Identification and Characterization of miRNAs in Self-Rooted and Grafted Malus Reveals Critical Networks Associated with Flowering

Abstract
:1. Introduction
2. Results
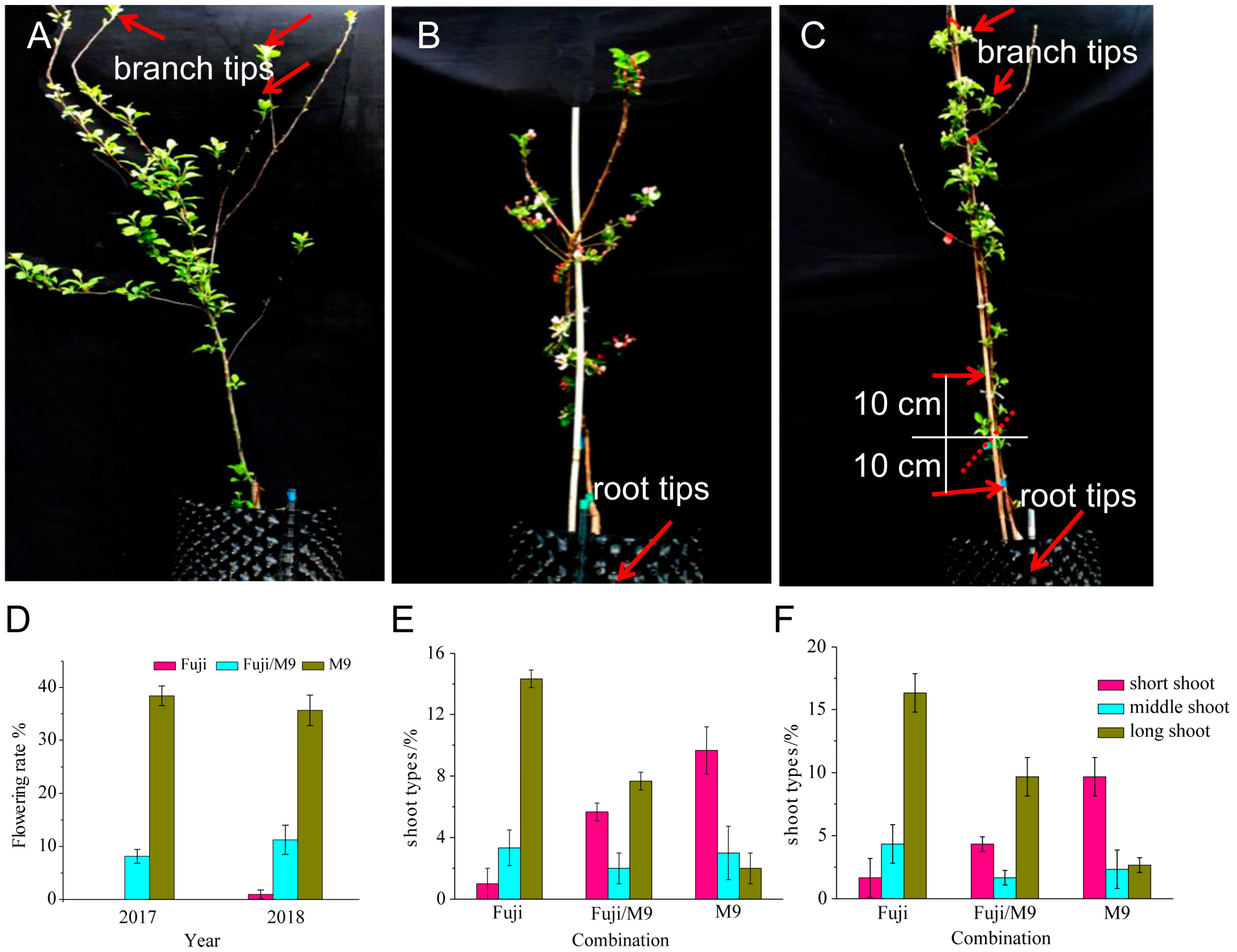
2.1. Differences in Phenotypes among Three Apple Materials
2.2. Differences in Leaf and Bud Mineral and Sugar Contents among Different Apple Trees
2.3. Evaluation of miRNA Sequencing Data
2.4. Identification of Known and Novel miRNAs and Their Expression Profiles
2.5. Targets of Known miRNAs and Their Expression Patterns in Different Materials
2.6. qRT-PCR Analysis of Candidate Differentially Expressed Genes
3. Discussion
3.1. Flowering Rate and N, P, K, and Soluble Sugar Contents Are Affected by Grafting
3.2. Grafting-Induced miRNAs Involved in Bud Growth and Flower Induction
3.3. Regulatory Network of miRNAs–mRNAs in Induction of Flowering
4. Materials and Methods
4.1. Plant Materials
4.2. Determination of Total Nitrogen, Phosphorus, and Potassium Contents
4.3. Determination of Soluble Sugar and Starch Contents
4.4. Construction and Sequencing of sRNA Libraries
4.5. Analysis of High-Throughput Sequencing Data
4.6. Verification of Expression of Differentially Expressed miRNAs and Their Predicted Targets by qRT-PCR
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Koepke, T.; Dhingra, A. Rootstock scion somatogenetic interactions in perennial composite plants. Plant Cell Rep. 2013, 32, 1321–1327. [Google Scholar] [CrossRef] [PubMed]
- Jackson, J.E. World-Wide Development of High Density Planting in Research and Practice. Acta Hortic. 1989, 243, 17–28. [Google Scholar] [CrossRef]
- Polat, M.; Dolgun, O.; YїLdїRїM, A.; AşKїN, M.A.; Gökbayrak, Z. Graft union formation of spur apple varieties grafted on different rootstocks. J. Food Agric. Environ. 2010, 88, 490–493. [Google Scholar]
- El-Wahab, E.H.S.A. Grafting Cucumber onto Some Rootstocks for Controlling Root- Knot Nematodes. Am. Phys. Soc. 2014, 39, 1109–1129. [Google Scholar]
- Li, C.; Li, Y.; Bai, L.; Zhang, T.; He, C.; Yan, Y.; Yu, X. Grafting-responsive miRNAs in cucumber and pumpkin seedlings identified by high-throughput sequencing at whole genome level. Physiol. Plant 2014, 151, 406–422. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Lagos-Quintana, M.; Rauhut, R.; Lendeckel, W.; Tuschl, T. Identification of novel genes coding for small expressed RNAs. Science 2001, 294, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Llave, C.; Kasschau, K.D.; Rector, M.A.; Carrington, J.C. Endogenous and Silencing-Associated Small RNAs in Plants. Plant Cell 2002, 14, 1605–1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, W.; Li, J.; Song, R.; Messing, J.; Chen, X. CARPEL FACTORY, a Dicer Homolog, and HEN1, a Novel Protein, Act in microRNA Metabolism in Arabidopsis thaliana. Curr. Biol. 2002, 12, 1484–1495. [Google Scholar] [CrossRef]
- Bao, N.; Lye, K.-W.; Barton, M.K. MicroRNA Binding Sites in Arabidopsis Class III HD-ZIP mRNAs Are Required for Methylation of the Template Chromosome. Dev. Cell 2004, 7, 653–662. [Google Scholar] [CrossRef] [PubMed]
- Jones-Rhoades, M.W.; Bartel, D.P. Computational Identification of Plant MicroRNAs and Their Targets, Including a Stress-Induced miRNA. Mol. Cell 2004, 14, 787–799. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.; Zhang, D.; Zhao, C.; Li, Y.; Ma, J.; An, N.; Han, M. Shoot bending promotes flower bud formation by miRNA-mediated regulation in apple (Malus domestica Borkh.). Plant Biotechnol. J. 2016, 14, 749–770. [Google Scholar] [CrossRef] [PubMed]
- Pant, B.; Buhtz, A.; Kehr, J.; Scheible, W. MicroRNA399 is a long-distance signal for the regulation of plant phosphate homeostasis. Plant J. 2008, 53, 731–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buhtz, A.; Springer, F.; Chappell, L.; Baulcombe, D.C.; Kehr, J. Identification and characterization of small RNAs from the phloem of Brassica napus. Plant J. Cell. Mol. Biol. 2008, 53, 739–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasai, A.; Kanehira, A.; Harada, T. miR172 can move long distances in Nicotiana benthamiana. Open Plant Sci. J. 2010, 4, 1–6. [Google Scholar] [CrossRef]
- Bhogale, S.; Mahajan, A.S.; Natarajan, B.; Rajabhoj, M.; Thulasiram, H.V.; Banerjee, A.K. MicroRNA156: A Potential Graft-Transmissible MicroRNA That Modulates Plant Architecture and Tuberization in Solanum tuberosum ssp. andigena. Plant Physiol. 2014, 164, 1011–1027. [Google Scholar] [CrossRef] [PubMed]
- Park, M.Y.; Wu, G.; Gonzalez-Sulser, A.; Vaucheret, H.; Poethig, R.S. Nuclear Processing and Export of microRNAs in Arabidopsis. Proc. Natl. Acad. Sci. USA 2005, 102, 3691–3696. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.J.; Lee, J.H.; Kim, W.; Jung, H.S.; Huijser, P.; Ahn, J.H. The microRNA156-SQUAMOSA PROMOTER BINDING PROTEIN-LIKE3 module regulates ambient temperature-responsive flowering via FLOWERING LOCUS T in Arabidopsis. Plant Physiol. 2012, 159, 461–478. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, A.; Wu, M.-F.; Yang, L.; Wu, G.; Poethig, R.S.; Wagner, D. The MicroRNA-Regulated SBP-Box Transcription Factor SPL3 Is a Direct Upstream Activator of LEAFY, FRUITFULL, and APETALA1. Dev. Cell 2009, 17, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Xu, Y.; Shi, M.; Lai, Y.; Wu, X.; Wang, H.; Zhu, Z.; Poethig, R.S.; Wu, G. Repression of miR156 by miR159 Regulates the Timing of the Juvenile-to-Adult Transition in Arabidopsis. Plant Cell 2017, 29, 1293–1304. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Sun, C.; Liu, D.D.; Hao, Y.J.; You, C.X. Ectopic expression of the apple Md-miR172e gene alters flowering time and floral organ identity in Arabidopsis. Plant Cell Tissue Organ Cult. 2015, 123, 535–546. [Google Scholar] [CrossRef]
- Sun, C.; Zhao, Q.; Liu, D.D.; You, C.X.; Hao, Y.J. Ectopic expression of the apple Md-miRNA156h gene regulates flower and fruit development in Arabidopsis. Plant Cell Tissue Organ Cult. 2013, 112, 343–351. [Google Scholar] [CrossRef]
- Keller, M.; Kummer, M.; Vasconcelos, M.C. Reproductive growth of grapevines in response to nitrogen supply and rootstock. Aust. J. Grape Wine Res. 2010, 7, 12–18. [Google Scholar] [CrossRef]
- Samad, A.; Mcneil, D.L.; Khan, Z.U. Effect of interstock bridge grafting (M9 dwarfing rootstock and same cultivar cutting) on vegetative growth, reproductive growth and carbohydrate composition of mature apple trees. Sci. Hortic. 1999, 79, 23–38. [Google Scholar] [CrossRef]
- Lobo, I. Basic Local Alignment Search Tool (BLAST). J. Mol. Biol. 2012, 215, 403–410. [Google Scholar]
- Ma, B.K.; Xu, J.Z.; Sun, J.S. Consideration for high density planting with dwarf rootstocks in apple in China. J. Fruit Sci. 2010, 27, 105–109. [Google Scholar]
- Gonã§Alves, B.; Moutinho-Pereira, J.; Santos, A.; Silva, A.P.; Bacelar, E.; Correia, C.; Rosa, E. Scion-rootstock interaction affects the physiology and fruit quality of sweet cherry. Tree Physiol. 2006, 26, 93–104. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, D.; Rouphael, Y.; Colla, G.; Venema, J.H.; Colla, G. Grafting as a tool to improve tolerance of vegetables to abiotic stresses: Thermal stress, water stress and organic pollutants. Sci. Hortic. 2010, 127, 162–171. [Google Scholar] [CrossRef]
- Shalom, L.; Samuels, S.; Zur, N.; Shlizerman, L.; Doronfaigenboim, A.; Blumwald, E.; Sadka, A. Fruit load induces changes in global gene expression and in abscisic acid (ABA) and indole acetic acid (IAA) homeostasis in citrus buds. J. Exp. Bot. 2014, 65, 3029. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, D.; Xing, L.; Zhang, S.; Zhao, C.; Han, M. Effect of exogenous 6-benzylaminopurine (6-BA) on branch type, floral induction and initiation, and related gene expression in ‘Fuji’ apple (Malus domestica Borkh). Plant Growth Regul. 2016, 79, 65–70. [Google Scholar] [CrossRef]
- Xing, L.B.; Zhang, D.; Li, Y.M.; Shen, Y.W.; Zhao, C.P.; Ma, J.J.; An, N.; Han, M.Y. Transcription Profiles Reveal Sugar and Hormone Signaling Pathways Mediating Flower Induction in Apple (Malus domestica Borkh.). Plant Cell Physiol. 2015, 56, 2052–2068. [Google Scholar] [CrossRef] [PubMed]
- Dadpour, M.R.; Movafeghi, A.; Grigorian, W.; Omidi, Y. Determination of floral initiation in Malus domestica: A novel morphogenetic approach. Biol. Plant. 2011, 55, 243–252. [Google Scholar] [CrossRef]
- Devi, K.L.; Singh, U.C. Effect of nitrogen on growth, flowering and yield of tuberose (Polianthes tuberosa L.) cv. Single. J. Ornam. Hortic. 2010, 13, 228–232. [Google Scholar]
- Stephenson, A.G. Flower and Fruit Abortion: Proximate Causes and Ultimate Functions. Ann. Rev. Ecol. Syst. 1981, 12, 253–279. [Google Scholar] [CrossRef]
- Ito, A.; Hayama, H.; Kashimura, Y. Sugar metabolism in buds during flower bud formation: A comparison of two Japanese pear [Pyrus pyrifolia (Burm.) Nak.] cultivars possessing different flowering habits. Sci. Hortic. 2002, 96, 163–175. [Google Scholar] [CrossRef]
- Julian, C.; Herrero, M.; Rodrigo, J. Flower induction and development in fruiting and non-fruiting shoots in relation to fruit set in apricot (Prunus armeniaca L.). Trees 2010, 24, 833–841. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.H.; Ju, Y.; Seo, P.J.; Lee, J.H.; Park, C.M. The SOC1-SPL module integrates photoperiod and gibberellic acid signals to control flowering time in Arabidopsis. Plant J. 2012, 69, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.Y.; Zhang, L.; Li, W.W.; Hu, X.L.; Wang, M.B.; Fan, Y.L.; Zhang, C.Y.; Wang, L. Stress-induced early flowering is mediated by miR169 in Arabidopsis thaliana. J. Exp. Bot. 2014, 65, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Niu, Q.; Qian, M.; Liu, G.; Yang, F.; Teng, Y. A genome-wide identification and characterization of mircoRNAs and their targets in ‘Suli’ pear (Pyrus pyrifolia white pear group). Planta 2013, 238, 1095–1112. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Li, M.; Gao, F.; Li, S.; Zhu, Y.; Yang, P. Identification of Conserved and Novel microRNAs from Liriodendron chinense Floral Tissues. PLoS ONE 2012, 7, e44696. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.W.; Czech, B.; Weigel, D. miR156-regulated SPL transcription factors define an endogenous flowering pathway in Arabidopsis thaliana. Cell 2009, 138, 738–749. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Peral, M.M.; Li, J.; Li, Y.; Allen, R.S.; Schnippenkoetter, W.; Ohms, S.; White, R.G.; Millar, A.A. The MicroRNA159-Regulated GAMYB-like Genes Inhibit Growth and Promote Programmed Cell Death in Arabidopsis. Plant Physiol. 2010, 154, 757–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aukerman, M.J.; Sakai, H. Regulation of flowering time and floral organ identity by a MicroRNA and its APETALA2-like target genes. Plant Cell 2003, 15, 2730–2741. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.; Adam, H.; Díaz-Mendoza, M.; Żurczak, M.; González-Schain, N.D.; Suárez-López, P. Graft-transmissible induction of potato tuberization by the microRNA miR172. Development 2009, 136, 2873–2881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buhtz, A.; Pieritz, J.; Springer, F.; Kehr, J. Phloem small RNAs, nutrient stress responses, and systemic mobility. BMC Plant Biol. 2010, 10, 64. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Lee, I. Regulation and function of SOC1, a flowering pathway integrator. J. Exp. Bot. 2010, 61, 2247–2254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, J.; Lee, H.; Kim, M.; Lee, I. Analysis of flowering pathway integrators in Arabidopsis. Plant Cell Physiol. 2005, 46, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Cavell, A.J. A rapid method for the determination of nitrogen, phosphorus and potassium in plant materials. J. Sci. Food Agric. 2010, 5, 195–200. [Google Scholar] [CrossRef]
- Fan, S.; Zhang, D.; Lei, C.; Chen, H.; Xing, L.; Ma, J.; Zhao, C.; Han, M. Proteome Analyses Using iTRAQ Labeling Reveal Critical Mechanisms in Alternate Bearing Malus prunifolia. J. Proteome Res. 2016, 15, 3602–3616. [Google Scholar] [CrossRef] [PubMed]
- Maness, N. Extraction and Analysis of Soluble Carbohydrates. In Plant Stress Tolerance: Methods and Protocols; Sunkar, R., Ed.; Humana Press: Totowa, NJ, USA, 2010; pp. 341–370. [Google Scholar]
- Li, R.; Yu, C.; Li, Y.; Lam, T.W.; Yiu, S.M.; Kristiansen, K.; Wang, J. SOAP2: An improved ultrafast tool for short read alignment. Bioinformatics 2009, 25, 1966–1967. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Liu, Y.; Yan, H.; You, Q.; Yi, X.; Du, Z.; Xu, W.; Su, Z. agriGO v2.0: A GO analysis toolkit for the agricultural community, 2017 update. Nucleic Acids Res. 2017, 45, W122–W129. [Google Scholar] [CrossRef] [PubMed]
- Velasco, R.; Zharkikh, A.; Affourtit, J.; Dhingra, A.; Cestaro, A.; Kalyanaraman, A.; Fontana, P.; Bhatnagar, S.K.; Troggio, M.; Pruss, D. The genome of the domesticated apple (Malus × domestica Borkh.). Nat. Genet. 2010, 42, 833–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Zhang, D.; Gao, C.; Zhao, M.; Wu, H.; Li, Y.; Shen, Y.; Han, M. Identification, Classification, and Expression Analysis of GRAS Gene Family in Malus domestica. Front. Physiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.C.; Wang, L.M.; Wang, M.; Song, B.; Tan, S.; Teng, J.F.; Duan, D.X. MicroRNA-195 downregulates Alzheimer’s disease amyloid-β production by targeting BACE1. Brain Res. Bull. 2012, 88, 596–601. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Cultivar | Sampling Time | ||
|---|---|---|---|---|
| May 11, 30 DAFB | July 11, 70 DAFB | Sept. 23, 153 DAFB | ||
| Fresh weight (FW) (g) | Fuji | 52.467 ± 0.321 b | 59.633 ± 1.793 b | 56.400 ± 5.950 c |
| Fuji/M9 | 53.600 ± 4.178 b | 79.167 ± 1.074 a | 75.167 ± 0.500 b | |
| M9 | 57.700 ± 6.146 b | 77.300 ± 1.639 a | 89.767 ± 0.560 a | |
| Dry weight (DW) (g) | Fuji | 18.447 ± 0.185 bc | 21.000 ± 1.472 c | 20.576 ± 2.210 d |
| Fuji/M9 | 17.533 ± 1.330 c | 28.252 ± 1.045 b | 29.208 ± 0.430 c | |
| M9 | 20.933 ± 2.332 ab | 29.433 ± 1.222 b | 37.300 ± 1.351 a | |
| DW/FW | Fuji | 35.161 ± 0.392 a | 35.215 ± 1.612 c | 36.662 ± 0.845 d |
| Fuji/M9 | 32.717 ± 0.285 b | 35.681 ± 0.940 c | 38.857 ± 0.534 c | |
| M9 | 36.267 ± 0.752 a | 38.083 ± 1.577 b | 41.550 ± 1.408 b | |
| Leaf area (cm2) | Fuji | 3127.546 ± 36.797 a | 5022.645 ± 97.551 b | 6476.498 ± 115.639 b |
| Fuji/M9 | 3007.270 ± 58.385 a | 4879.042 ± 73.521 b | 6364.967 ± 139.101 b | |
| M9 | 3100.655 ± 51.934 a | 5938.932 ± 108.548 a | 9090.272 ± 155.033 a | |
| Types | Read Counts | |||||
|---|---|---|---|---|---|---|
| BS | RS | BG | UP | DP | RG | |
| Clean reads | 11,686,623 | 11,590,477 | 11,775,098 | 11,874,809 | 11,663,829 | 11,896,826 |
| Reads mapped to genome | 3,667,388 | 3,335,867 | 3,580,308 | 3,546,267 | 3,756,158 | 2,954,069 |
| rRNA snRNA snoRNA tRNA | 637,318 | 457,197 | 367,837 | 278,159 | 427,007 | 540,889 |
| miRNA | 434,003 | 256,294 | 139,371 | 236,376 | 194,101 | 326,400 |
| Known miRNAs | Targets | |||
|---|---|---|---|---|
| Gene ID | GO | Arabidopsis ID | Description | |
| miR156 | MD06G1138800 | GO:0003677, GO:0005634 | AT5G50670.1 | SPL |
| MD06G1204000 | GO:0003677, GO:0005634 | AT1G69170.1 | ||
| MD07G1111600 | GO:0003677, GO:0005634 | AT1G27360.1 | ||
| MD09G1008900 | GO:0003677, GO:0005634 | AT5G50670.1 | ||
| MD12G1060200 | GO:0003677, GO:0005634 | AT2G42200.1 | ||
| MD13G1059500 | GO:0003677, GO:0005634 | AT1G69170.1 | ||
| MD13G1120300 | GO:0003677, GO:0005634 | AT5G43270.1 | ||
| MD14G1060200 | GO:0003677, GO:0005634 | AT2G42200.1 | ||
| MD14G1215300 | GO:0003677, GO:0005634 | AT1G69170.1 | ||
| MD16G1058800 | GO:0003677, GO:0005634 | AT1G69170.1 | ||
| MD16G1122000 | GO:0003677, GO:0005634 | AT5G50670.1 | ||
| MD17G1001400 | GO:0003677, GO:0005634 | AT5G50670.1 | ||
| miR159 | MD01G1178100 | GO:0003677 | AT3G60460.1 | MYB |
| MD07G1085000 | GO:0003677 | AT3G11440.1 | ||
| MD07G1101000 | GO:0003677 | AT3G60460.1 | ||
| MD11G1215600 | GO:0003677 | AT4G26930.1 | ||
| MD11G1216000 | GO:0003677 | AT2G32460.1 | ||
| miR171 | MD00G1043300 | AT4G00150.1 | GRAS | |
| MD02G1210200 | AT4G00150.1 | |||
| MD04G1088100 | AT4G00150.1 | |||
| MD07G1115800 | AT2G45160.1 | |||
| MD08G1167600 | AT4G08250.1 | |||
| MD15G1353200 | AT4G08250.1 | |||
| miR172 | MD01G1113400 | GO:0003700, GO:0006355 | AT2G28550.1 | AP2 |
| MD03G1107900 | GO:0003700, GO:0006355 | AT2G28550.3 | ||
| MD04G1105200 | GO:0003700, GO:0006355 | AT2G28550.3 | ||
| MD07G1180900 | GO:0003700, GO:0006355 | AT2G28550.2 | ||
| MD11G1121200 | GO:0003700, GO:0006355 | AT2G28550.3 | ||
| MD12G1125900 | GO:0003700, GO:0006355 | AT2G28550.3 | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
An, N.; Fan, S.; Yang, Y.; Chen, X.; Dong, F.; Wang, Y.; Xing, L.; Zhao, C.; Han, M. Identification and Characterization of miRNAs in Self-Rooted and Grafted Malus Reveals Critical Networks Associated with Flowering. Int. J. Mol. Sci. 2018, 19, 2384. https://doi.org/10.3390/ijms19082384
An N, Fan S, Yang Y, Chen X, Dong F, Wang Y, Xing L, Zhao C, Han M. Identification and Characterization of miRNAs in Self-Rooted and Grafted Malus Reveals Critical Networks Associated with Flowering. International Journal of Molecular Sciences. 2018; 19(8):2384. https://doi.org/10.3390/ijms19082384
Chicago/Turabian StyleAn, Na, Sheng Fan, Yang Yang, Xilong Chen, Feng Dong, Yibin Wang, Libo Xing, Caiping Zhao, and Mingyu Han. 2018. "Identification and Characterization of miRNAs in Self-Rooted and Grafted Malus Reveals Critical Networks Associated with Flowering" International Journal of Molecular Sciences 19, no. 8: 2384. https://doi.org/10.3390/ijms19082384
APA StyleAn, N., Fan, S., Yang, Y., Chen, X., Dong, F., Wang, Y., Xing, L., Zhao, C., & Han, M. (2018). Identification and Characterization of miRNAs in Self-Rooted and Grafted Malus Reveals Critical Networks Associated with Flowering. International Journal of Molecular Sciences, 19(8), 2384. https://doi.org/10.3390/ijms19082384