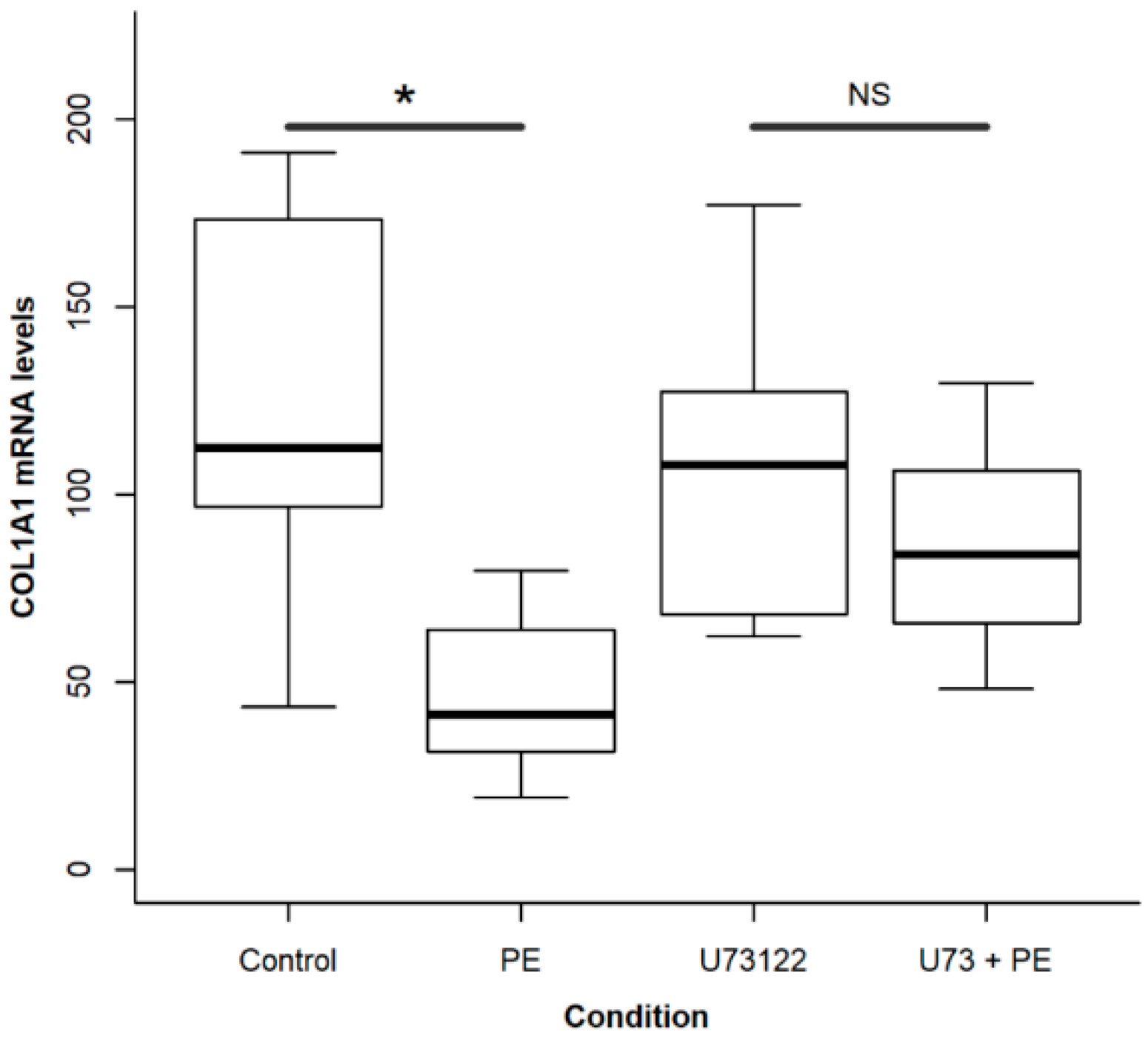
2.4. A Calcium Signal Is Elicited by PE and a Phospholipase C Inhibitor Decreases PE Effect on Collagen Expression and Fibroblast Apoptosis
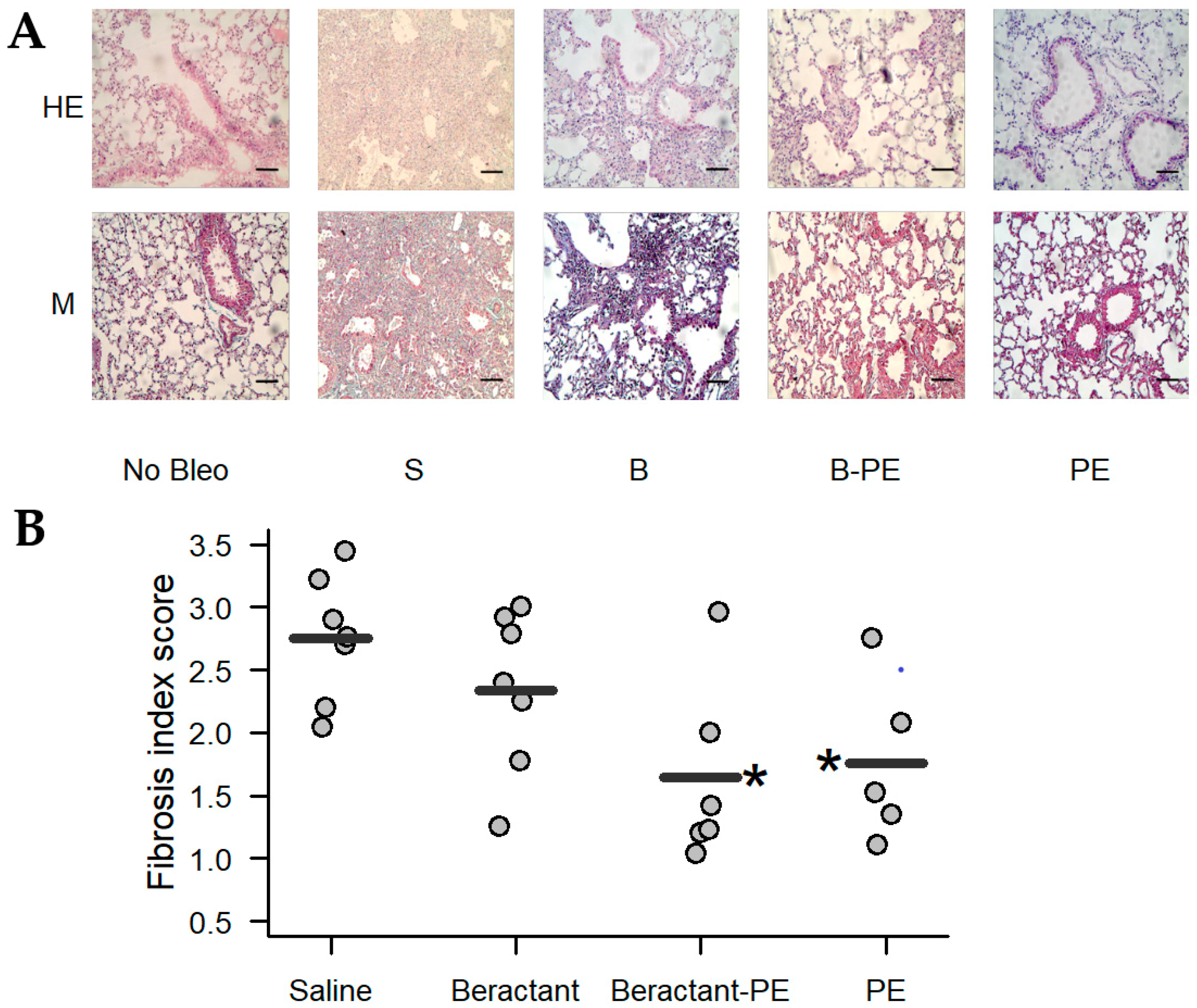
2.5. Phosphatidylethanolamine Mitigates Bleomycin-Induced Lung Fibrosis
3. Discussion
4. Materials and Methods
4.1. Isolation and Purification of Normal Human Lung Fibroblasts
4.2. RNA Isolation and RT-PCR Analysis
4.3. Detection of Apoptosis
4.4. Cytosolic Ca2+ Measurements
4.5. In Vivo Murine Model
4.6. Soluble Collagen Assay
4.7. Histologic Examination
4.8. Preparation of Phospholipids
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 0Ca2+ | Removal of extracellular Ca2+ |
| 2-APB | 2-Aminoethoxydiphenyl borate |
| CPA | Cyclopiazonic acid |
| DMSO | Dimethyl sulfoxide |
| DPPC | Dipalmitoilphosphatidilcoline |
| ER | Endoplasmic reticulum |
| InsP3 | Inositol-1,4,5-trisphosphate |
| IPF | Idiopathic pulmonary fibrosis |
| NHLF | Normal human lung fibroblasts |
| PC | Phosphatidylcholine |
| PE | Phosphatidylethanolamine |
| PG | Phosphatidylglycerol |
| PL | Phospholipids |
| PLC | Phospholipase C |
| RLU | Relative Light Units |
| SERCA | Sarco-endoplasmic reticulum Ca2+-ATPase |
| SOCE | Store-operated Ca2+ entry |
References
- Lederer, D.J.; Martinez, F.J. Idiopathic Pulmonary Fibrosis. N. Engl. J. Med. 2018, 378, 1811–1823. [Google Scholar] [CrossRef] [PubMed]
- King, T.E., Jr.; Pardo, A.; Selman, M. Idiopathic pulmonary fibrosis. Lancet 2011, 378, 1949–1961. [Google Scholar] [CrossRef]
- Harari, S.; Caminati, A. IPF: New insight on pathogenesis and treatment. Allergy 2010, 65, 537–553. [Google Scholar] [CrossRef] [PubMed]
- Strieter, R.M. What differentiates normal lung repair and fibrosis? Inflammation, resolution of repair and fibrosis. Proc. Am. Thorac. Soc. 2008, 5, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Y.Y.; Veldhuizen, R.A.; Neumann, A.W.; Petersen, N.O.; Possmayer, F. Current perspectives in pulmonary surfactant—Inhibition, enhancement and evaluation. Biochim. Biophys. Acta 2008, 1778, 1947–1977. [Google Scholar] [CrossRef] [PubMed]
- Poets, C.F.; Lorenz, L. Prevention of bronchopulmonary dysplasia in extremely low gestational age neonates: Current evidence. Arch. Dis. Child. Fetal Neonatal Ed. 2018, 103, F285–F291. [Google Scholar] [CrossRef] [PubMed]
- Vazquez de Lara, L.G.; Becerril, C.; Montano, M.; Ramos, C.; Maldonado, V.; Melendez, J.; Phelps, D.S.; Pardo, A.; Selman, M. Surfactant components modulate fibroblast apoptosis and type I collagen and collagenase-1 expression. Am. J. Physiol. Lung Cell. Mol. Physiol. 2000, 279, L950–L957. [Google Scholar] [CrossRef] [PubMed]
- Guzman-Silva, A.; Vazquez de Lara, L.G.; Torres-Jacome, J.; Vargaz-Guadarrama, A.; Flores-Flores, M.; Pezzat, S.E.; Lagunas-Martinez, A.; Mendoza-Milla, C.; Tanzi, F.; Moccia, F.; et al. Lung beractant increases free cytosolic levels of Ca2+ in human lung fibroblasts. PLoS ONE 2015, 10, e0134564. [Google Scholar] [CrossRef]
- Darby, I.A.; Hewitson, T.D. Fibroblast differentiation in wound healing and fibrosis. Int. Rev. Cytol. 2007, 257, 143–179. [Google Scholar] [CrossRef] [PubMed]
- Dragoni, S.; Laforenza, U.; Bonetti, E.; Lodola, F.; Bottino, C.; Berra-Romani, R.; Carlo, B.G.; Cinelli, M.P.; Guerra, G.; Pedrazzoli, P.; et al. Vascular endothelial growth factor stimulates endothelial colony forming cells proliferation and tubulogenesis by inducing oscillations in intracellular Ca2+ concentration. Stem Cells 2011, 29, 1898–1907. [Google Scholar] [CrossRef] [PubMed]
- Moccia, F.; Dragoni, S.; Poletto, V.; Rosti, V.; Tanzi, F.; Ganini, C.; Porta, C. Orai1 and transient receptor potential channels as novel molecular targets to impair tumor neovascularization in renal cell carcinoma and other malignancies. Anticancer Agents Med. Chem. 2014, 14, 296–312. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.; Ruppert, C.; Markart, P.; Lubke, N.; Ermert, L.; Weissmann, N.; Breithecker, A.; Ermert, M.; Seeger, W.; Gunther, A. Changes in pulmonary surfactant function and composition in bleomycin-induced pneumonitis and fibrosis. Toxicol. Appl. Pharmacol. 2004, 195, 218–231. [Google Scholar] [CrossRef] [PubMed]
- Akella, A.; Deshpande, S.B. Pulmonary surfactants and their role in pathophysiology of lung disorders. Indian J. Exp. Biol. 2013, 51, 5–22. [Google Scholar] [PubMed]
- Phelps, D.S.; Umstead, T.M.; Mejia, M.; Carrillo, G.; Pardo, A.; Selman, M. Increased surfactant protein-A levels in patients with newly diagnosed idiopathic pulmonary fibrosis. Chest 2004, 125, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Casals, C. Role of surfactant protein A (SP-A)/lipid interactions for SP-A functions in the lung. Pediatr. Pathol. Mol. Med. 2001, 20, 249–268. [Google Scholar] [CrossRef] [PubMed]
- Cutroneo, K.R.; White, S.L.; Phan, S.H.; Ehrlich, H.P. Therapies for bleomycin induced lung fibrosis through regulation of TGF-β1 induced collagen gene expression. J. Cell. Physiol. 2007, 211, 585–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calabresi, C.; Arosio, B.; Galimberti, L.; Scanziani, E.; Bergottini, R.; Annoni, G.; Vergani, C. Natural aging, expression of fibrosis-related genes and collagen deposition in rat lung. Exp. Gerontol. 2007, 42, 1003–1011. [Google Scholar] [CrossRef] [PubMed]
- Boston, M.E.; Frech, G.C.; Chacon-Cruz, E.; Buescher, E.S.; Oelberg, D.G. Surfactant releases internal calcium stores in neutrophils by G protein-activated pathway. Exp. Biol. Med. 2004, 229, 99–107. [Google Scholar] [CrossRef]
- Lewis, R.S. Calcium oscillations in T-cells: Mechanisms and consequences for gene expression. Biochem. Soc. Trans. 2003, 31, 925–929. [Google Scholar] [CrossRef] [PubMed]
- Parekh, A.B. Decoding cytosolic Ca2+ oscillations. Trends Biochem. Sci. 2011, 36, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Gomez, A.M.; Ruiz-Hurtado, G.F.; Benitah, J.P.; Dominguez-Rodriguez, A. Ca2+ fluxes involvement in gene expression during cardiac hypertrophy. Curr. Vasc. Pharmacol. 2013, 11, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Janssen, L.J.; Farkas, L.; Rahman, T.; Kolb, M.R. ATP stimulates Ca2+-waves and gene expression in cultured human pulmonary fibroblasts. Int. J. Biochem. Cell Biol. 2009, 41, 2477–2484. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Duan, F.; Kolb, M.R.; Janssen, L.J. Platelet derived growth factor-evoked Ca2+ wave and matrix gene expression through phospholipase C in human pulmonary fibroblast. Int. J. Biochem. Cell Biol. 2013, 45, 1516–1524. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Sheng, W.; Sun, R.; Janssen, L.J. Ca2+/calmodulin-dependent protein kinase IIβ and IIdelta mediate TGFβ-induced transduction of fibronectin and collagen in human pulmonary fibroblasts. Am. J. Physiol. Lung Cell. Mol. Physiol. 2017, 312, L510–L519. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Ren, Y.; Ma, Y.; Wang, Y.; Jiang, H.; Chaudhari, S.; Davis, M.E.; Zuckerman, J.E.; Ma, R. Negative regulation of Smad1 pathway and collagen IV expression by store-operated Ca2+ entry in glomerular mesangial cells. Am. J. Physiol. Ren. Physiol. 2017, 312, F1090–F1100. [Google Scholar] [CrossRef] [PubMed]
- Klenz, U.; Saleem, M.; Meyer, M.C.; Galla, H.J. Influence of lipid saturation grade and headgroup charge: A refined lung surfactant adsorption model. Biophys. J. 2008, 95, 699–709. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.G. How lipids affect the activities of integral membrane proteins. Biochim. Biophys. Acta 2004, 1666, 62–87. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Li, M.; Chen, T.; Sun, H.; Zhu, J.; Li, X.; Wu, F.; Wang, B.; Li, J.; Chen, Y. PE-induced apoptosis in SMMC7721 cells: Involvement of Erk and Stat signaling pathways. Int. J. Mol. Med. 2014, 34, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Li, J.; Guan, H.; Cai, W.; Bai, X.; Fang, X.; Hu, X.; Wang, Y.; Wang, H.; Zheng, Z.; et al. Anti-fibrotic actions of interleukin-10 against hypertrophic scarring by activation of PI3K/AKT and STAT3 signaling pathways in scar-forming fibroblasts. PLoS ONE 2014, 9, e98228. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Kim, H.J.; Woo, K.J.; Cho, D.; Bang, S.I. Lipo-PGE1 suppresses collagen production in human dermal fibroblasts via the ERK/Ets-1 signaling pathway. PLoS ONE 2017, 12, e0179614. [Google Scholar] [CrossRef] [PubMed]
- Zuccolo, E.; Di, B.C.; Lodola, F.; Orecchioni, S.; Scarpellino, G.; Kheder, D.A.; Poletto, V.; Guerra, G.; Bertolini, F.; Balduini, A.; et al. Stromal cell-derived factor-1α promotes endothelial colony-forming cell migration through the Ca2+-dependent activation of the extracellular signal-regulated kinase 1/2 and phosphoinositide 3-kinase/AKT pathways. Stem Cells Dev. 2018, 27, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Araki, T.; van Egmond, W.N.; van Haastert, P.J.; Williams, J.G. Dual regulation of a Dictyostelium STAT by cGMP and Ca2+ signaling. J. Cell Sci. 2010, 123, 837–841. [Google Scholar] [CrossRef] [PubMed]
- Kolb, M.; Bonella, F.; Wollin, L. Therapeutic targets in idiopathic pulmonary fibrosis. Respir. Med. 2017, 131, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Loney, R.W.; Anyan, W.R.; Biswas, S.C.; Rananavare, S.B.; Hall, S.B. The accelerated late adsorption of pulmonary surfactant. Langmuir 2011, 27, 4857–4866. [Google Scholar] [CrossRef] [PubMed]
- Jordanova, A.; Georgiev, G.A.; Alexandrov, S.; Todorov, R.; Lalchev, Z. Influence of surfactant protein C on the interfacial behavior of phosphatidylethanolamine monolayers. Eur. Biophys. J. 2009, 38, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Kaviratna, A.S.; Banerjee, R. Nanovesicle aerosols as surfactant therapy in lung injury. Nanomedicine 2012, 8, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Yang, R.; Zhong, J.G.; Fang, F.; Jiang, J.J.; Liu, M.Y.; Lu, J. Aerosolised surfactant generated by a novel noninvasive apparatus reduced acute lung injury in rats. Crit. Care 2009, 13, R31. [Google Scholar] [CrossRef] [PubMed]
- Fattman, C.L.; Gambelli, F.; Hoyle, G.; Pitt, B.R.; Ortiz, L.A. Epithelial expression of TIMP-1 does not alter sensitivity to bleomycin-induced lung injury in C57BL/6 mice. Am. J. Physiol. Lung Cell. Mol. Physiol. 2008, 294, L572–L581. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Intratracheal Bleomycin | Treatment | n |
|---|---|---|---|
| 1 | No (normal saline) | Normal saline (1 mL) | 7 |
| 2 | Yes (4 units Kg−1) | Normal saline (1 mL) | 7 |
| 3 | Yes (4 units Kg−1) | Beractant (400 mg·Kg−1) | 7 |
| 4 | Yes (4 units Kg−1) | Beractant-PE (400 mg·Kg−1) * | 6 |
| 5 | Yes (4 units Kg−1) | PE (400 mg·Kg−1) | 5 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vazquez-de-Lara, L.G.; Tlatelpa-Romero, B.; Romero, Y.; Fernández-Tamayo, N.; Vazquez-de-Lara, F.; M. Justo-Janeiro, J.; Garcia-Carrasco, M.; De-la-Rosa Paredes, R.; Cisneros-Lira, J.G.; Mendoza-Milla, C.; et al. Phosphatidylethanolamine Induces an Antifibrotic Phenotype in Normal Human Lung Fibroblasts and Ameliorates Bleomycin-Induced Lung Fibrosis in Mice. Int. J. Mol. Sci. 2018, 19, 2758. https://doi.org/10.3390/ijms19092758
Vazquez-de-Lara LG, Tlatelpa-Romero B, Romero Y, Fernández-Tamayo N, Vazquez-de-Lara F, M. Justo-Janeiro J, Garcia-Carrasco M, De-la-Rosa Paredes R, Cisneros-Lira JG, Mendoza-Milla C, et al. Phosphatidylethanolamine Induces an Antifibrotic Phenotype in Normal Human Lung Fibroblasts and Ameliorates Bleomycin-Induced Lung Fibrosis in Mice. International Journal of Molecular Sciences. 2018; 19(9):2758. https://doi.org/10.3390/ijms19092758
Chicago/Turabian StyleVazquez-de-Lara, Luis G., Beatriz Tlatelpa-Romero, Yair Romero, Nora Fernández-Tamayo, Fernando Vazquez-de-Lara, Jaime M. Justo-Janeiro, Mario Garcia-Carrasco, René De-la-Rosa Paredes, José G. Cisneros-Lira, Criselda Mendoza-Milla, and et al. 2018. "Phosphatidylethanolamine Induces an Antifibrotic Phenotype in Normal Human Lung Fibroblasts and Ameliorates Bleomycin-Induced Lung Fibrosis in Mice" International Journal of Molecular Sciences 19, no. 9: 2758. https://doi.org/10.3390/ijms19092758