Calcitriol and Calcidiol Can Sensitize Melanoma Cells to Low–LET Proton Beam Irradiation

 ,
,  , and
, and 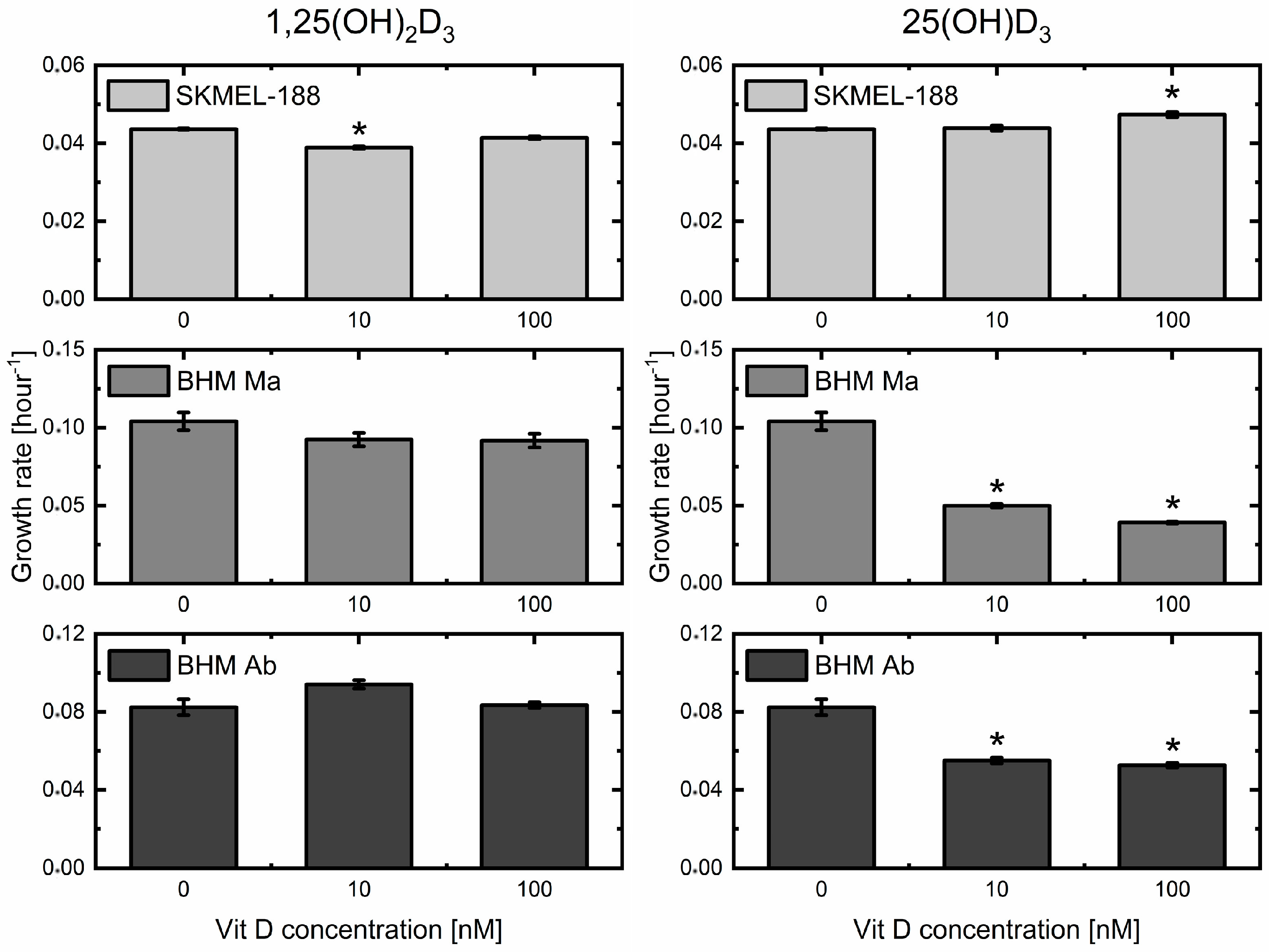{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Impact of Two Metabolites of Vitamin D3 on Melanoma Cell Proliferation
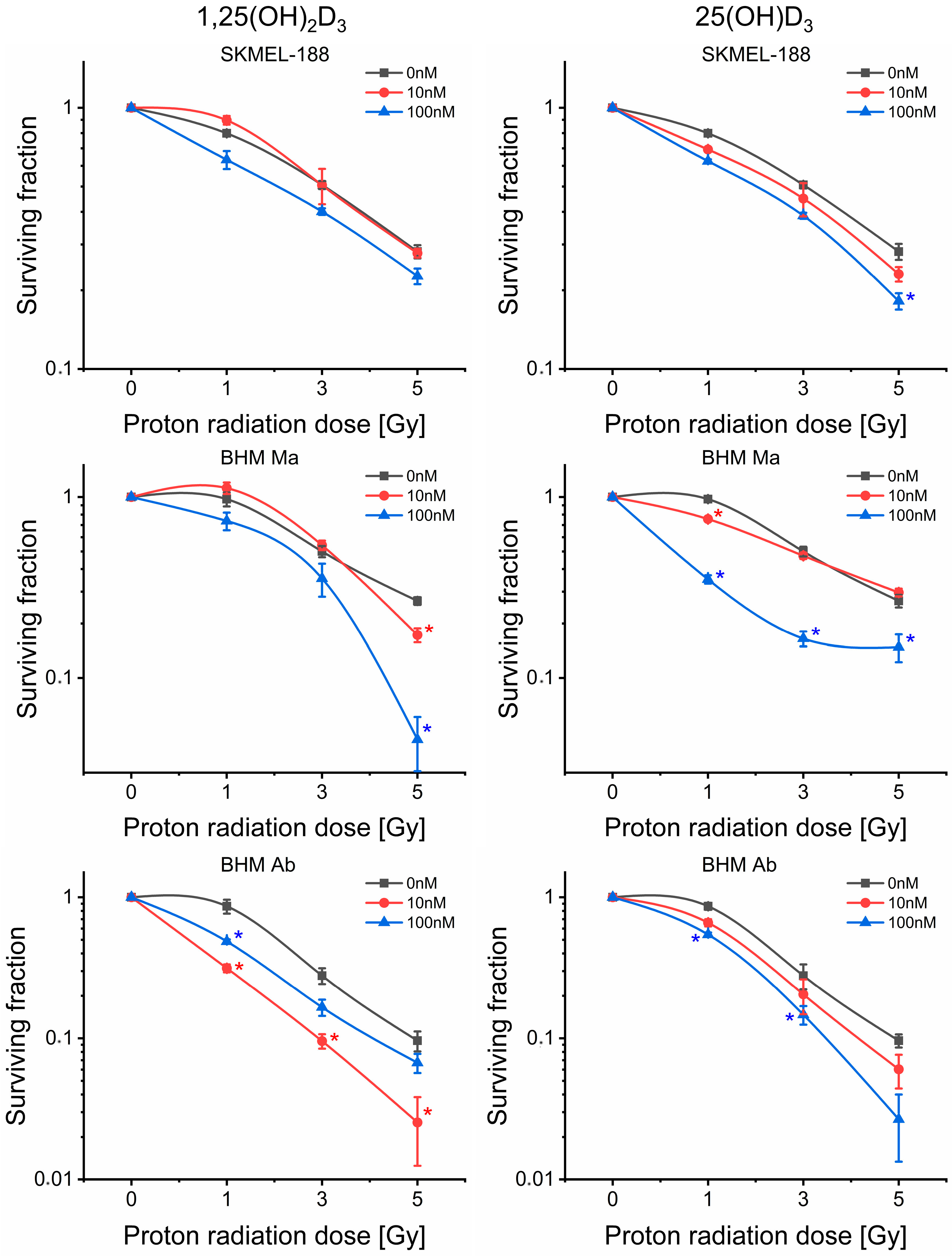
2.2. Proton Beam Radiosensitivity of Melanoma Cells
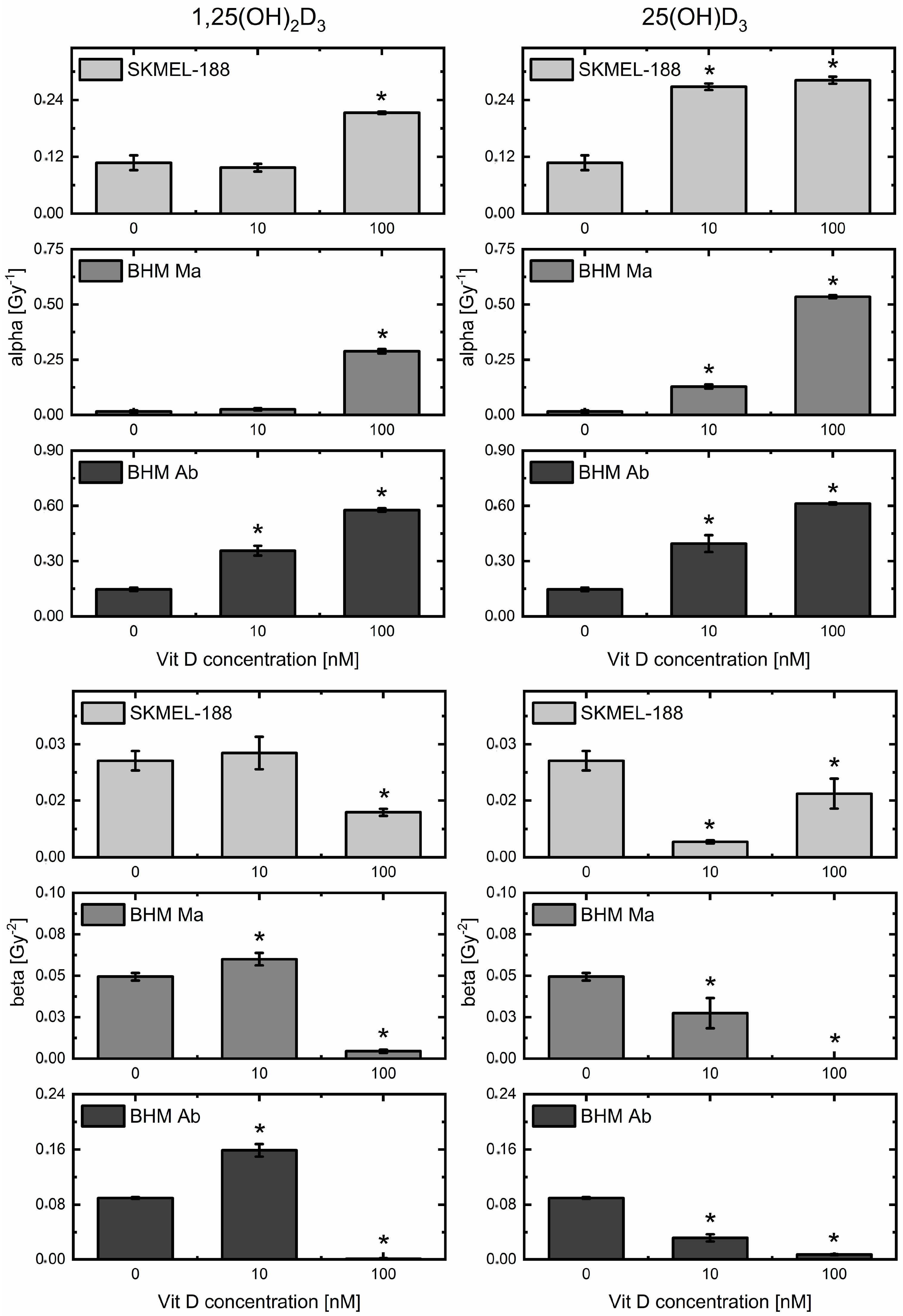
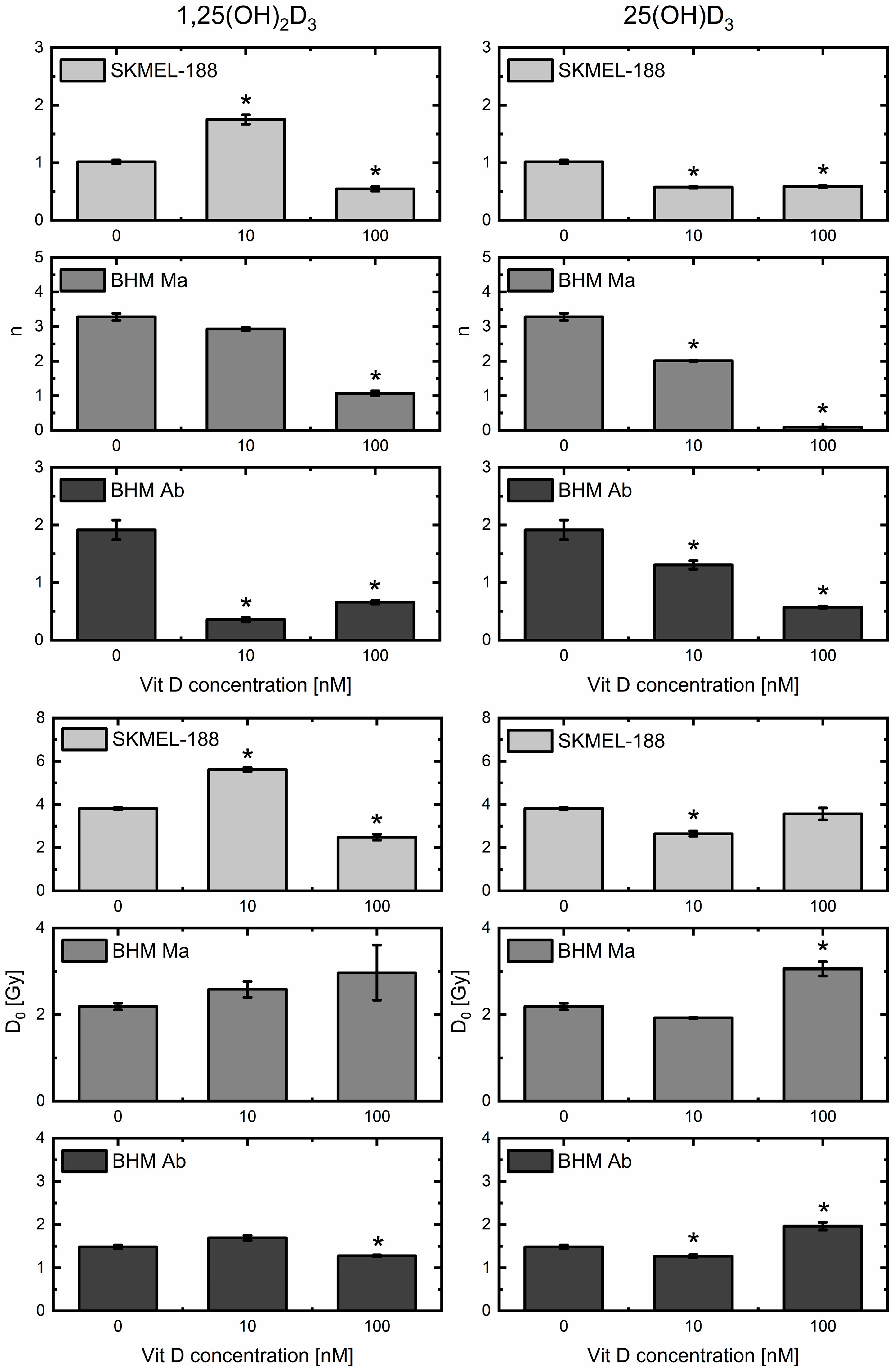
2.3. Vitamin D3 Derivatives Influence Proton Beam Radiosensitivity of Melanoma Cells
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. Cell Culture
4.3. Irradiation Procedure
4.4. Data Analysis
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ATM | Ataxia-telangiectasia mutated kinase |
| BHM | Bomirski hamster melanoma |
| CSDA | Continues slowing down approximation |
| DSB | Double strand break |
| IR | Ionizing radiation |
| LQ | Linear-quadratic |
| mTor | Mammalian target of rapamycin |
| MTSH | Multitarget—single hit |
| SF | Surviving fraction |
| SOBP | Spread-out bragg peak |
| SSB | Single strand break |
| VDR | Vitamin D receptor |
References
- Matthews, N.H.; Li, W.-Q.; Qureshi, A.A.; Weinstock, M.A.; Cho, E. Epidemiology of Melanoma. In Cutaneous Melanoma: Etiology and Therapy; Codon Publications: Brisbane, Australia, 2017. [Google Scholar]
- Slominski, A.T.; Carlson, J.A. Melanoma resistance: A bright future for academicians and a challenge for patient advocates. Mayo Clin. Proc. 2014, 89, 429–433. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, S.; Watson, I.R. Molecular characterisation of cutaneous melanoma: Creating a framework for targeted and immune therapies. Br. J. Cancer 2016, 115, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Su, M.Y.; Fisher, D.E. Immunotherapy in the Precision Medicine Era: Melanoma and Beyond. PLoS Med. 2016, 13, e1002196. [Google Scholar] [CrossRef] [PubMed]
- Levesque, M.P.; Cheng, P.F.; Raaijmakers, M.I.G.; Saltari, A.; Dummer, R. Metastatic melanoma moves on: Translational science in the era of personalized medicine. Cancer Metastasis Rev. 2017, 36, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Ordonez-Moran, P.; Larriba, M.J.; Pendas-Franco, N.; Aguilera, O.; Gonzalez-Sancho, J.M.; Munoz, A. Vitamin D and cancer: An update of in vitro and in vivo data. Front. Biosci. 2005, 10, 2723–2749. [Google Scholar] [CrossRef] [PubMed]
- Feldman, D.; Krishnan, A.V.; Swami, S.; Giovannucci, E.; Feldman, B.J. The role of vitamin D in reducing cancer risk and progression. Nat. Rev. Cancer 2014, 14, 342–357. [Google Scholar] [CrossRef] [PubMed]
- Grant, W.B. Roles of Solar UVB and Vitamin D in Reducing Cancer Risk and Increasing Survival. Anticancer Res. 2016, 36, 1357–1370. [Google Scholar] [PubMed]
- Slominski, A.T.; Brozyna, A.A.; Zmijewski, M.A.; Józwicki, W.; Jetten, A.M.; Mason, R.S.; Tuckey, R.C.; Elmets, C.A. Vitamin D signalling and melanoma: Role of Vitamin D and its receptors in melanoma progression and management. Lab. Investig. 2017, 97, 706–724. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Brożyna, A.A.; Skobowiat, C.; Zmijewski, M.A.; Kim, T.K.; Janjetovic, Z.; Oak, A.S.; Jozwicki, W.; Jetten, A.M.; Mason, R.S.; et al. On the role of classical and novel forms of vitamin D in melanoma progression and management. J. Steroid Biochem. Mol. Biol. 2018, 177, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Wasiewicz, T.; Szyszka, P.; Cichorek, M.; Janjetovic, Z.; Tuckey, R.C.; Slominski, A.T.; Zmijewski, M.A. Antitumor effects of vitamin D analogues on hamster and mouse melanoma cell lines in relation to melanin pigmentation. Int. J. Mol. Sci. 2015, 16, 6645–6667. [Google Scholar] [CrossRef] [PubMed]
- Polar, M.K.; Gennings, C.; Park, M.; Gupta, M.S.; Gewirtz, D.A. Effect of the vitamin D3 analog ILX 23-7553 on apoptosis and sensitivity to fractionated radiation in breast tumour cells and normal human fibroblasts. Cancer Chemother. Pharmacol. 2003, 51, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Sundaram, S.; Sea, A.; Feldman, S.; Strawbridge, R.; Hoopes, P.J.; Demidenko, E.; Binderup, L.; Gewirtz, D.A. The combination of a potent vitamin D3 analog, EB 1089, with ionizing radiation reduces tumour growth and induces apoptosis of MCF-7 breast tumour xenografts in nude mice. Clin. Cancer Res. 2003, 9, 2350–2356. [Google Scholar] [PubMed]
- Wang, Z.; Liu, J.; Si, S.; Xu, B.; Wang, Y.; Ren, Y.; Dai, Z.; Cui, D.; Su, X.; Song, S. Vitamin D improves the sensitization of lung cancer to radiotherapy. Int. J. Clin. Exp. Med. 2017, 10, 4921–4928. [Google Scholar]
- Sundaram, S.; Gewirtz, D.A. The vitamin D3 analog EB 1089 enhances the response of human breast tumor cells to radiation. Radiat. Res. 1999, 152, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, M.; Sundaram, S.; Gennings, C.; Carter, H.; Gewirtz, D.A. The vitamin D3 analog, ILX-23-7553, enhances the response to Adriamycin and irradiation in MCF-7 breast tumor cells. Cancer Chemother. Pharmacol. 2001, 47, 429–436. [Google Scholar] [CrossRef]
- Weitsman, G.E.; Koren, R.; Zuck, E.; Rotem, C.; Liberman, U.A.; Ravid, A. Vitamin D sensitizes breast cancer cells to the action of H2O2: Mitochondria as a convergence point in the death pathway. Free Radic. Biol. Med. 2005, 39, 266–278. [Google Scholar] [CrossRef] [PubMed]
- Dunlap, N.; Schwartz, G.G.; Eads, D.; Cramer, S.D.; Sherk, A.B.; John, V.; Koumenis, C. 1α,25-Dihydroxyvitamin D3 (calcitriol) and its analogue, 19-nor-1α,25(OH)2D2, potentiate the effects of ionising radiation on human prostate cancer cells. Br. J. Cancer 2003, 89, 746–753. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Brozyna, A.; Jozwicki, W.; Tuckey, R.C. Vitamin D as an adjuvant in melanoma therapy. Melanoma Manag. 2015, 2, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Beilhartz, G.; Roy, Y.; Richard, C.L.; Curtin, M.; Brown, L.; Cadieux, D.; Coppolino, M.; Farach-Carson, M.C.; Nemere, I.; et al. Nuclear translocation of the 1,25D3-MARRS (membrane associated rapid response to steroids) receptor protein and NFκB in differentiating NB4 leukemia cells. Exp. Cell Res. 2010, 316, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; De Noronha, R.G.; Roecker, A.J.; Pyrzynska, B.; Khwaja, F.; Zhang, Z.; Zhang, H.; Teng, Q.; Nicholson, A.C.; Giannakakou, P.; et al. Identification of a novel small-molecule inhibitor of the hypoxia-inducible factor 1 pathway. Cancer Res. 2005, 65, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Skobowiat, C.; Oak, A.S.W.; Kim, T.-K.; Yang, C.H.; Pfeffer, L.M.; Tuckey, R.C.; Slominski, A.T. Noncalcemic 20-hydroxyvitamin D3 inhibits human melanoma growth in in vitro and in vivo models. Oncotarget 2016, 8, 9823–9834. [Google Scholar] [CrossRef] [PubMed]
- Brozyna, A.A.; Jozwicki, W.; Janjetovic, Z.; Slominski, A.T. Expression of vitamin D receptor decreases during progression of pigmented skin lesions. Hum. Pathol. 2011, 42, 618–631. [Google Scholar] [CrossRef] [PubMed]
- Brozyna, A.A.; Jóźwicki, W.; Janjetovic, Z.; Slominski, A.T. Expression of the vitamin D-activating enzyme 1α-hydroxylase (CYP27B1) decreases during melanoma progression. Hum. Pathol. 2013, 44, 374–387. [Google Scholar] [CrossRef] [PubMed]
- Brozyna, A.A.; Jóźwicki, W.; Slominski, A.T. Decreased VDR expression in cutaneous melanomas as marker of tumor progression: New data and analyses. Anticancer Res. 2014, 34, 2735–2744. [Google Scholar] [PubMed]
- Brożyna, A.A.; Jochymski, C.; Janjetovic, Z.; Jóźwicki, W.; Tuckey, R.C.; Slominski, A.T. CYP24A1 expression inversely correlates with melanoma progression: Clinic-pathological studies. Int. J. Mol. Sci. 2014, 15, 19000–19017. [Google Scholar] [CrossRef] [PubMed]
- Chagani, S.; Kyryachenko, S.; Yamamoto, Y.; Kato, S.; Ganguli-Indra, G.; Indra, A.K. In vivo Role of Vitamin D Receptor Signaling in UVB-Induced DNA Damage and Melanocyte Homeostasis. J. Investig. Dermatol. 2016, 136, 2108–2111. [Google Scholar] [CrossRef] [PubMed]
- Girdhani, S.; Sachs, R.; Hlatky, L. Biological Effects of Proton Radiation: What We Know and Don’t Know. Radiat. Res. 2013, 179, 257–272. [Google Scholar] [CrossRef] [PubMed]
- Gerelchuluun, A.; Hong, Z.; Sun, L.; Suzuki, K.; Terunuma, T.; Yasuoka, K.; Sakae, T.; Moritake, T.; Tsuboi, K. Induction of in situ DNA double-strand breaks and apoptosis by 200 MeV protons and 10 MV X-rays in human tumour cell lines. Int. J. Radiat. Biol. 2011, 87, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Jasińska-Konior, K.; Pochylczuk, K.; Czajka, E.; Michalik, M.; Romanowska-Dixon, B.B.; Swakoń, J.; Urbańska, K.; Elas, M.; Jasinska-Konior, K.; Pochylczuk, K.; et al. Proton beam irradiation inhibits the migration of melanoma cells. PLoS ONE 2017, 12, e0186002. [Google Scholar] [CrossRef] [PubMed]
- Grosse, N.; Fontana, A.O.; Hug, E.B.; Lomax, A.; Coray, A.; Augsburger, M.; Paganetti, H.; Sartori, A.A.; Pruschy, M. Deficiency in homologous recombination renders mammalian cells more sensitive to proton versus photon irradiation. Int. J. Radiat. Oncol. Biol. Phys. 2014, 88, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Antoccia, A.; Sgura, A.; Berardinelli, F.; Cavinato, M.; Cherubini, R.; Gerardi, S.; Tanzarella, C. Cell Cycle Perturbations and Genotoxic Effects in Human Primary Fibroblasts Induced by Low-energy Protons and X/γ-rays. J. Radiat. Res. 2009, 50, 457–468. [Google Scholar] [CrossRef] [PubMed]
- Śniegocka, M.; Podgórska, E.; Plonka, P.M.; Elas, M.; Romanowska-Dixon, B.; Szczygiel, M.; Żmijewski, M.A.; Cichorek, M.; Markiewicz, A.; Brożyna, A.A.; et al. Transplantable melanomas in hamsters and gerbils as models for human melanoma. Sensitization in melanoma radiotherapy—From animal models to clinical trials. Int. J. Mol. Sci. 2018, 19, 1048. [Google Scholar] [CrossRef] [PubMed]
- Buch, K.; Peters, T.; Nawroth, T.; Sänger, M.; Schmidberger, H.; Langguth, P. Determination of cell survival after irradiation via clonogenic assay versus multiple MTT Assay—A comparative study. Radiat. Oncol. 2012, 7, 1. [Google Scholar] [CrossRef] [PubMed]
- Franken, N.A.P.; Oei, A.L.; Kok, H.P.; Rodermond, H.M.; Sminia, P.; Crezee, J.; Stalpers, L.J.A.; Barendsen, G.W. Cell survival and radiosensitisation: Modulation of the linear and quadratic parameters of the LQ model (Review). Int. J. Oncol. 2013, 42, 1501–1515. [Google Scholar] [CrossRef] [PubMed]
- Bodgi, L.; Canet, A.; Pujo-Menjouet, L.; Lesne, A.; Victor, J.-M.; Foray, N. Mathematical models of radiation action on living cells: From the target theory to the modern approaches. A historical and critical review. J. Theor. Biol. 2016, 394, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Mehta, R.G.; Hussain, E.A.; Mehta, R.R.; Das Gupta, T.K. Chemoprevention of mammary carcinogenesis by 1α-hydroxyvitamin D5, a synthetic analog of Vitamin D. Mutat. Res. 2003, 523–524, 253–264. [Google Scholar] [CrossRef]
- Welsh, J.; Wietzke, J.A.; Zinser, G.M.; Byrne, B.; Smith, K.; Narvaez, C.J. Vitamin D-3 Receptor as a Target for Breast Cancer Prevention. J. Nutr. 2003, 133, 2425S–2433S. [Google Scholar] [CrossRef] [PubMed]
- Lamprecht, S.A.; Lipkin, M. Chemoprevention of colon cancer by calcium, vitamin D and folate: Molecular mechanisms. Nat. Rev. Cancer 2003, 3, 601–614. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, K. 1,25-Dihydroxyvitamin D3 is a preventive factor in the metastasis of lung cancer. Carcinogenesis 2004, 26, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, A.V.; Medicine, S.; Feldman, D. Vitamin D Analogs in Cancer Prevention and Therapy. 2003, 164. [Google Scholar] [CrossRef]
- Szyszka, P.; Zmijewski, M.A.; Slominski, A.T. New vitamin D analogs as potential therapeutics in melanoma. Expert Rev. Anticancer Ther. 2012, 12, 585–599. [Google Scholar] [CrossRef] [PubMed]
- Moreno, J.; Krishnan, A.V.; Feldman, D. Molecular mechanisms mediating the anti-proliferative effects of Vitamin D in prostate cancer. J. Steroid Biochem. Mol. Biol. 2005, 97, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, P.; Chatterjee, M. Antiproliferative role of vitamin D and its analogs—A brief overview. Mol. Cell. Biochem. 2003, 253, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Thill, M.; Woeste, A.; Reichert, K.; Fischer, D.; Rody, A.; Friedrich, M.; Köster, F. Vitamin D inhibits ovarian cancer cell line proliferation in combination with celecoxib and suppresses cyclooxygenase-2 expression. Anticancer Res. 2015, 35, 1197–1203. [Google Scholar] [PubMed]
- Chen, J.; Wang, J.; Kim, T.K.; Tieu, E.W.; Tang, E.K.Y.; Lin, Z.; Kovacic, D.; Miller, D.D.; Postlethwaite, A.; Tuckey, R.C.; et al. Novel vitamin D analogs as potential therapeutics: Metabolism, toxicity profiling and antiproliferative activity. Anticancer Res. 2014, 34, 2153–2164. [Google Scholar] [PubMed]
- Kozielewicz, P.; Grafton, G.; Kutner, A.; Curnow, S.J.; Gordon, J.; Barnes, N.M. Novel vitamin D analogues; cytotoxic and anti-proliferative activity against a diffuse large B-cell lymphoma cell line and B-cells from healthy donors. J. Steroid Biochem. Mol. Biol. 2016, 164, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Humeniuk-Polaczek, R.; Marcinkowska, E. Impaired nuclear localization of vitamin D receptor in leukemia cells resistant to calcitriol-induced differentiation. J. Steroid Biochem. Mol. Biol. 2004, 88, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Agadir, A.; Lazzaro, G.; Zheng, Y.; Zhang, X.-K.; Mehta, R. Resistance of HBL100 human breast epithelial cells to vitamin D action. Carcinogenesis 1999, 20, 577–582. [Google Scholar] [CrossRef] [PubMed]
- Seifert, M.; Rech, M.; Meineke, V.; Tilgen, W.; Reichrath, J. Differential biological effects of 1,25-dihydroxyVitamin D3 on melanoma cell lines in vitro. J. Steroid Biochem. Mol. Biol. 2004, 89–90, 375–379. [Google Scholar] [CrossRef] [PubMed]
- Reichrath, J.; Rech, M.; Moeini, M.; Meese, E.; Tilgen, W.; Seifert, M. In vitro comparison of the vitamin D endocrine system in 1,25(OH)2D3-responsive and -resistant melanoma cells. Cancer Biol. Ther. 2007, 6, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Janjetovic, Z.; Kim, T.K.; Wright, A.C.; Grese, L.N.; Riney, S.J.; Nguyen, M.N.; Tuckey, R.C. Novel vitamin D hydroxyderivatives inhibit melanoma growth and show differential effects on normal melanocytes. Anticancer Res. 2012, 32, 3733–3742. [Google Scholar] [PubMed]
- Sun, J.; Yin, M.; Zhu, S.; Liu, L.; Zhu, Y.; Wang, Z.; Xu, R.X.; Chang, S. Ultrasound-mediated destruction of oxygen and paclitaxel loaded lipid microbubbles for combination therapy in hypoxic ovarian cancer cells. Ultrason. Sonochem. 2016, 28, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Janjetovic, Z.; Brozyna, A.A.; Tuckey, R.C.; Kim, T.K.; Nguyen, M.N.; Jozwicki, W.; Pfeffer, S.R.; Pfeffer, L.M.; Slominski, A.T. High basal NF-κB activity in nonpigmented melanoma cells is associated with an enhanced sensitivity to vitamin D3 derivatives. Br. J. Cancer 2011, 105, 1874–1884. [Google Scholar] [CrossRef] [PubMed]
- Sarna, M.; Zadlo, A.; Czuba-Pelech, B.; Urbanska, K. Nanomechanical phenotype of melanoma cells depends solely on the amount of endogenous pigment in the cells. Int. J. Mol. Sci. 2018, 19, 607. [Google Scholar] [CrossRef] [PubMed]
- Wietrzyk, J.; Nevozhay, D.; Filip, B.; Milczarek, M.; Kutner, A. The antitumor effect of lowered doses of cytostatics combined with new analogs of vitamin D in mice. Anticancer Res. 2007, 27, 3387–3398. [Google Scholar] [PubMed]
- Pelczynska, M.; Switalska, M.; Maciejewska, M.; Jaroszewicz, I.; Kutner, A.; Opolski, A. Antiproliferative activity of vitamin D compounds in combination with cytostatics. Anticancer Res. 2006, 26, 2701–2705. [Google Scholar] [PubMed]
- Ma, Y.; Yu, W.D.; Trump, D.L.; Johnson, C.S. 1,25D3 enhances antitumor activity of gemcitabine and cisplatin in human bladder cancer models. Cancer 2010, 116, 3294–3303. [Google Scholar] [CrossRef] [PubMed]
- Newhauser, W.D.; Zhang, R. The physics of proton therapy. Phys. Med. Biol. 2015, 60, R155–R209. [Google Scholar] [CrossRef] [PubMed]
- Fokas, E.; Kraft, G.; An, H.; Engenhart-Cabillic, R. Ion beam radiobiology and cancer: Time to update ourselves. Biochim. Biophys. Acta Rev. Cancer 2009, 1796, 216–229. [Google Scholar] [CrossRef] [PubMed]
- Folkard, M.; Prise, K.M.; Vojnovic, B.; Newman, H.C.; Roper, M.J.; Michael, B.D. Inactivation of V79 cells by low-energy protons, deuterons and helium-3 ions. Int. J. Radiat. Biol. 1996, 69, 729–738. [Google Scholar] [CrossRef] [PubMed]
- Kundrät, P. Detailed analysis of the cell-inactivation mechanism by accelerated protons and light ions. Phys. Med. Biol. 2006, 51, 1185–1199. [Google Scholar] [CrossRef] [PubMed]
- Kundrát, P.; Lokajíček, M.; Hromčíková, H. Probabilistic two-stage model of cell inactivation by ionizing particles. Phys. Med. Biol. 2005, 50, 1433–1447. [Google Scholar] [CrossRef] [PubMed]
- Trémezaygues, L.; Seifert, M.; Vogt, T.; Tilgen, W.; Reichrath, J. 1,25-Dihydroxyvitamin D3 modulates effects of ionizing radiation (IR) on human keratinocytes: In vitro analysis of cell viability/proliferation, DNA-damage and -repair. J. Steroid Biochem. Mol. Biol. 2010, 121, 324–327. [Google Scholar] [CrossRef] [PubMed]
- Murray, P.J.; Cornelissen, B.; Vallis, K.A.; Chapman, S.J. DNA double-strand break repair: A theoretical framework and its application. J. R. Soc. Interface 2016, 13, 20150679. [Google Scholar] [CrossRef] [PubMed]
- Murray, D.; Mirzayans, R.; Parliament, M. New insights into patient radiosensitivity in relation to the biology of the ATM protein. Transl. Cancer Res. 2016, 5, S809–S814. [Google Scholar] [CrossRef]
- Janczyk, A.; Wolnicka-Glubisz, A.; Chmura, A.; Elas, M.; Matuszak, Z.; Stochel, G.; Urbanska, K. NO-dependent phototoxicity of Roussin’s black salt against cancer cells. Nitric Oxide Biol. Chem. 2004, 10, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Brindell, M.; Kuliś, E.; Elmroth, S.K.C.; Urbańska, K.; Stochel, G. Light-induced anticancer activity of [RuCl2(DMSO)4] complexes. J. Med. Chem. 2005, 48, 7298–7304. [Google Scholar] [CrossRef] [PubMed]
- Bomirski, A.; Słominski, A.; Bigda, J. The natural history of a family of transplantable melanomas in hamsters. Cancer Metastasis Rev. 1988, 7, 95–118. [Google Scholar] [CrossRef] [PubMed]
- Słominski, A. Rapid melanization of Bomirski amelanotic melanoma cells in cell culture. Biosci. Rep. 1983, 3, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Słomiński, A. Some properties of Bomirski Ab amelanotic melanoma cells, which underwent spontaneous melanization in primary cell culture—Growth kinetics, cell morphology, melanin content and tumorigenicity. J. Cancer Res. Clin. Oncol. 1985, 109, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Słominski, A.; Moellmann, G.; Kuklinska, E.; Bomirski, A.; Pawelek, J. Positive regulation of melanin pigmentation by two key substrates of the melanogenic pathway, l-tyrosine and L-dopa. J. Cell Sci. 1988, 89, 287–296. [Google Scholar] [PubMed]
- Słomiński, A.; Bomirski, A. Phenotypic changes of Ab hamster melanoma during long-term culture. Anticancer Res. 1985, 5, 403–409. [Google Scholar] [PubMed]
- Boberek, M.; Swakoń, J.; Stolarczyk, L.; Olko Pawełand Waligórski, M. A monitoring system for the 60 MeV radiotherapy proton beam at IFJ PAN using a scintillating screen and a CCD camera. Rom. Rep. Phys. 2014, 66, 5–15. [Google Scholar]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Podgorska, E.; Drzal, A.; Matuszak, Z.; Swakon, J.; Slominski, A.; Elas, M.; Urbanska, K. Calcitriol and Calcidiol Can Sensitize Melanoma Cells to Low–LET Proton Beam Irradiation. Int. J. Mol. Sci. 2018, 19, 2236. https://doi.org/10.3390/ijms19082236
Podgorska E, Drzal A, Matuszak Z, Swakon J, Slominski A, Elas M, Urbanska K. Calcitriol and Calcidiol Can Sensitize Melanoma Cells to Low–LET Proton Beam Irradiation. International Journal of Molecular Sciences. 2018; 19(8):2236. https://doi.org/10.3390/ijms19082236
Chicago/Turabian StylePodgorska, Ewa, Agnieszka Drzal, Zenon Matuszak, Jan Swakon, Andrzej Slominski, Martyna Elas, and Krystyna Urbanska. 2018. "Calcitriol and Calcidiol Can Sensitize Melanoma Cells to Low–LET Proton Beam Irradiation" International Journal of Molecular Sciences 19, no. 8: 2236. https://doi.org/10.3390/ijms19082236
APA StylePodgorska, E., Drzal, A., Matuszak, Z., Swakon, J., Slominski, A., Elas, M., & Urbanska, K. (2018). Calcitriol and Calcidiol Can Sensitize Melanoma Cells to Low–LET Proton Beam Irradiation. International Journal of Molecular Sciences, 19(8), 2236. https://doi.org/10.3390/ijms19082236