Evaluation of the Genetic Diversity and Differentiation of Black Locust (Robinia pseudoacacia L.) Based on Genomic and Expressed Sequence Tag-Simple Sequence Repeats



Abstract
:
1. Introduction
2. Results
2.1. Polymorphism and Determination of SSR Markers
2.2. Characteristics of EST-SSRs and G-SSRs
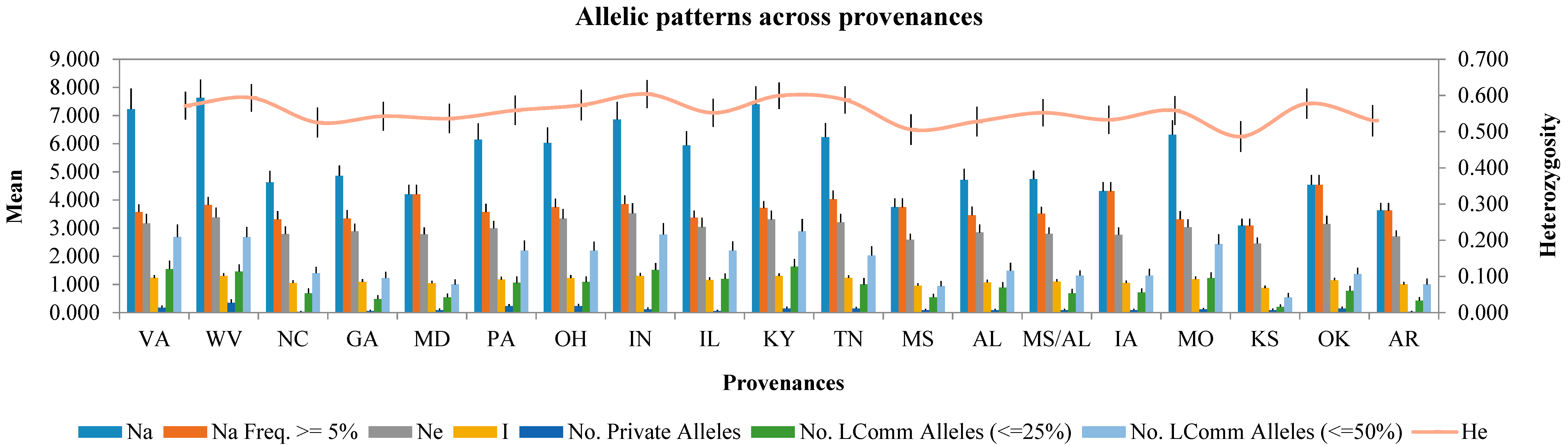
2.3. Genetic Diversity Analysis
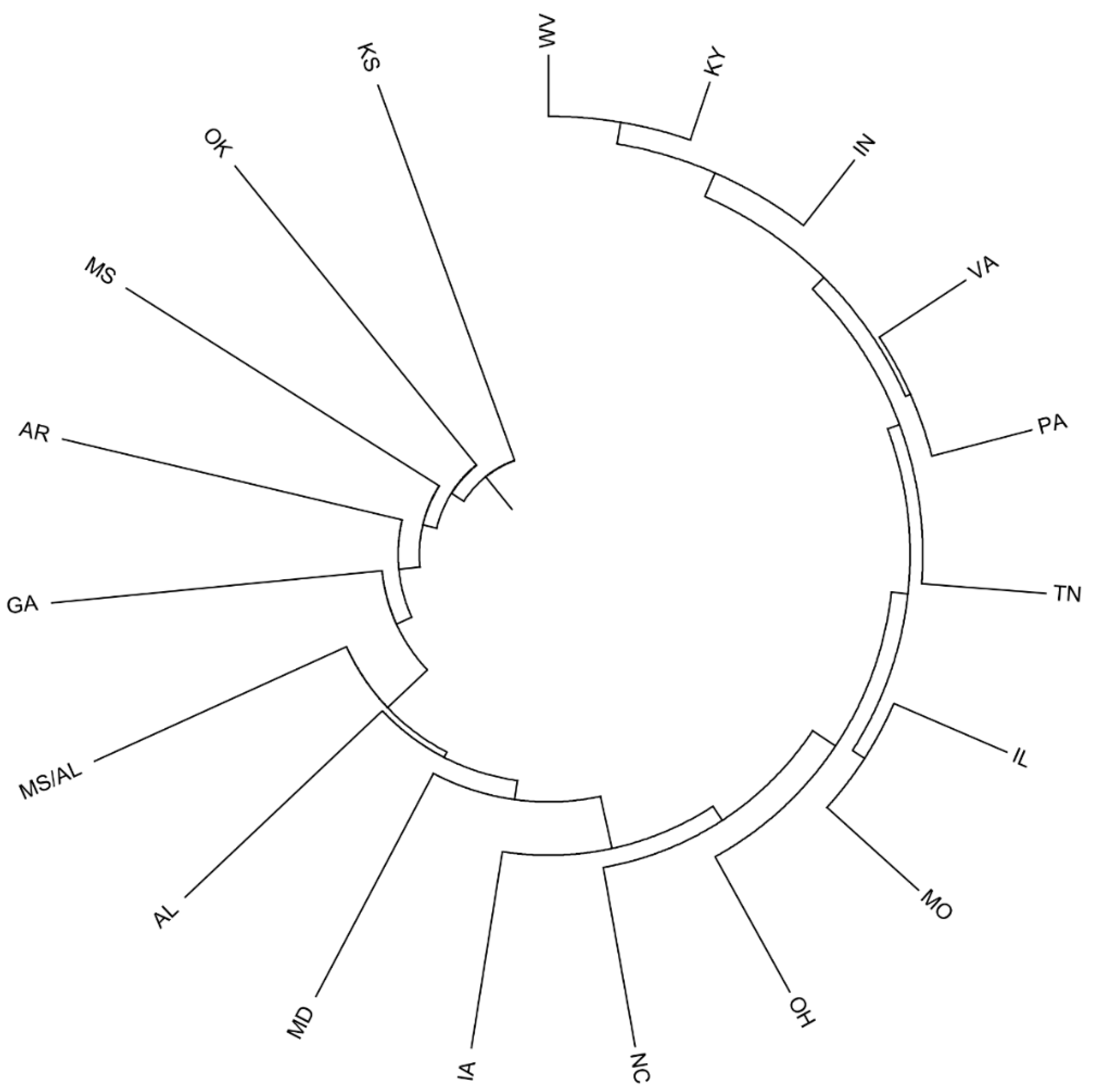
2.4. Provenance Differentiation Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. DNA Extraction
4.3. SSR Marker Screening
4.4. Comparison of EST-SSR and G-SSR outcomes
4.5. SSR Amplification of Provenances
4.6. Genetic Diversity and Differentiation Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Young, A.; Boyle, T.; Brown, T. The population genetic consequences of habitat fragmentation for plants. Trend. Ecol. Evol. 1996, 11, 413–418. [Google Scholar] [CrossRef]
- Dlugosch, K.M.; Parker, I.M. Founding events in species invasions: Genetic variation, adaptive evolution, and the role of multiple introductions. Mol. Ecol. 2008, 17, 431–449. [Google Scholar] [CrossRef] [PubMed]
- Aitken, S.N.; Yeaman, S.; Holliday, J.A.; Wang, T.; Curtis-McLane, S. Adaptation, migration or extirpation: Climate change outcomes for tree populations. Evol. Appl. 2008, 1, 95–111. [Google Scholar] [CrossRef] [PubMed]
- Strayer, D.L.; Eviner, V.T.; Jeschke, J.M.; Pace, M.L. Understanding the long-term effects of species invasions. Trend. Ecol. Evol. 2006, 21, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Machida-Hirano, R. Diversity of potato genetic resources. Breed. Sci. 2015, 65, 26–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vir, R.; Lakhanpaul, S.; Malik, S.; Umdale, S.; Bhat, K.V. Utilization of germplasm for the genetic improvement of mung bean (Vigna radiata (L.) Wilczek): The constraints and the opportunities. In Gene Pool Diversity and Crop Improvement; Rajpal, V., Rao, S., Raina, S., Eds.; Springer International Publishing: Cham, Switzerland, 2016; Volume 10, pp. 367–391. [Google Scholar]
- Kumar, S.; Ambreen, H.; Variath, M.T.; Rao, A.R.; Agarwal, M.; Kumar, A.; Goel, S.; Jagannath, A. Utilization of molecular, phenotypic, and geographical diversity to develop compact composite core collection in the oilseed crop, Safflower (Carthamus tinctorius L.) through maximization strategy. Front. Plant Sci. 2016, 7, 1554. [Google Scholar] [CrossRef] [PubMed]
- Mhlaba, Z.B.; Mashilo, J.; Shimelis, H.; Assefa, A.B.; Modi, A.T. Progress in genetic analysis and breeding of tepary bean (Phaseolus acutifolius A. Gray): A review. Sci. Hortic. 2018, 237, 112–119. [Google Scholar] [CrossRef]
- Cohen, J.I.; Williams, J.T.; Plucknett, D.L.; Shands, H. Ex situ conservation of plant genetic resources: Global development and environmental concerns. Science 1991, 253, 866–872. [Google Scholar] [CrossRef] [PubMed]
- Rao, N.K. Plant genetic resources: Advancing conservation and use through biotechnology. Afr. J. Biotechnol. 2004, 3, 136–145. [Google Scholar] [CrossRef]
- Ulloa, M.; Stewart, J.M.; Garcia-C, E.A.; Godoy-A, S.; Gaytan-M, A.; Acosta, N.S. Cotton genetic resources in the western states of Mexico: In situ conservation status and germplasm collection for ex situ preservation. Genet. Resour. Crop Evol. 2006, 53, 653–668. [Google Scholar] [CrossRef]
- Boring, L.R.; Swank, W.T. The role of black locust (Robinia pseudo-acacia) in forest succession. J. Ecol. 1984, 72, 749–766. [Google Scholar] [CrossRef]
- Surles, S.E.; Hamrick, J.L.; Bongarten, B.C. Allozyme variation in black locust (Robinia pseudoacacia). Can. J. For. Res. 1989, 19, 471–479. [Google Scholar] [CrossRef]
- Rédei, K.; Osvath-Bujtas, Z.; Veperdi, I. Black locust (Robinia pseudoacacia L.) improvement in Hungary: A review. Acta Silvatica et Lignaria Hungarica 2008, 4, 127–132. [Google Scholar]
- Kropf, U.; Korošec, M.; Bertoncelj, J.; Ogrinc, N.; Nečemer, M.; Kump, P.; Golob, T. Determination of the geographical origin of Slovenian black locust, lime and chestnut honey. Food Chem. 2010, 121, 839–846. [Google Scholar] [CrossRef]
- Benesperi, R.; Giuliani, C.; Zanetti, S.; Gennai, M.; Lippi, M.M.; Guidi, T.; Nascimbene, J.; Foggi, B. Forest plant diversity is threatened by Robinia pseudoacacia (black-locust) invasion. Biodivers. Conserv. 2012, 21, 3555–3568. [Google Scholar] [CrossRef]
- Tian, C.; He, X.; Zhong, Y.; Chen, J. Effect of inoculation with ecto-and arbuscular mycorrhizae and Rhizobium on the growth and nitrogen fixation by black locust, Robinia pseudoacacia. New For. 2003, 25, 125–131. [Google Scholar] [CrossRef]
- Qiu, L.P.; Zhang, X.C.; Cheng, J.M.; Yin, X.Q. Effects of black locust (Robinia pseudoacacia) on soil properties in the loessial gully region of the Loess Plateau, China. Plant Soil 2010, 332, 207–217. [Google Scholar] [CrossRef]
- Zhang, J.G.; Guan, J.H.; Shi, W.Y.; Yamanaka, N.; Du, S. Interannual variation in stand transpiration estimated by sap flow measurement in a semi-arid black locust plantation, Loess Plateau, China. Ecohydrology 2015, 8, 137–147. [Google Scholar] [CrossRef]
- Chang, C.S.; Bongarten, B.; Hamrick, J. Genetic structure of natural populations of black locust (Robinia pseudoacacia L.) at Coweeta, North Carolina. J. Plant Res. 1998, 111, 17–24. [Google Scholar] [CrossRef]
- Liesebach, H.; Schneck, V. Genetic diversity and differentiation in a black locust (Robinia pseudoacacia L.) progeny test. Int. J. For. Genet. 2004, 11, 151–161. [Google Scholar]
- Huo, X.M.; Han, H.W.; Zhang, J.; Yang, M.S. Genetic diversity of Robinia pseudoacacia populations in China detected by AFLP markers. Front. Agric. China 2009, 3, 337–345. [Google Scholar] [CrossRef]
- Sun, F.; Yang, M.S.; Zhang, J.; Gu, J.T. ISSR analysis of genetic diversity of Robinia pseudoacacia populations (in Chinese). J. Plant Genet. Resour. 2009, 10, 91–96. [Google Scholar]
- Pejic, I.; Ajmone-Marsan, P.; Morgante, M.; Kozumplick, V.; Castiglioni, P.; Taramino, G.; Motto, M. Comparative analysis of genetic similarity among maize inbred lines detected by RFLPs, RAPDs, SSRs, and AFLPs. Theor. Appl. Genet. 1998, 97, 1248–1255. [Google Scholar] [CrossRef]
- Li, Y.C.; Korol, A.B.; Fahima, T.; Beiles, A.; Nevo, E. Microsatellites: Genomic distribution, putative functions and mutational mechanisms: A review. Mol. Ecol. 2002, 11, 2453–2465. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, A.; Zarei, A.; McKenna, J.R.; Bujdoso, G.; Woeste, K.E. Genetic diversity of Persian walnut (Juglans regia) in the cold-temperate zone of the United States and Europe. Sci. Hortic. 2017, 220, 36–41. [Google Scholar] [CrossRef]
- Lian, C.; Hogetsu, T. Development of microsatellite markers in black locust (Robinia pseudoacacia) using a dual-supression-PCR technique. Mol. Ecol. Resour. 2002, 2, 211–213. [Google Scholar] [CrossRef]
- Lian, C.; Oishi, R.; Miyashita, N.; Hogetsu, T. High somatic instability of a microsatellite locus in a clonal tree, Robinia pseudoacacia. Theor. Appl. Genet. 2004, 108, 836–841. [Google Scholar] [CrossRef] [PubMed]
- Mishima, K.; Hirao, T.; Urano, S.; Watanabe, A.; Takata, K. Isolation and characterization of microsatellite markers from Robinia pseudoacacia L. Mol. Ecol. Resour. 2008, 9, 850–852. [Google Scholar] [CrossRef] [PubMed]
- Liesebach, H.; Ewald, E. Optimisation of a multiplex PCR assay of nuclear microsatellite markers for population genetics and clone identification in Robinia pseudoacacia L. Silvae Genet. 2012, 61, 142–148. [Google Scholar] [CrossRef]
- Malvolti, M.E.; Olimpieri, I.; Pollegioni, P.; Cseke, K.; Keserű, Z.; Rédei, K. Black locust (Robinia pseudoacacia L.) root cuttings: Diversity and identity revealed by SSR genotyping: A case study. South-East Eur. For. 2015, 6, 201–217. [Google Scholar] [CrossRef]
- Bretting, P.K.; Duvick, D.N. Dynamic conservation of plant genetic resources. Adv. Agron. 1997, 61, 51. [Google Scholar]
- Liu, S.; Liu, H.; Wu, A.; Hou, Y.; An, Y.; Wei, C. Construction of fingerprinting for tea plant (Camellia sinensis) accessions using new genomic SSR markers. Mol. Breed. 2017, 37, 93. [Google Scholar] [CrossRef]
- Sathyanarayana, N.; Pittala, R.K.; Tripathi, P.K.; Chopra, R.; Singh, H.R.; Belamkar, V.; Bhardwaj, P.K.; Doyle, J.J.; Egan, A.N. Transcriptomic resources for the medicinal legume Mucuna pruriens: De novo transcriptome assembly, annotation, identification and validation of EST-SSR markers. BMC Genom. 2017, 18, 409. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.Y.; Zhou, C.P.; Zhang, Y.; Zhang, W.Q.; Gan, X.H.; Zhang, H.X.; Guo, Y.; Gan, S.M. A novel set of 223 EST-SSR markers in Casuarina L. ex Adans.: Polymorphisms, cross-species transferability, and utility for commercial clone genotyping. Tree Genet. Genomes 2018, 14, 30. [Google Scholar] [CrossRef]
- Albanèse, V.; Biguet, N.F.; Kiefer, H.; Bayard, E.; Mallet, J.; Meloni, R. Quantitative effects on gene silencing by allelic variation at a tetranucleotide microsatellite. Hum. Mol. Genet. 2001, 10, 1785–1792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nandha, P.S.; Singh, J. Comparative assessment of genetic diversity between wild and cultivated barley using gSSR and EST-SSR markers. Plant. Breed. 2014, 133, 28–35. [Google Scholar] [CrossRef]
- Choudhary, S.B.; Sharma, H.K.; Kumar, A.A.; Chowdhury, I.; Maruthi, R.T.; Kak, A. Genetic diversity spectrum and marker trait association for agronomic traits in global accessions of Linum usitatissimum L. Ind. Crop. Prod. 2017, 108, 604–615. [Google Scholar] [CrossRef]
- Wen, M.F.; Wang, H.Y.; Xia, Z.Q.; Zou, M.L.; Lu, C.; Wang, W.Q. Development of EST-SSR and genomic-SSR markers to assess genetic diversity in Jatropha Curcas L. BMC Res. Notes 2010, 3, 42. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.Z.; Wu, F.; Luo, K.; Zhao, Y.F.; Yan, Q.; Zhang, Y.F.; Wang, Y.R.; Zhang, J.Y. Cross-species transferability of EST-SSR markers developed from the transcriptome of Melilotus and their application to population genetics research. Sci. Rep. 2017, 7, 17959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selkoe, K.A.; Toonen, R.J. Microsatellites for ecologists: A practical guide to using and evaluating microsatellite markers. Ecol. Lett. 2006, 9, 615–629. [Google Scholar] [CrossRef] [PubMed]
- Lazrek, F.; Roussel, V.; Ronfort, J.; Cardinet, G.; Chardon, F.; Aouani, M.E.; Huguet, T. The use of neutral and non-neutral SSRs to analyse the genetic structure of a Tunisian collection of Medicago truncatula lines and to reveal associations with eco-environmental variables. Genetica 2009, 135, 391. [Google Scholar] [CrossRef] [PubMed]
- Bruford, M.W.; Ciofi, C.; Funk, S.M. Characteristics of microsatellites. In Molecular Tools for Screening Biodiversity; Springer: Dordrecht, The Netherlands, 1998; pp. 202–205. [Google Scholar]
- Pandian, S.; Satish, L.; Rameshkumar, R.; Muthuramalingam, P.; Rency, A.S.; Rathinapriya, P.; Ramesh, M. Analysis of population structure and genetic diversity in an exotic germplasm collection of Eleusine coracana (L.) Gaertn. using genic-SSR markers. Gene 2018, 653, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Slatkin, M.; Barton, N.H. A comparison of three indirect methods for estimating average levels of gene flow. Evolution 1989, 43, 1349–1368. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.S.; Black, R. The Wahlund effect and the geographical scale of variation in the intertidal limpet Siphonaria sp. Mar. Biol. 1984, 79, 295–302. [Google Scholar] [CrossRef]
- Zhao, K.Q. Development and Application of EST-SSR Primers in Robinia pseudoacacia L. Master’s Dissertation, Beijing Forestry University, Beijing, China, 2014. [Google Scholar]
- Guo, Q.; Wang, J.X.; Su, L.Z.; Lv, W.; Sun, Y.H.; Li, Y. Development and evaluation of a novel set of EST-SSR markers based on transcriptome sequences of black locust (Robinia pseudoacacia L.). Genes 2017, 8, 177. [Google Scholar] [CrossRef] [PubMed]
- Schuelke, M. An economic method for the fluorescent labeling of PCR fragments. Nat. Biotechnol. 2000, 18, 233–234. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.O.D.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic sofware for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic sofware for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Goudet, J. FSTAT, a Program to Estimate and Test Gene Diversity and Fixation Indices (Version 2.9.3). Lausanne University. Available online: http://www2.unil.ch/popgen/softwares/fstat.htm (accessed on 23 August 2018).
- Liu, K.J.; Muse, S.V. PowerMarker: An integrated analysis environment for genetic marker analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Lischer, H.E. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.W.; Tompson, E.A. Performing the exact test of Hardy-Weinberg proportion for multiple alleles. Biometrics 1992, 48, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Ewens, W.J. The sampling theory of selectively neutral haplotypes. Theor. Pop. Biol. 1972, 3, 87–112. [Google Scholar] [CrossRef]
- Oosterhout, C.V.; Hutchinson, W.F.; Wills, D.P.M.; Shipley, P. MICRO CHECKER: Sofware for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Wilberg, M.J.; Dreher, B.P. Genecap: A program for analysis of multi-locus genotype data for non-invasive sampling and capture-recapture population estimation. Mol. Ecol. Notes 2004, 4, 783–785. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Repeat Type | EST-SSR Polymorphic Number | Remaining Number after Screening | Percentage | G-SSR Polymorphic Number | Remaining Number after Screening | Percentage |
|---|---|---|---|---|---|---|
| Di- | 4 | 2 | 7.143 | 11 | 7 | 100 |
| Tri- | 14 | 10 | 35.714 | - | 0 | 0 |
| Tetra- | 3 | 3 | 10.714 | - | 0 | 0 |
| Penta- | 7 | 5 | 17.857 | - | 0 | 0 |
| Hexa- | 9 | 8 | 28.571 | - | 0 | 0 |
| Total | 37 | 28 | 100 | 11 | 7 | 100 |
| SSR Marker Type | Genotype Number | Allele Number (N) | Gene Diversity (H) | Polymorphism Information Content (PIC) |
|---|---|---|---|---|
| EST-SSR | 24.286 | 10.464 | 0.535 | 0.503 |
| G-SSR | 65.429 | 20.857 | 0.835 | 0.817 |
| Species | EST-SSR Number of Loci | Percentage | G-SSR Noumber of Loci | Percentage |
|---|---|---|---|---|
| Gleditsia sinensis | 20 | 57.143 | 8 | 61.538 |
| Cercis chinensis | 20 | 57.143 | 6 | 46.154 |
| Wisteria sinensis (Sims) Sweet | 21 | 60.000 | 8 | 61.538 |
| Trifolium repens | 17 | 48.571 | 0 | 0.000 |
| Amorpha fruticosa | 19 | 54.286 | 8 | 61.538 |
| Sophora japonica | 21 | 60.000 | 9 | 69.231 |
| Sophora japonica var. pendula | 21 | 60.000 | 7 | 53.846 |
| Robinia pseudoacacia ‘Frisia’ | 34 | 97.143 | 7 | 53.846 |
| Robinia pseu-doacacia var. decaisneana | 32 | 91.429 | 9 | 69.231 |
| Mean | 22.778 | 65.079 | 6.889 | 52.99 |
| Source of Variation | Degree of Freedom | Sum of Squares | Mean Square | Percentage Variation | Percentage Variation (%) | p-Value |
|---|---|---|---|---|---|---|
| Among Provenances | 18 | 686.448 | 38.136 | 0.715 | 3% | <0.01 |
| Within Provenances | 348 | 8565.051 | 24.612 | 24.612 | 97% | <0.01 |
| Total | 366 | 9251.499 | 25.327 | 100% |
| Provenance Name | Source Sites | Sample Number | Latitude (N) | Longitude (W) | |
|---|---|---|---|---|---|
| VA | Black Burg | 12 | 37°26′ | 80°45′ | |
| Independence | 12 | 36°37′ | 81°07′ | ||
| Washington DC | 14 | 38°27′ | 76°28′ | ||
| WV | New River Gorge | 14 | 38°04′ | 81°03′ | |
| Morgantown | 11 | 39°36′ | 79°57′ | ||
| West Huntington | 14 | 38°23′ | 82°29′ | ||
| NC | Huntersville | 12 | 39°08′ | 90°45′ | |
| GA | Blue Ridge Lake | 13 | 34°51′ | 84°19′ | |
| MD | Old National Pike | 9 | 39°42′ | 78°18′ | |
| PA | Big Beaver Blvd | 13 | 40°32′ | 80°18′ | |
| Bedford | 15 | 40°08′ | 78°30′ | ||
| OH | Cadiz piedmont | 12 | 40°07′ | 81°13′ | |
| Cincinnati | 10 | 39°03′ | 84°31′ | ||
| IN | Fisher | 12 | 39°56′ | 85°53′ | |
| Georgetwon | 13 | 38°17′ | 85°55′ | ||
| Elberfeld | 8 | 38°10′ | 87°26′ | ||
| IL | Bloomington | 12 | 40°28′ | 89°01′ | |
| Toe Exit | 12 | 39°03′ | 88°40′ | ||
| KY | Mt Sterling | 11 | 38°03′ | 84°02′ | |
| Bowling Green | 14 | 37°00′ | 86°17′ | ||
| Kentucky Lake | 6 | 36°59′′ | 88°08′ | ||
| Wickliffe | 10 | 37°01′ | 89°03′ | ||
| TN | Knoxville | 13 | 35°52′ | 83°57′ | |
| Waverly | 14 | 35°53′ | 87°39′ | ||
| MS | Sardis | 8 | 34°26′ | 89°53′ | |
| AL | Upper Elkton RD | 11 | 34°56′ | 86°53′ | |
| MS/AL | Ms/AL Border | 13 | 34°11′ | 88°06′ | |
| IA | Brulington | 10 | 40°50′ | 91°08′ | |
| MO | Hannidal | 8 | 39°43′ | 91°21′ | |
| ST James | 14 | 37°56′ | 91°08′ | ||
| KS | Riverton | 4 | 37°06′ | 94°42′ | |
| OK | Pryor | 8 | 36°19′ | 95°18′ | |
| AR | Colt | 5 | 35°21′ | 80°51′ | |
| Total Number | 19 | 33 | 367 | - | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Q.; Li, X.; Yang, S.; Yang, Z.; Sun, Y.; Zhang, J.; Cao, S.; Dong, L.; Uddin, S.; Li, Y. Evaluation of the Genetic Diversity and Differentiation of Black Locust (Robinia pseudoacacia L.) Based on Genomic and Expressed Sequence Tag-Simple Sequence Repeats. Int. J. Mol. Sci. 2018, 19, 2492. https://doi.org/10.3390/ijms19092492
Guo Q, Li X, Yang S, Yang Z, Sun Y, Zhang J, Cao S, Dong L, Uddin S, Li Y. Evaluation of the Genetic Diversity and Differentiation of Black Locust (Robinia pseudoacacia L.) Based on Genomic and Expressed Sequence Tag-Simple Sequence Repeats. International Journal of Molecular Sciences. 2018; 19(9):2492. https://doi.org/10.3390/ijms19092492
Chicago/Turabian StyleGuo, Qi, Xiuyu Li, Shuhong Yang, Zhiheng Yang, Yuhan Sun, Jiangtao Zhang, Sen Cao, Li Dong, Saleem Uddin, and Yun Li. 2018. "Evaluation of the Genetic Diversity and Differentiation of Black Locust (Robinia pseudoacacia L.) Based on Genomic and Expressed Sequence Tag-Simple Sequence Repeats" International Journal of Molecular Sciences 19, no. 9: 2492. https://doi.org/10.3390/ijms19092492