Intrathecal Injection of Dual Zipper Kinase shRNA Alleviating the Neuropathic Pain in a Chronic Constrictive Nerve Injury Model


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
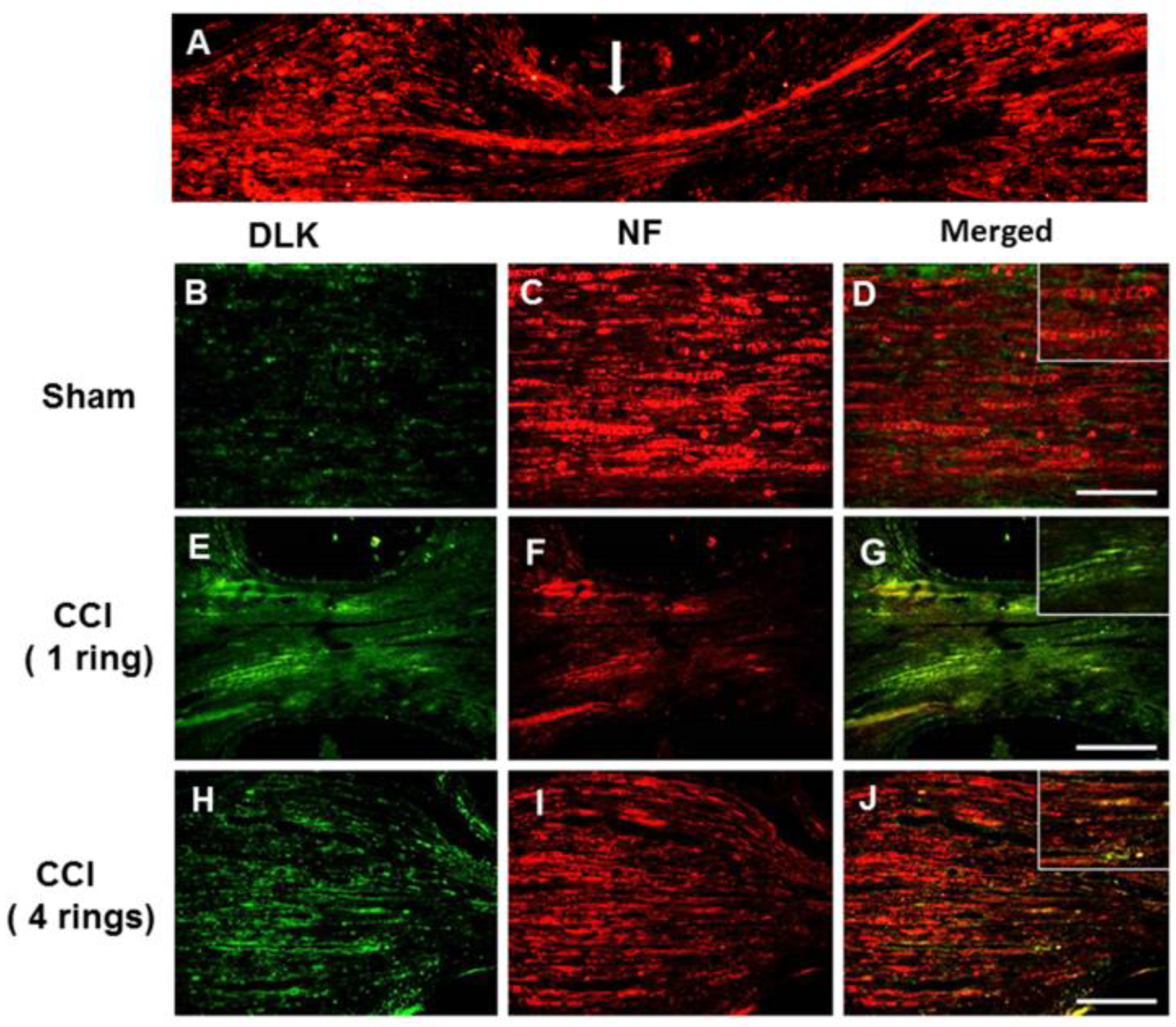
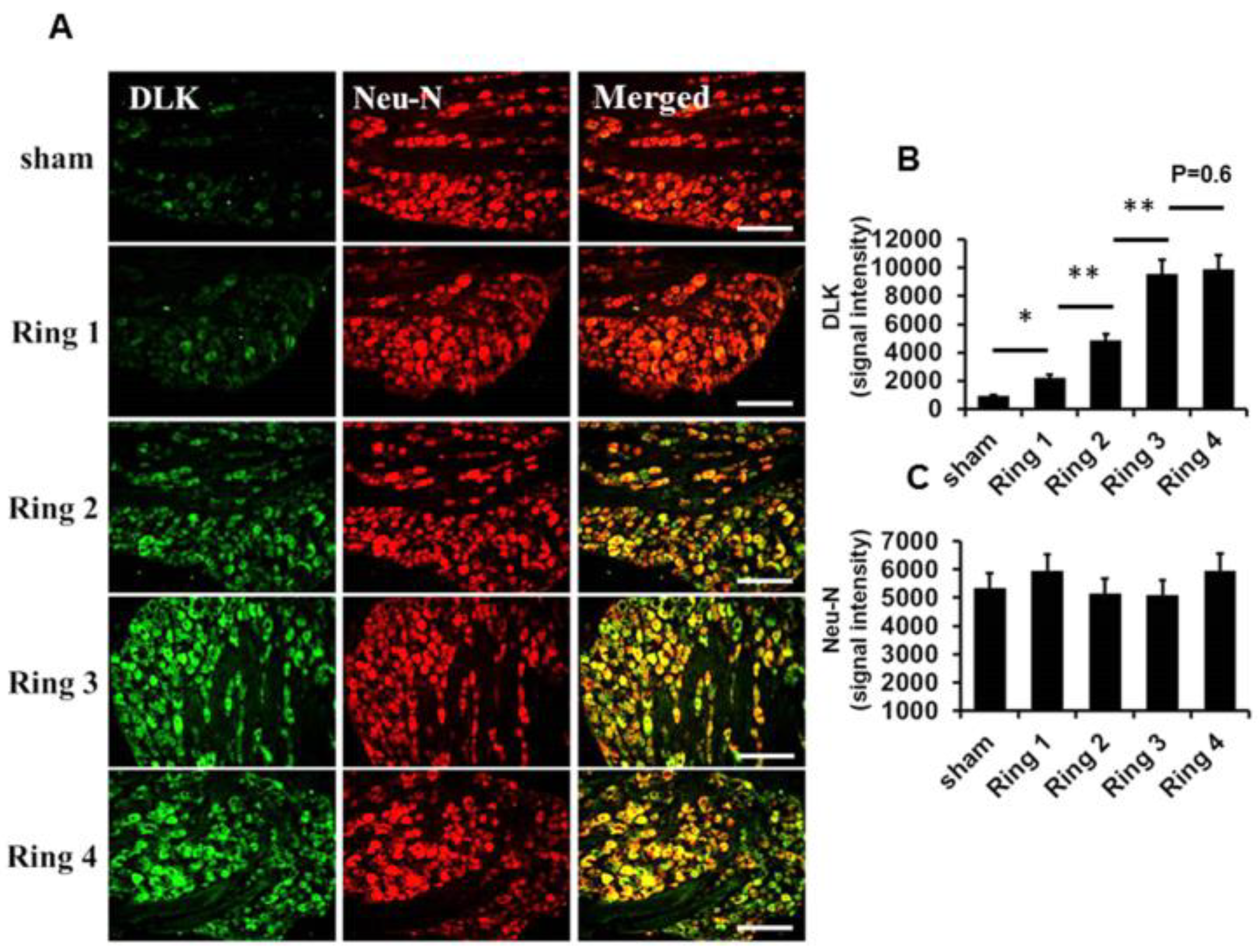
2.1. Escalated Expression of DLK Expression in Dorsal Root Ganglion Cells Subjected to Different Intensities of Nerve Injury
2.2. Construction of DLK shRNA for the Assessment of Inhibition Effects
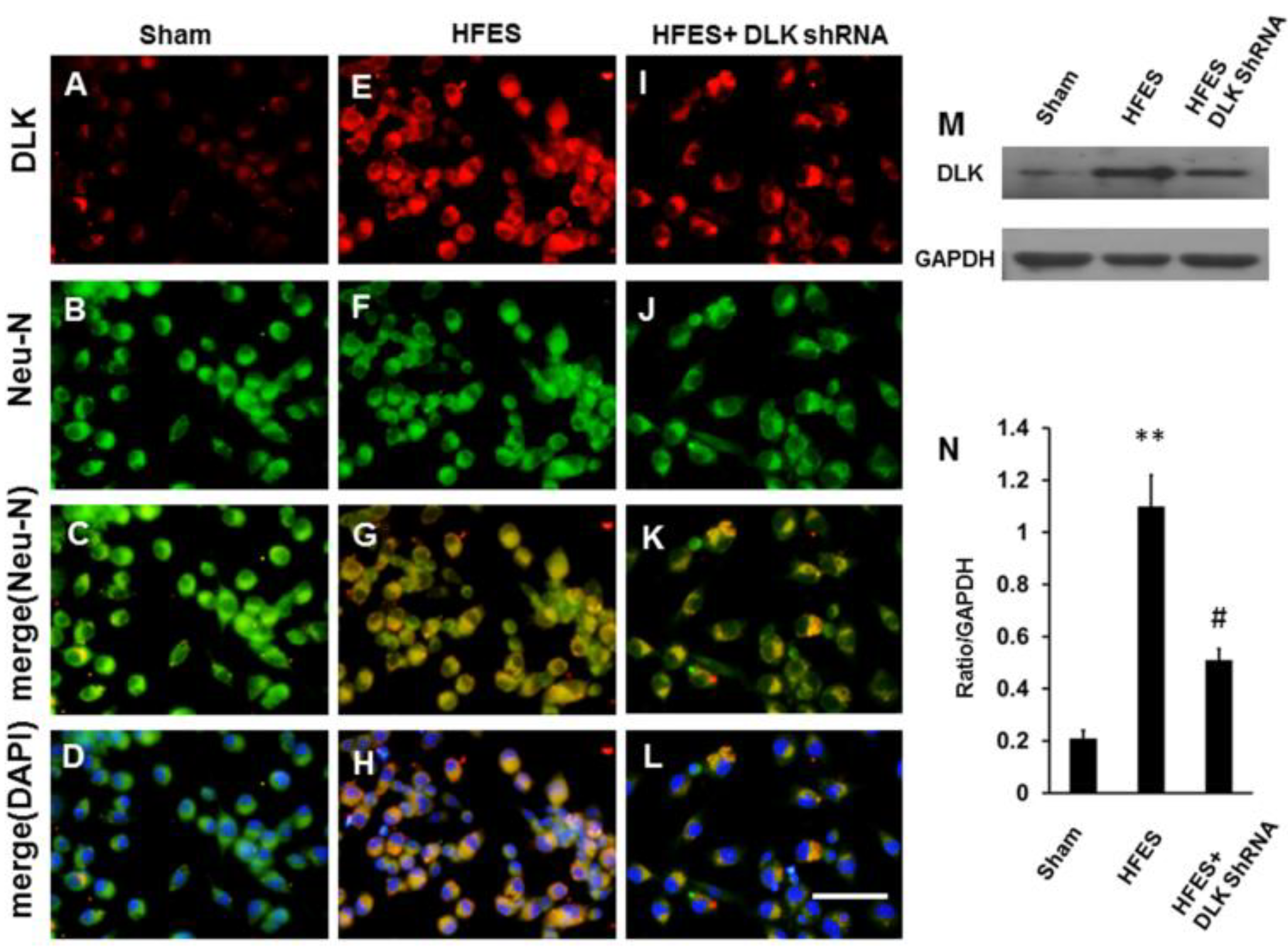
2.3. Expression of DLK in Primary Dorsal Root Ganglion Cells Culture Subjected to High-Frequency Electrical Stimulation Abolished by DLK shRNA
2.4. Improvement of Neurobehavior of CCI by the Intrathecal Injection of DLK shRNA
2.5. Attenuation of DLK over the Dorsal Root Ganglion and Hippocampus Subjected to Intrathecal Injection of shRNA of DLK
3. Discussion
4. Materials and Methods
4.1. Assessment of High Frequency Electrically Stimulated Dorsal Root Ganglion Primary Culture Cells Treated by DLK shRNA
4.2. Construction of DLK shRNA
| shRNA1: AAGTTGGCAGCACCAACACTGATGAGCGA; |
| shRNA2: AAGGAGGTGTCCTGGTCTACTGAAGTCAC; |
| shRNA3: CCTGTCTGGACAATGATTGGCAAAGCCTA; |
| shRNA4: CAGTAGCCTGGATGGCTCCTGAAGTGATC |
4.3. Animal Model of CCI
4.4. Mechanical Allodynia and Thermal Hyperalgesia
4.5. CatWalk Automated Quantitative Gait Analysis
4.6. Somatosensory Evoked Potential
4.7. Western Blot Analysis
4.8. Immunohistochemical ANALYSIS
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DLK | Dual leucine zipper kinase |
| shRNA | Short hairpin RNA |
| MAP3K | Mitogen-activated protein kinase kinase kinase |
| MAPK | Mitogen-activated protein kinase |
| ERK | Extracellular signal-regulated kinase |
| JNK | c-Jun N-terminal kinase |
| DRG | Dorsal root ganglion |
References
- Baron, R.; Tolle, T.R. Assessment and diagnosis of neuropathic pain. Curr. Opin. Support. Palliat. Care 2008, 2, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Treede, R.D.; Jensen, T.S.; Campbell, J.N.; Cruccu, G.; Dostrovsky, J.O.; Griffin, J.W.; Hansson, P.; Hughes, R.; Nurmikko, T.; Serra, J. Neuropathic pain: Redefinition and a grading system for clinical and research purposes. Neurology 2008, 70, 1630–1635. [Google Scholar] [CrossRef] [PubMed]
- Hains, B.C.; Waxman, S.G. Activated microglia contribute to the maintenance of chronic pain after spinal cord injury. J. Neurosci. 2006, 26, 4308–4317. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.X.; Zhuang, Z.Y.; Woolf, C.J.; Ji, R.R. p38 mitogen-activated protein kinase is activated after a spinal nerve ligation in spinal cord microglia and dorsal root ganglion neurons and contributes to the generation of neuropathic pain. J. Neurosci. 2003, 23, 4017–4022. [Google Scholar] [CrossRef]
- Tsuda, M.; Mizokoshi, A.; Shigemoto-Mogami, Y.; Koizumi, S.; Inoue, K. Activation of p38 mitogen-activated protein kinase in spinal hyperactive microglia contributes to pain hypersensitivity following peripheral nerve injury. Glia 2004, 45, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Z.Y.; Gerner, P.; Woolf, C.J.; Ji, R.R. ERK is sequentially activated in neurons, microglia, and astrocytes by spinal nerve ligation and contributes to mechanical allodynia in this neuropathic pain model. Pain 2005, 114, 149–159. [Google Scholar] [CrossRef] [PubMed]
- DeLeo, J.A.; Tanga, F.Y.; Tawfik, V.L. Neuroimmune activation and neuroinflammation in chronic pain and opioid tolerance/hyperalgesia. Neurosci. Rev. J. Bringing Neurobiol. Neurol. Psychiatry 2004, 10, 40–52. [Google Scholar] [CrossRef] [PubMed]
- Ji, R.R.; Strichartz, G. Cell signaling and the genesis of neuropathic pain. Sci. STKE Signal Transduct. Knowl. Environ. 2004, 2004, reE14. [Google Scholar] [CrossRef] [PubMed]
- Watkins, L.R.; Milligan, E.D.; Maier, S.F. Glial activation: A driving force for pathological pain. Trends Neurosci. 2001, 24, 450–455. [Google Scholar] [CrossRef]
- Hirai, S.; Izawa, M.; Osada, S.; Spyrou, G.; Ohno, S. Activation of the JNK pathway by distantly related protein kinases, MEKK and MUK. Oncogene 1996, 12, 641–650. [Google Scholar] [PubMed]
- Holzman, L.B.; Merritt, S.E.; Fan, G. Identification, molecular cloning, and characterization of dual leucine zipper bearing kinase. A novel serine/threonine protein kinase that defines a second subfamily of mixed lineage kinases. J. Biol. Chem. 1994, 269, 30808–30817. [Google Scholar] [PubMed]
- Sakuma, H.; Ikeda, A.; Oka, S.; Kozutsumi, Y.; Zanetta, J.P.; Kawasaki, T. Molecular cloning and functional expression of a cDNA encoding a new member of mixed lineage protein kinase from human brain. J. Biol. Chem. 1997, 272, 28622–28629. [Google Scholar] [CrossRef] [PubMed]
- Hirai, S.; Cui, D.F.; Miyata, T.; Ogawa, M.; Kiyonari, H.; Suda, Y.; Aizawa, S.; Banba, Y.; Ohno, S. The c-Jun N-terminal kinase activator dual leucine zipper kinase regulates axon growth and neuronal migration in the developing cerebral cortex. J. Neurosci. 2006, 26, 11992–12002. [Google Scholar] [CrossRef] [PubMed]
- Hirai, S.; Kawaguchi, A.; Suenaga, J.; Ono, M.; Cui, D.F.; Ohno, S. Expression of MUK/DLK/ZPK, an activator of the JNK pathway, in the nervous systems of the developing mouse embryo. Gene Expr. Patterns GEP 2005, 5, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Mata, M.; Merritt, S.E.; Fan, G.; Yu, G.G.; Holzman, L.B. Characterization of dual leucine zipper-bearing kinase, a mixed lineage kinase present in synaptic terminals whose phosphorylation state is regulated by membrane depolarization via calcineurin. J. Biol. Chem. 1996, 271, 16888–16896. [Google Scholar] [CrossRef] [PubMed]
- Saxena, S.; Caroni, P. Mechanisms of axon degeneration: From development to disease. Prog. Neurobiol. 2007, 83, 174–191. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.R.; Press, C.; Daniels, R.W.; Sasaki, Y.; Milbrandt, J.; DiAntonio, A. A dual leucine kinase-dependent axon self-destruction program promotes Wallerian degeneration. Nat. Neurosci. 2009, 12, 387–389. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.S.; Wu, Y.; Culver, D.G.; Glass, J.D. Pathogenesis of axonal degeneration: Parallels between Wallerian degeneration and vincristine neuropathy. J. Neuropathol. Exp. Neurol. 2000, 59, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Kong, J.; Xu, Z. Massive mitochondrial degeneration in motor neurons triggers the onset of amyotrophic lateral sclerosis in mice expressing a mutant SOD1. J. Neurosci. 1998, 18, 3241–3250. [Google Scholar] [CrossRef]
- Tedeschi, A.; Bradke, F. The DLK signalling pathway—A double-edged sword in neural development and regeneration. EMBO Rep. 2013, 14, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, K.; Sloan, S.A.; Bennett, M.L.; Scholze, A.R.; O’Keeffe, S.; Phatnani, H.P.; Guarnieri, P.; Caneda, C.; Ruderisch, N.; et al. An RNA-sequencing transcriptome and splicing database of glia, neurons, and vascular cells of the cerebral cortex. J. Neurosci. 2014, 34, 11929–11947. [Google Scholar] [CrossRef] [PubMed]
- Le Pichon, C.E.; Meilandt, W.J.; Dominguez, S.; Solanoy, H.; Lin, H.; Ngu, H.; Gogineni, A.; Sengupta Ghosh, A.; Jiang, Z.; Lee, S.H.; et al. Loss of dual leucine zipper kinase signaling is protective in animal models of neurodegenerative disease. Sci. Transl. Med. 2017, 9, eaag0394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watkins, T.A.; Wang, B.; Huntwork-Rodriguez, S.; Yang, J.; Jiang, Z.; Eastham-Anderson, J.; Modrusan, Z.; Kaminker, J.S.; Tessier-Lavigne, M.; Lewcock, J.W. DLK initiates a transcriptional program that couples apoptotic and regenerative responses to axonal injury. Proc. Natl. Acad. Sci. USA 2013, 110, 4039–4044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, C.Y.; Sheu, M.L.; Cheng, F.C.; Chen, C.J.; Su, H.L.; Sheehan, J.; Pan, H.C. Comprehensive analysis of neurobehavior associated with histomorphological alterations in a chronic constrictive nerve injury model through use of the CatWalk XT system. J. Neurosurg. 2014, 120, 250–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, H.L.; Chiang, C.Y.; Lu, Z.H.; Cheng, F.C.; Chen, C.J.; Sheu, M.L.; Sheehan, J.; Pan, H.C. Late administration of high-frequency electrical stimulation increases nerve regeneration without aggravating neuropathic pain in a nerve crush injury. BMC Neurosci. 2018, 19, 37. [Google Scholar] [CrossRef] [PubMed]
- Hirai, S.; Banba, Y.; Satake, T.; Ohno, S. Axon formation in neocortical neurons depends on stage-specific regulation of microtubule stability by the dual leucine zipper kinase-c-Jun N-terminal kinase pathway. J. Neurosci. 2011, 31, 6468–6480. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.S.; Wang, B.; Pozniak, C.D.; Chen, M.; Watts, R.J.; Lewcock, J.W. DLK induces developmental neuronal degeneration via selective regulation of proapoptotic JNK activity. J. Cell Biol. 2011, 194, 751–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itoh, A.; Horiuchi, M.; Wakayama, K.; Xu, J.; Bannerman, P.; Pleasure, D.; Itoh, T. ZPK/DLK, a mitogen-activated protein kinase kinase kinase, is a critical mediator of programmed cell death of motoneurons. J. Neurosci. 2011, 31, 7223–7228. [Google Scholar] [CrossRef] [PubMed]
- Bennett, G.J.; Xie, Y.K. A peripheral mononeuropathy in rat that produces disorders of pain sensation like those seen in man. Pain 1988, 33, 87–107. [Google Scholar] [CrossRef]
- Chiang, C.Y.; Liu, S.A.; Sheu, M.L.; Chen, F.C.; Chen, C.J.; Su, H.L.; Pan, H.C. Feasibility of Human Amniotic Fluid Derived Stem Cells in Alleviation of Neuropathic Pain in Chronic Constrictive Injury Nerve Model. PLoS ONE 2016, 11, e0159482. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.Y.; Liu, M.G.; Yuan, D.L.; Wang, Y.; He, Y.; Wang, D.D.; Chen, X.F.; Zhang, F.K.; Li, H.; He, X.S.; et al. Nociception-induced spatial and temporal plasticity of synaptic connection and function in the hippocampal formation of rats: A multi-electrode array recording. Mol. Pain 2009, 5, 55. [Google Scholar] [PubMed]
- Liu, M.G.; Chen, J. Roles of the hippocampal formation in pain information processing. Neurosci. Bull. 2009, 25, 237–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wlaschin, J.J.; Gluski, J.M.; Nguyen, E.; Silberberg, H.; Thompson, J.H.; Chesler, A.T.; Le Pichon, C.E. Dual leucine zipper kinase is required for mechanical allodynia and microgliosis after nerve injury. eLife 2018, 7, e33910. [Google Scholar] [CrossRef] [PubMed]
- Hirai, T.; Enomoto, M.; Kaburagi, H.; Sotome, S.; Yoshida-Tanaka, K.; Ukegawa, M.; Kuwahara, H.; Yamamoto, M.; Tajiri, M.; Miyata, H.; et al. Intrathecal AAV serotype 9-mediated delivery of shRNA against TRPV1 attenuates thermal hyperalgesia in a mouse model of peripheral nerve injury. Mol. Ther. 2014, 22, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Cheng, F.C.; Sheu, M.L.; Su, H.L.; Chen, C.J.; Sheehan, J.; Pan, H.C. Detection of subtle neurological alterations by the Catwalk XT gait analysis system. J. Neuroeng. Rehabil. 2014, 11, 62. [Google Scholar] [CrossRef] [PubMed]








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sheu, M.-L.; Chiang, C.-Y.; Su, H.-L.; Chen, C.-J.; Sheehan, J.; Pan, H.-C. Intrathecal Injection of Dual Zipper Kinase shRNA Alleviating the Neuropathic Pain in a Chronic Constrictive Nerve Injury Model. Int. J. Mol. Sci. 2018, 19, 2421. https://doi.org/10.3390/ijms19082421
Sheu M-L, Chiang C-Y, Su H-L, Chen C-J, Sheehan J, Pan H-C. Intrathecal Injection of Dual Zipper Kinase shRNA Alleviating the Neuropathic Pain in a Chronic Constrictive Nerve Injury Model. International Journal of Molecular Sciences. 2018; 19(8):2421. https://doi.org/10.3390/ijms19082421
Chicago/Turabian StyleSheu, Meei-Ling, Chien-Yi Chiang, Hong-Lin Su, Chun-Jung Chen, Jason Sheehan, and Hung-Chuan Pan. 2018. "Intrathecal Injection of Dual Zipper Kinase shRNA Alleviating the Neuropathic Pain in a Chronic Constrictive Nerve Injury Model" International Journal of Molecular Sciences 19, no. 8: 2421. https://doi.org/10.3390/ijms19082421