Effects of Funneliformis mosseae on Root Metabolites and Rhizosphere Soil Properties to Continuously-Cropped Soybean in the Potted-Experiments

Abstract
:1. Introduction
2. Results
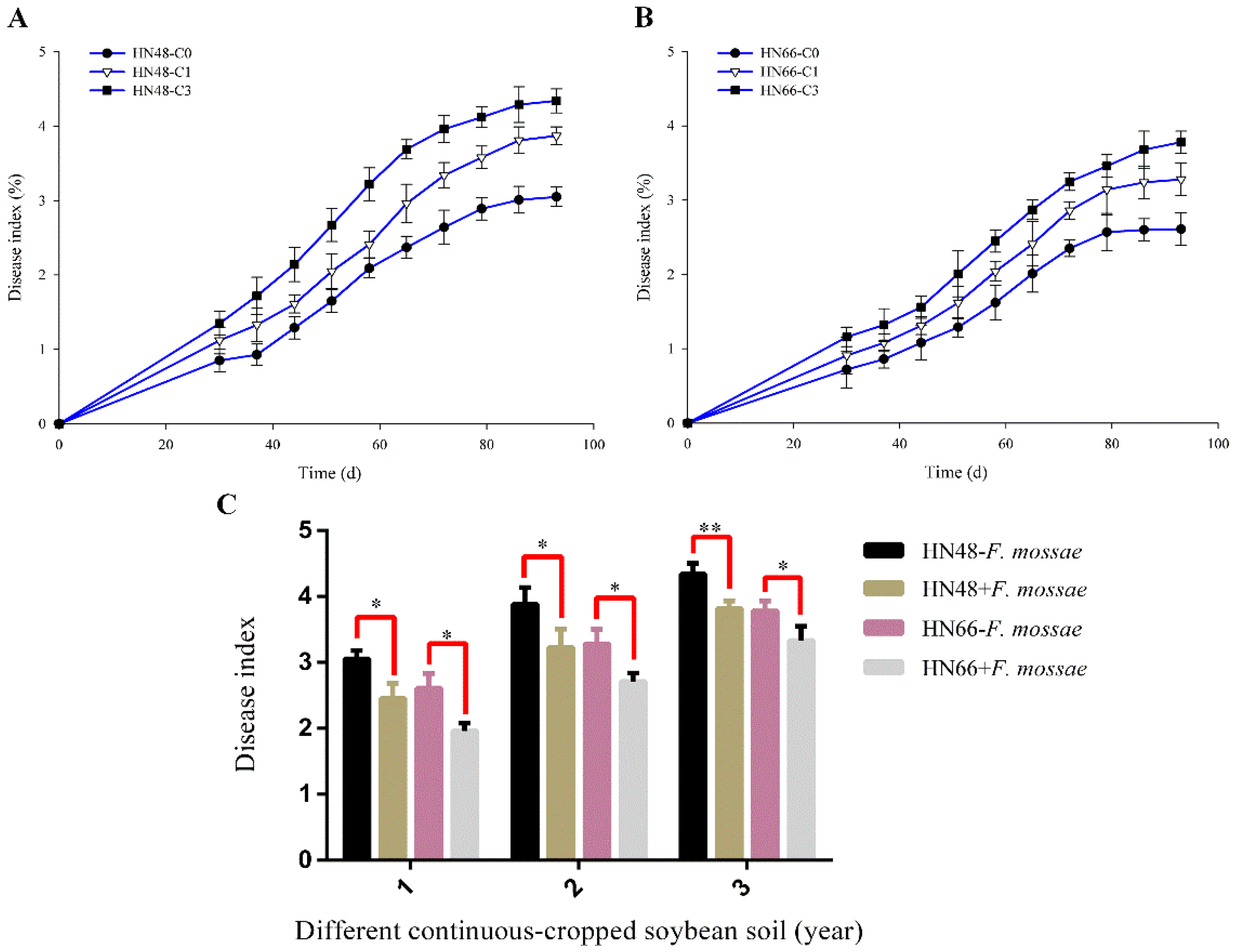
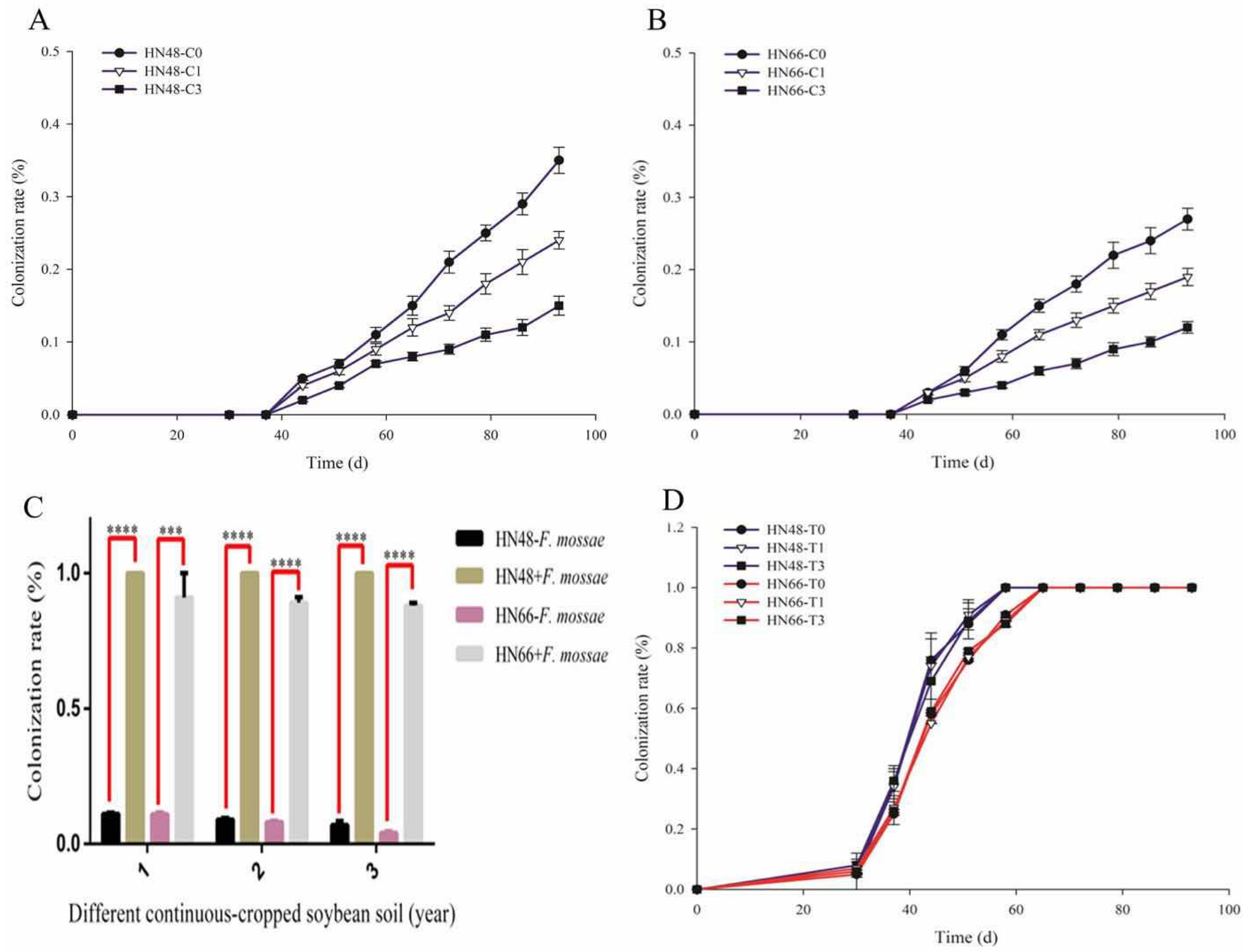
2.1. Effect of F. mosseae on Root Rot Index and the AMF Colonization Rate in Different Conditions Potted-Soybean
2.2. Total DNA Extraction and Specific DNA Fragment Amplification from Root and Rhizosphere Soil
2.3. Metabolite Profiling in Continuously Cropped Soybean under Potted-Experiments
2.3.1. Metabolite Profiling of HN48 and HN66 Root Samples from Continuously Cropped Soybean under Potted-Experiments
2.3.2. Metabolite Profiling of HN48 and HN66 Soil Samples from Continuously Cropped Soybean under Potted-Experiments
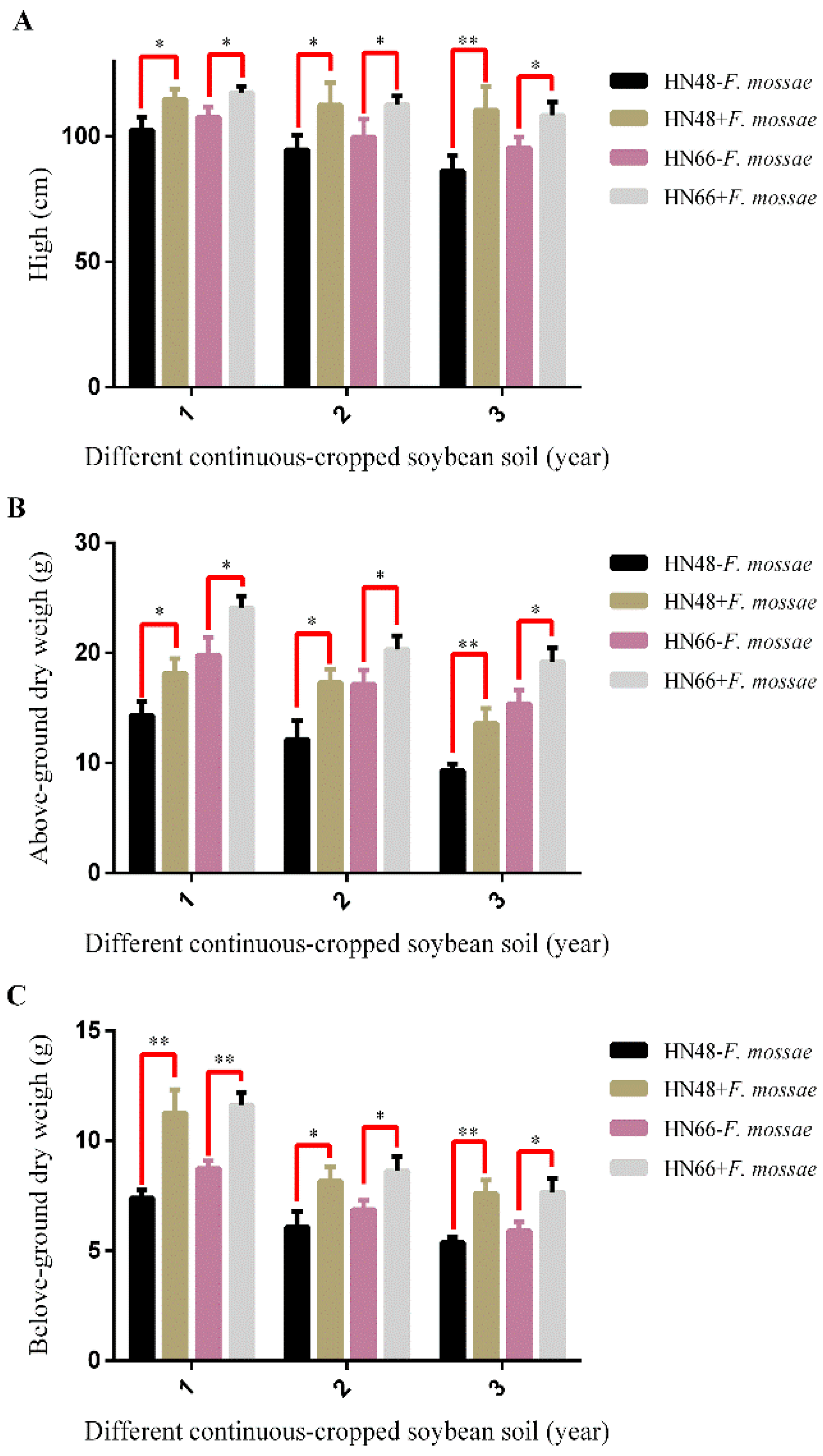
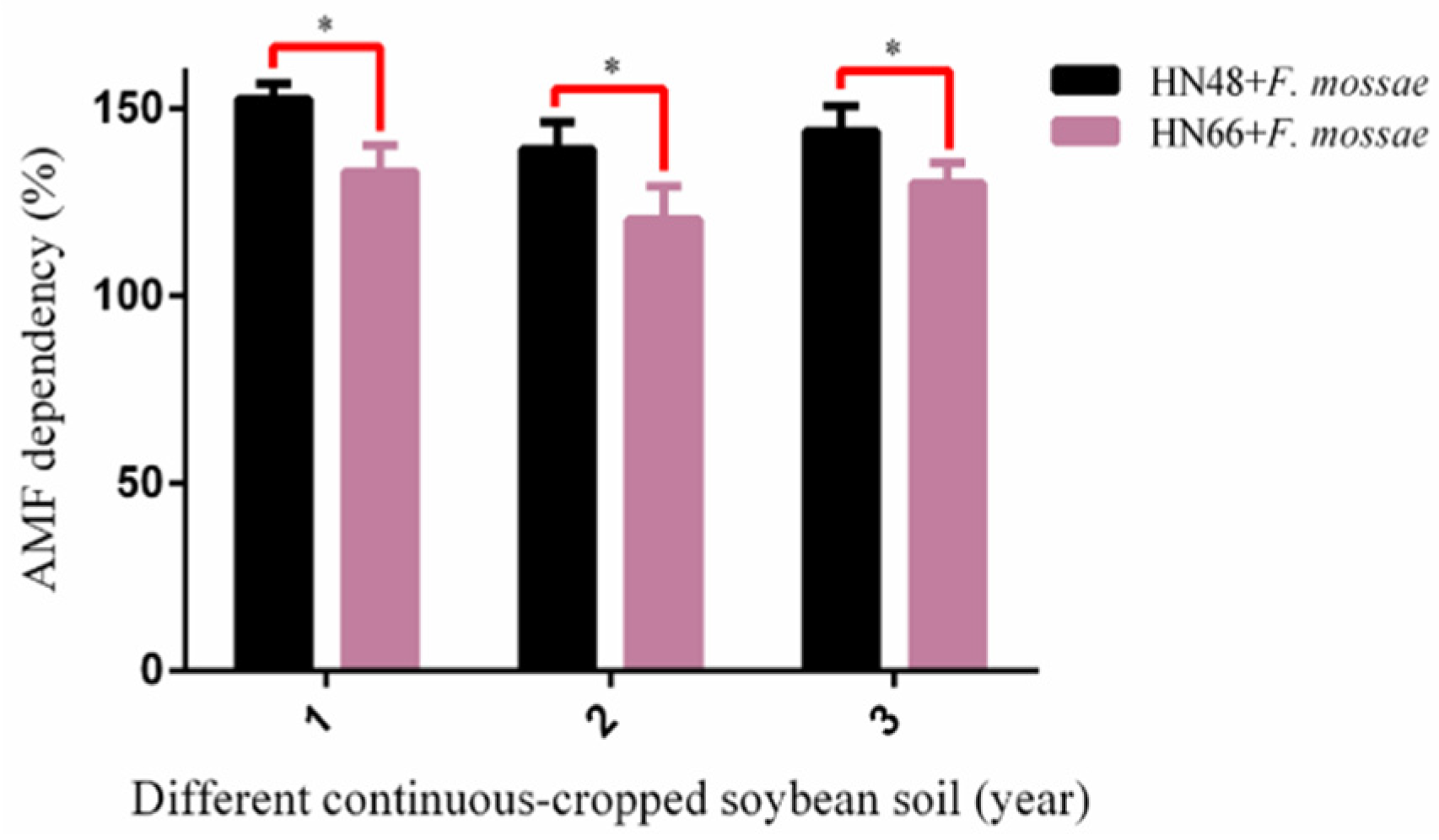
2.4. Determination of Soybean Biomass and Rhizosphere Soil Physicochemical Properties from Continuously Cropped Soybean under Potted-Experiments
2.4.1. Measurement of Biomass in Continuously Cropped Soybean under Potted-Experiments
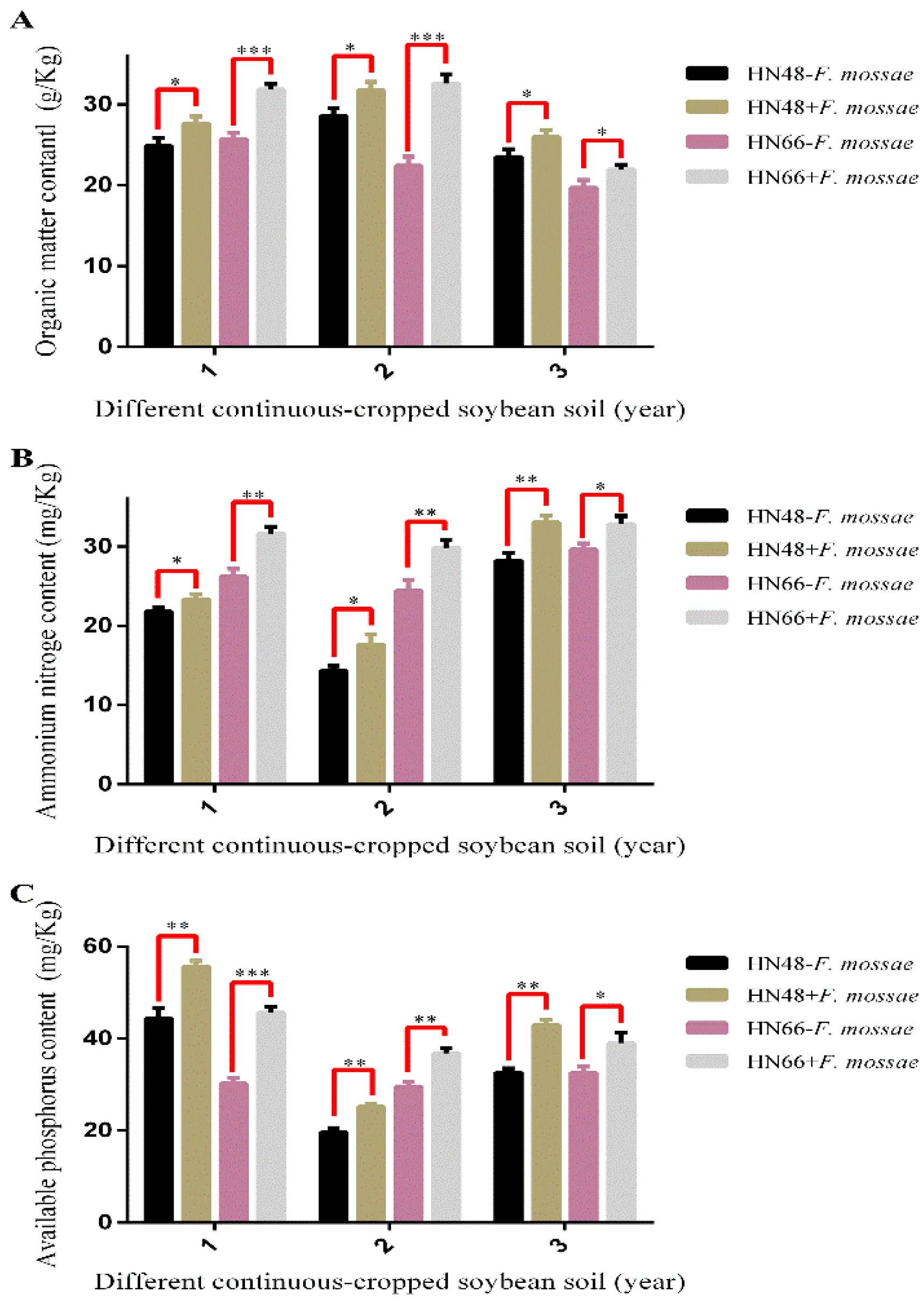
2.4.2. Determination of Rhizosphere Soil Physicochemical Properties in Continuously Cropped Soybean
3. Discussion
3.1. Effect of F. mosseae on The Disease Index of Root Rot in Two Soybean Cultivars over Three Years of Continuous Cropping Under Potted-Experiments
3.2. Effect of F. mosseae on Root Tissue Metabolites and Root Exudates Produced by the Two Cultivars over Three Years of Continuous Cropping under Potted-Experiments
3.3. Effect of F. mosseae on the Biomass and Physicochemical Properties of Rhizosphere Soil in Two Soybean Cultivars over Three Years of Continuous Cropping under Potted-Experiments
4. Materials and Methods
4.1. Soybean Cultivars and Experimental Microbial Agent
4.2. Growth and Maintenance of Soybean Plants
4.3. Detection of Soybean Root Rot Index and AMF Colonization Rate
4.4. Test Sample Collection
4.5. DNA Extraction and Specific Fragment Amplification
4.6. Extraction and Identification of Root Metabolites
4.7. Determination of Continuously Cropped Soybean Biomass and Rhizosphere Soil Physicochemical Properties
4.8. Data Analysis
5. Conclusions
- F. mosseae can reduce the incidence of root rot in continuously cropped soybean.
- F. mosseae inoculation can affect the metabolite profiling in soybean roots.
- Inoculation with F. mosseae increase biomass in continuously cropped soybean.
- F. mosseae inoculation can improve the soybean rhizosphere microenvironment.
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Chen, M.N.; Li, X.; Yu, S.L. Dynamic succession of soil bacterial community during continuous cropping of peanut (Arachis hypogaea L.). PLoS ONE 2014, 9, e101355. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Zhao, Q.Y.; Shen, Q.R. Different continuous cropping spans significantly affect microbial community membership and structure in a vanilla-grown soil as revealed by deep pysosequencing. Microb. Ecol. 2014, 70, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Gil, S.; Meriles, J.; Conforto, C.; Basanta, M. Response of soil microbial communities to different management practices in surface soils of a soybean agroecosystem in Argentina. Eur. J. Soil Biol. 2011, 47, 55–60. [Google Scholar] [CrossRef]
- Liu, J.B.; Xu, Y.L.; Li, C.J.; Wei, W.; Ma, L.L. Effect of Soybean Long Term Continual Cropping on Main Groups of Rhizosphere Fungi. Syst. Sci. Compr. Stud. Agric. 2009, 27, 105–113. [Google Scholar]
- Jeon, C.S.; Kim, G.H.; Koh, Y.J. Root rot of balloon flower (Platycodon grandiflorum) caused by Fusarium solani and Fusarium oxysporum. Plant Pathol. J. 2013, 29, 440–445. [Google Scholar] [CrossRef] [PubMed]
- Scherm, B.; Balmas, V.; Migheli, Q. Fusarium culmorum: Causal agent of foot and root rot and head blight on wheat. Mol. Plant Pathol. 2013, 14, 323–341. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Xu, Y.L.; Li, S.X. Impact of long-term continuous cropping on the Fusarium population in soybean rhizosphere. Ying Yong Sheng Tai Xue Bao 2014, 25, 497–504. [Google Scholar] [PubMed]
- Aliferis, K.A.; Faubert, D.; Jabaji, S. A metabolic profiling strategy for the dissection of plant defense against fungal pathogens. PLoS ONE 2014, 9, e111930. [Google Scholar] [CrossRef] [PubMed]
- Scandiani, M.M.; Luque, A.G.; Spampinato, C.P. Metabolic profiles of soybean roots during early stages of Fusarium tucumaniae infection. J. Exp. Bot. 2015, 66, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.K.; Morang, P.; Dileep-Kumar, B.S. Plant growth-promoting rhizobacterial strain-mediated induced systemic resistance in tea (Camellia sincnsis (L.) O. Kuntze) through defense-related enzymes against brown root rot and charcoal stump rot. Appl. Biochem. Biotechnol. 2014, 174, 506–521. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.T.; Guo, N. Genetic analysis and fine mapping of RpsJS, a novel resistance gene to Phytophthora sojae in soybean [Glycine max (L.) Merr.]. Theor. Appl. Genet. 2014, 127, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Hartman, G.L.; West, E.D.; Herman, T.K. Crops that feed the world 2. Soybean-worldwide production, use, and constraints caused by pathogens and pests. Food Secur. 2011, 3, 5–17. [Google Scholar] [CrossRef]
- Favre, P.; Bapaume, L.; Reinhardt, D. A novel bioinformatics pipeline to discover genes related to arbuscular mycorrhizal symbiosis based on their evolutionary conservation pattern among higher plants. BMC Plant Biol. 2014, 14, 333. [Google Scholar] [CrossRef] [PubMed]
- Leifheit, E.F.; Veresoglou, S.D.; Rillig, M.C. Multiple factors influence the role of arbuscular mycorrhizal fungi in soil aggregation-a meta-analysis. Plant Soil 2014, 374, 523–537. [Google Scholar] [CrossRef]
- Walder, F.; Niemann, H.; Natarajan, M.; Lehmann, M.F.; Boller, T.; Wiemken, A. Mycorrhizal networks: Common goods of plants shared under unequal terms of trade. Plant Physiol. 2012, 159, 789–797. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, A.; Kathryn-Barto, E.; Rillig, M.C. Mycorrhizal responsiveness trends in annual crop plants and their wild relatives-a meta-analysis on studies from 1981 to 2010. Plant Soil 2012, 355, 231–250. [Google Scholar] [CrossRef]
- Zhang, H.S.; Li, G.P.; Pan, S.M. Castor bean growth and rhizosphere soil property response to different proportions of arbuscular mycorrhizal and phosphate-solubilizing fungi. Ecol. Res. 2014, 29, 181–190. [Google Scholar] [CrossRef]
- Huang, Y.M.; Srivastava, A.K.; Wu, Q.S. Mycorrhizal-induced calmodulin mediated changes in antioxidant enzymes and growth response of drought-stressed trifoliate orange. Front. Microbiol. 2014, 5, 682. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.G.; Zhu, Z.; Shen, Q.R. Trichoderma harzianum T-E5 significantly affects cucumber root exudates and fungal community in the cucumber rhizosphere. Appl. Soil Ecol. 2013, 72, 41–48. [Google Scholar] [CrossRef]
- Ortiz, N.; Armada, E.; Azcon, R. Contribution of arbuscular mycorrhizal fungi and/or bacteria to enhancing plant drought tolerance under natural soil conditions: Effectiveness of autochthonous or allochthonous strains. J. Plant Physiol. 2011, 174, 87–96. [Google Scholar] [CrossRef] [PubMed]
- El-Faiz, A.; Duponnois, R.; Hafidi, M. Effect of different amendments on growing of Canna indica L. inoculated with AMF on mining substrate. Int. J. Phytoremediat. 2015, 17, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Bellot, M.J.; Ortuno, M.F.; Sanchez-Blanco, M.J. Protective effects of Glomus iranicum var. tenuihypharum on soil and Viburnum tinus plants irrigated with treated wastewater under field conditions. Mycorrhiza 2015, 25, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Dhawi, F.; Datta, R.; Ramakrishna, W. Mycorrhiza and heavy metal resistant bacteria enhance growth, nutrient uptake and alter metabolic profile of sorghum grown in marginal soil. Chemosphere 2016, 157, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Son, S.Y.; Kim, N.K.; Lee, S.M. Metabolite fingerprinting, pathway analyses, and bioactivity correlations for plant species belonging to the Cornaceae, Fabaceae, and Rosaceae families. Plant Cell Rep. 2016. [Google Scholar] [CrossRef] [PubMed]
- Schubler, A.; Walker, C. The Glomeromycota. A Species List with New Families and New Genera. In Libraries at The Royal Botanic Garden Edinburgh; The Royal Botanic Garden Kew, Botanische Staatssammlung Munich: Munich, Germany; Oregon State University: Portland, OR, USA, 2010. [Google Scholar]
- Luan, F.G.; Zhang, L.L.; Zhang, H.Y. Analysis of microbial diversity and niche in rhizosphere soil of healthy and diseased cotton at the flowering stage in southern Xinjiang. Genet. Mol. Res. 2015, 14, 1602–1611. [Google Scholar] [CrossRef] [PubMed]
- Noel, G.R.; Wax, L.M. Population dynamics of Heterodera glycines in conventional tillage and no-tillage soybean/corn cropping systems. J. Nematol. 2003, 35, 104–109. [Google Scholar] [PubMed]
- Zhang, X.L.; Li, X.; Zhang, H.W. Ecological risk of long-term chlorimuron-ethyl applicationto soil microbial community: An in situ investigationin a continuously cropped soybean field in Northeast China. Environ. Sci. Pollut. Res. Int. 2011, 18, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Cui, J.Q.; Cai, B.Y. Analysis of the community compositions of rhizosphere fungi in soybeans continuous cropping fields. Microbiol. Res. 2015, 180, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.Q.; Wang, Y.; Cai, B.Y. Analyses of the community compositions of root rot pathogenic fungi in the soybean rhizosphere soil. Chil. J. Agric. Res. 2016, 76, 1–7. [Google Scholar] [CrossRef]
- Jie, W.G.; Liu, X.R.; Cai, B.Y. Diversity of rhizosphere soil arbuscular mycorrhizal fungi in various soybean cultivars under different continuous cropping regimes. PLoS ONE 2013, 8, e72898. [Google Scholar] [CrossRef] [PubMed]
- Jie, W.G.; Bai, L.; Cai, B.Y. Analysis of interspecific relationships between Funeliformis mosseae and Fusarium oxysporum in the continuous cropping of soybean rhizosphere soil during the branching period. Biocontrol Sci. Technol. 2015, 25, 1036–1051. [Google Scholar] [CrossRef]
- Qian, L.; Yu, W.J.; Cai, B.Y. Funnelifomis mosseae affects the root rot pathogen Fusarium oxysporum in soybeans. Acta Agric. Scand. B Soil Plant Sci. 2015, 64, 321–327. [Google Scholar]
- Krüger, G.J.; De-Villiers, A.J.; Strasser, R.J. Inhibition of photosystem II activities in soybean (Glycine max) genotypes differing in chilling sensitivity. S. Afr. J. Bot. 2014, 95, 85–96. [Google Scholar] [CrossRef]
- Lingo, N.; Ma, J.H.; Shen, Q.R. Identification and role of organic acids in watermelon root exudates for recruiting Paenibacillus polymyxa SQR-21 in the rhizosphere. Eur. J. Soil Biol. 2011, 47, 374–379. [Google Scholar] [CrossRef]
- Martin, B.C.; George, S.J.; Tibbet, M. The role of root exuded low molecular weight organic anions in facilitating petroleum hydrocarbon degradation: Current knowledge and future directions. Sci. Total Environ. 2014, 47, 2642–2653. [Google Scholar] [CrossRef] [PubMed]
- Wu, L. Identify the Substances in Root Exudates of Soybean and Study on Allelopathy. Master’s Thesis, Northeast Agricultural University, Harbin, China, 2010. [Google Scholar]
- Yang, X.; Zhang, L.H.; Dong, J.G. The extraction, isolatioin and identification of exudate from the root of Flaveria bidentis. J. Integr. Agric. 2014, 13, 105–114. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Dong, W.Y.; Sun, X.M. Responses of absolute and specific soil enzyme activities to long term additions of organic and mineral fertilizer. Sci. Total Environ. 2015, 536, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.; Liu, P.; Xu, G.D. The effects of rapeseed root exudates on the forms of aluminum in aluminum stressed rhizosphere soil. Crop Prot. 2011, 30, 631–636. [Google Scholar] [CrossRef]
- Mcgonigle, T.P.; Millers, M.H.; Evans, D.G. A new method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fungi. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef]
- May, L.A.; Smiley, B.; Schmidt, M.G. Comparative denaturing gradient gel electrophoresis analysis of fungal communities associated with whole plant corn silage. Can. J. Microbiol. 2011, 47, 829–841. [Google Scholar] [CrossRef]
- Van-Tuinen, D.; Zhao, B.; Gianinazzi-Pearson, V. PCR in studies of AM fungi: From primers to application. In Mycorrhiza Manual; Varma, A.K., Ed.; Springer: Heidelberg, Germany, 1988; pp. 387–399. [Google Scholar]
- Xia, J.; Mandal, R.; Sinelnikov, I.V.; Broadhurst, D.; Wishart, D.S. Metabo Analyst 2.0 a comprehensive server for metabolomic data analysis. Nucleic Acids Res. 2012, 40, 127–133. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Retention Time (min) | Compounds | Metabolic Level | VIP Value | p Value |
|---|---|---|---|---|
| 11.049 | Benzoic acid, 2-fluoro-, ethyl ester | Down | 2.2149 | 0.0096 |
| 42.635 | Bis(2-ethylhexyl) phthalate | Down | 2.1669 | 0.0104 |
| 43.42 | Hexacosane | Down | 2.1321 | 0.0165 |
| 33.973 | Hexadecanoic acid | Up | 2.1125 | 0.0021 |
| 21.62 | Dodecane | Up | 2.0025 | 0.0214 |
| 43.13 | Tricosane, 2-methyl- | Down | 1.9423 | 0.0475 |
| 30.164 | 1,2-Benzenedic carboxylic acid, bis(2-methylpropyl) ester | Down | 1.916 | 0.0235 |
| 31.87 | n-Pentadecanoic acid | Down | 1.9012 | 0.0032 |
| Retention Time (min) | Compounds | Metabolic Level | VIP Value | p Value |
|---|---|---|---|---|
| 12.317 | Propanoic acid, 2-(hydroxyl)- | Up | 1.9188 | 0.0092 |
| 46.432 | Tetracosane | Down | 1.7592 | 0.0162 |
| 42.635 | Bis(2-ethylhexyl) phthalate | Down | 1.7348 | 0.0208 |
| 44.384 | Heptadecane, 9-hexyl- | Down | 1.7081 | 0.0024 |
| 33.943 | Hexadecanoic acid | Up | 1.6488 | 0.0196 |
| 31.803 | Benzenepropanoic acid,3,5-bis(1,1-dimethylethyl)-4-hydroxy-, methyl ester | Down | 1.6223 | 0.0157 |
| 37.588 | cis-9-Hexadecenoic acid | Down | 1.5762 | 0.0184 |
| Retention Time (min) | Compounds | Metabolic Level | VIP Value | p Value |
|---|---|---|---|---|
| 12.218 | Benzene, (1-methyl-1-butenyl)- | Down | 2.1285 | 0.0452 |
| 39.282 | Heptadecane, 2-methyl | Up | 2.0193 | 0.0314 |
| 42.476 | Bis(2-ethylhexyl) phthalate | Down | 1.9912 | 0.0299 |
| 35.533 | Heneicosane | Up | 1.9749 | 0.0189 |
| 20.077 | Phenol, 2,4-bis(1,1-dimethylethyl) | Down | 1.8462 | 0.0329 |
| 19.953 | Sulfurous acid, 2-propyl tetradecyl ester | Down | 1.8423 | 0.0415 |
| 18.119 | Naphthalene, 1,3-dimethyl- | Down | 1.8116 | 0.0235 |
| 26.006 | Heptadecane | Up | 1.8056 | 0.0032 |
| Retention Time (min) | Compounds | Metabolic Level | VIP Value | p Value |
|---|---|---|---|---|
| 42.476 | Bis(2-ethylhexyl) phthalate | Down | 2.1566 | 0.0252 |
| 34.814 | Octacosane | Down | 2.1169 | 0.0424 |
| 36.337 | Octadecane, 2-methyl- | Up | 1.9912 | 0.0299 |
| 31.742 | Benzene, (1-methyl-1-butenyl)- | Down | 1.9749 | 0.0189 |
| 32.181 | Dibutyl phthalate | Down | 1.9156 | 0.0154 |
| 44.694 | Tricosane | Up | 1.8641 | 0.0077 |
| 46.108 | Tetracosane | Up | 1.8544 | 0.0044 |
| 20.077 | Phenol, 2,4-bis(1,1-dimethylethyl) | Down | 1.8356 | 0.0012 |
| 38.422 | Octadecane | Up | 1.638 | 0.0216 |
| 26.03 | Heptadecane | Down | 1.638 | 0.0216 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, J.-Q.; Sun, H.-B.; Sun, M.-B.; Liang, R.-T.; Jie, W.-G.; Cai, B.-Y. Effects of Funneliformis mosseae on Root Metabolites and Rhizosphere Soil Properties to Continuously-Cropped Soybean in the Potted-Experiments. Int. J. Mol. Sci. 2018, 19, 2160. https://doi.org/10.3390/ijms19082160
Cui J-Q, Sun H-B, Sun M-B, Liang R-T, Jie W-G, Cai B-Y. Effects of Funneliformis mosseae on Root Metabolites and Rhizosphere Soil Properties to Continuously-Cropped Soybean in the Potted-Experiments. International Journal of Molecular Sciences. 2018; 19(8):2160. https://doi.org/10.3390/ijms19082160
Chicago/Turabian StyleCui, Jia-Qi, Hai-Bing Sun, Ming-Bo Sun, Rui-Ting Liang, Wei-Guang Jie, and Bai-Yan Cai. 2018. "Effects of Funneliformis mosseae on Root Metabolites and Rhizosphere Soil Properties to Continuously-Cropped Soybean in the Potted-Experiments" International Journal of Molecular Sciences 19, no. 8: 2160. https://doi.org/10.3390/ijms19082160
APA StyleCui, J.-Q., Sun, H.-B., Sun, M.-B., Liang, R.-T., Jie, W.-G., & Cai, B.-Y. (2018). Effects of Funneliformis mosseae on Root Metabolites and Rhizosphere Soil Properties to Continuously-Cropped Soybean in the Potted-Experiments. International Journal of Molecular Sciences, 19(8), 2160. https://doi.org/10.3390/ijms19082160