Bornyl cis-4-Hydroxycinnamate Suppresses Cell Metastasis of Melanoma through FAK/PI3K/Akt/mTOR and MAPK Signaling Pathways and Inhibition of the Epithelial-to-Mesenchymal Transition

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
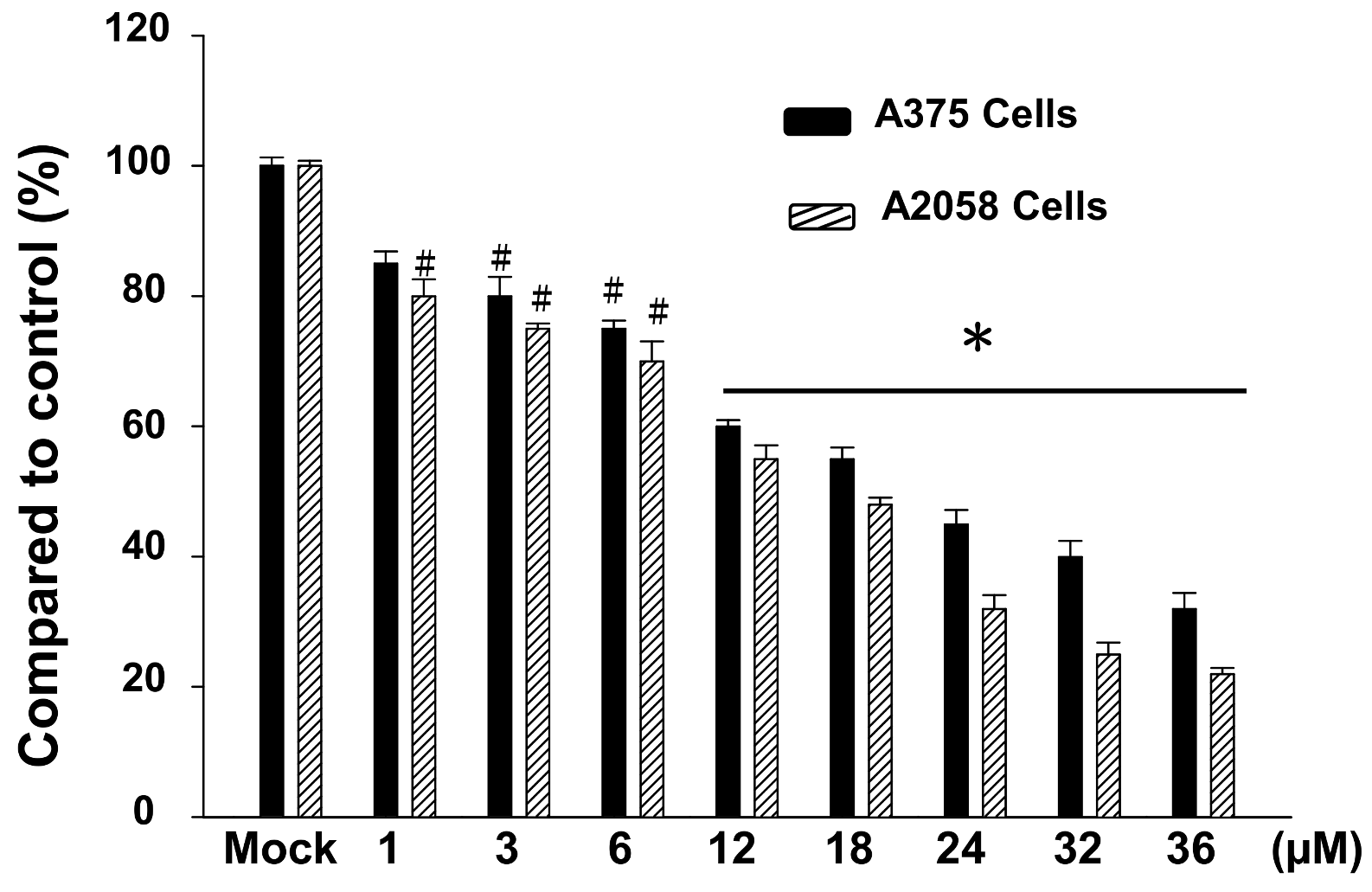
2.1. The Cytotoxic Effects of Bornyl cis-4-Hydroxycinnamate on A2058 and A375 Cells
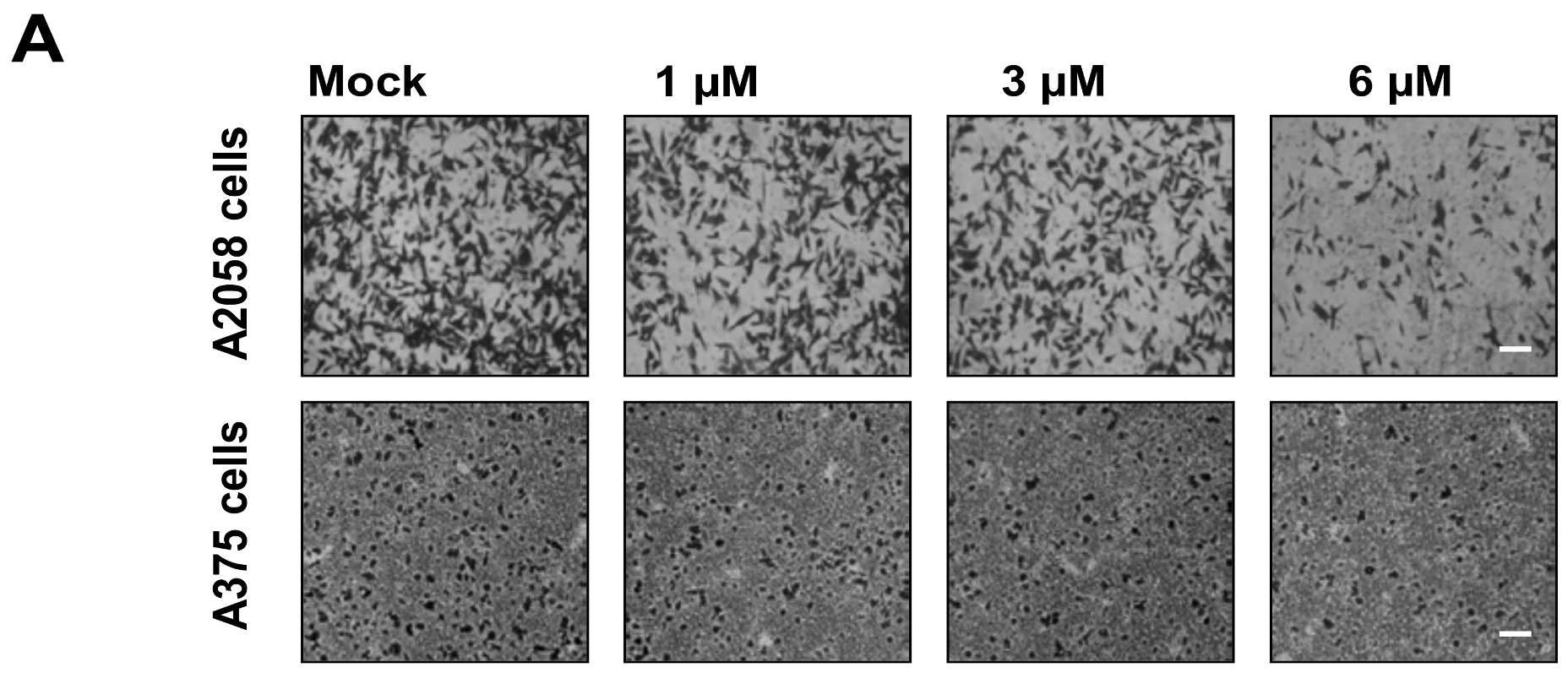
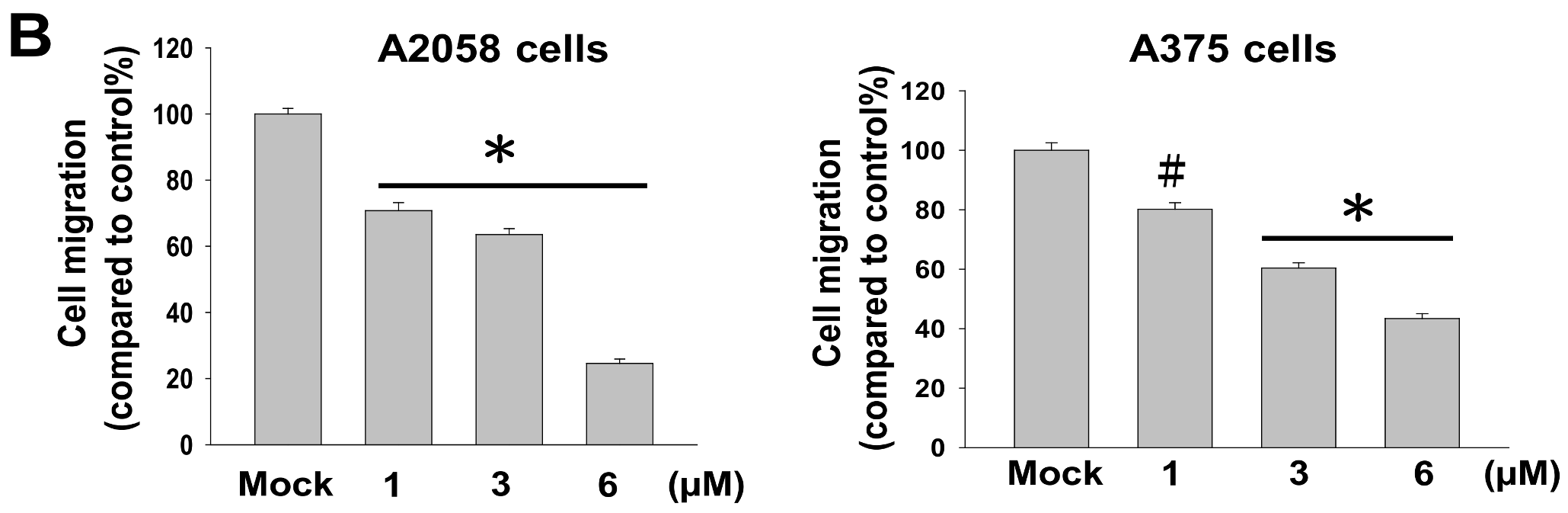
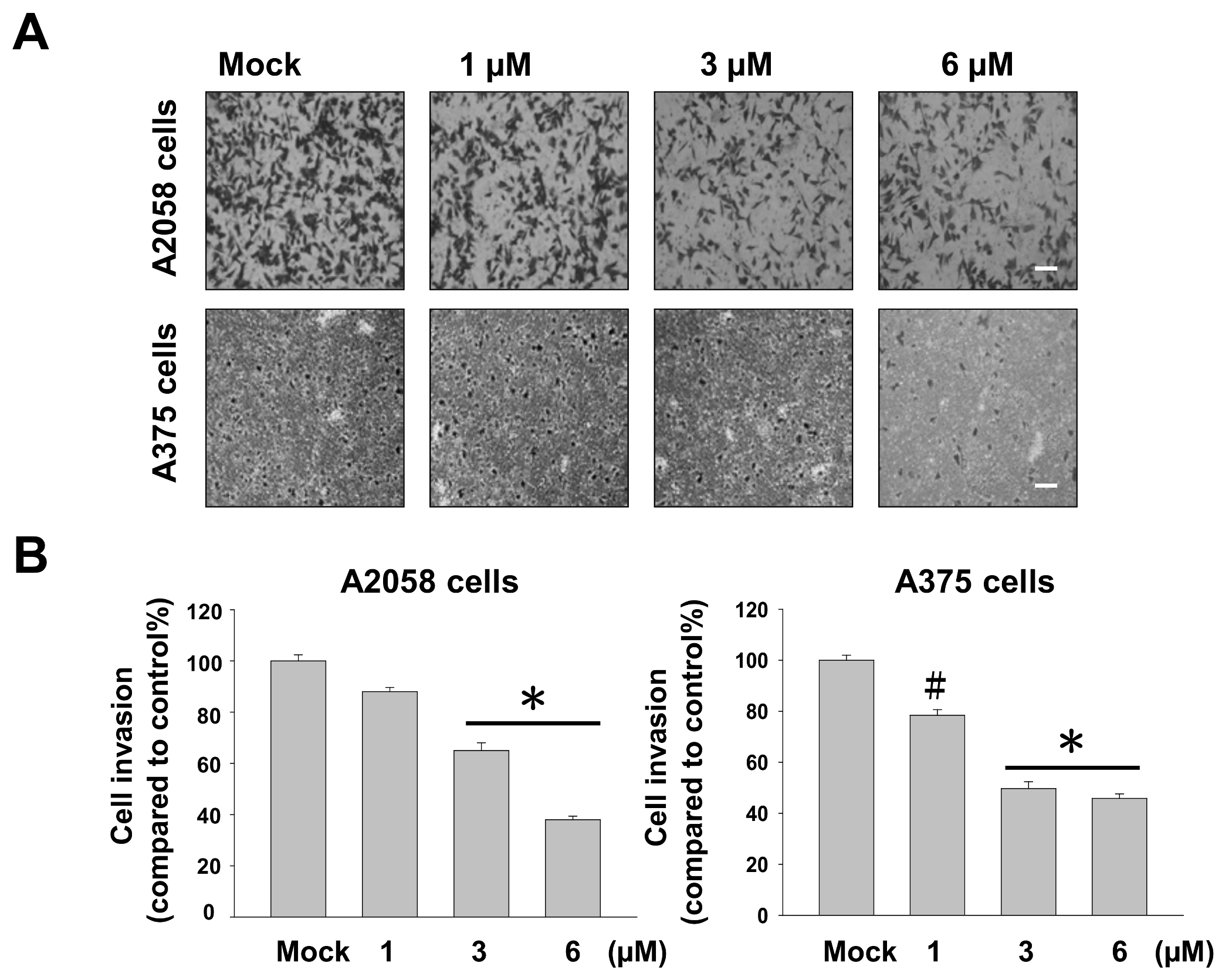
2.2. Bornyl cis-4-Hydroxycinnamate Inhibited Migration and Invasion of A2058 and A375 Cells
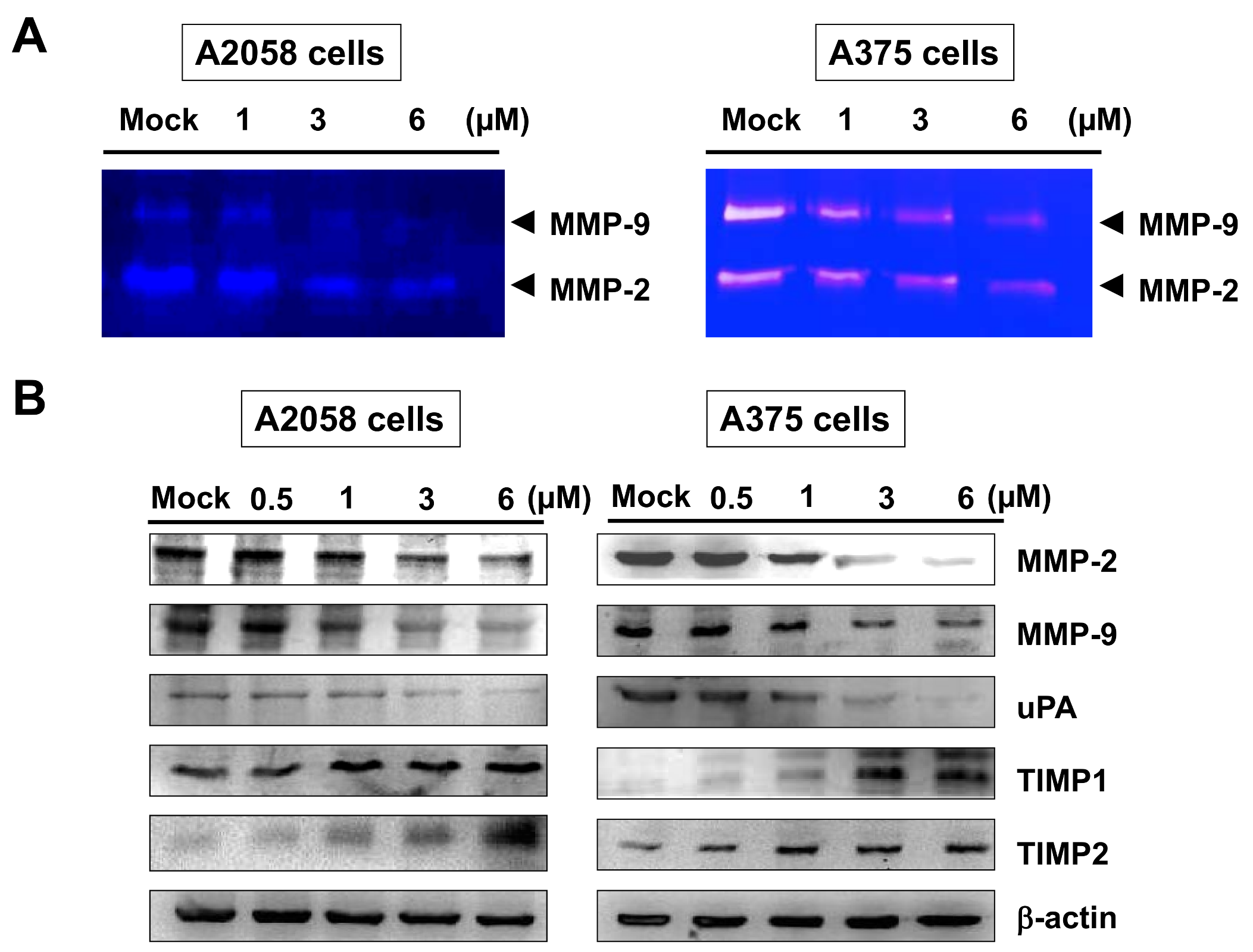
2.3. Bornyl cis-4-Hydroxycinnamate Reduced the MMP-2/-9 Activities and Regulated the Expressions of MMP-2, MMP-9, uPA, TIMP-1, and TIMP-2 Proteins in A2058 and A375 Cells
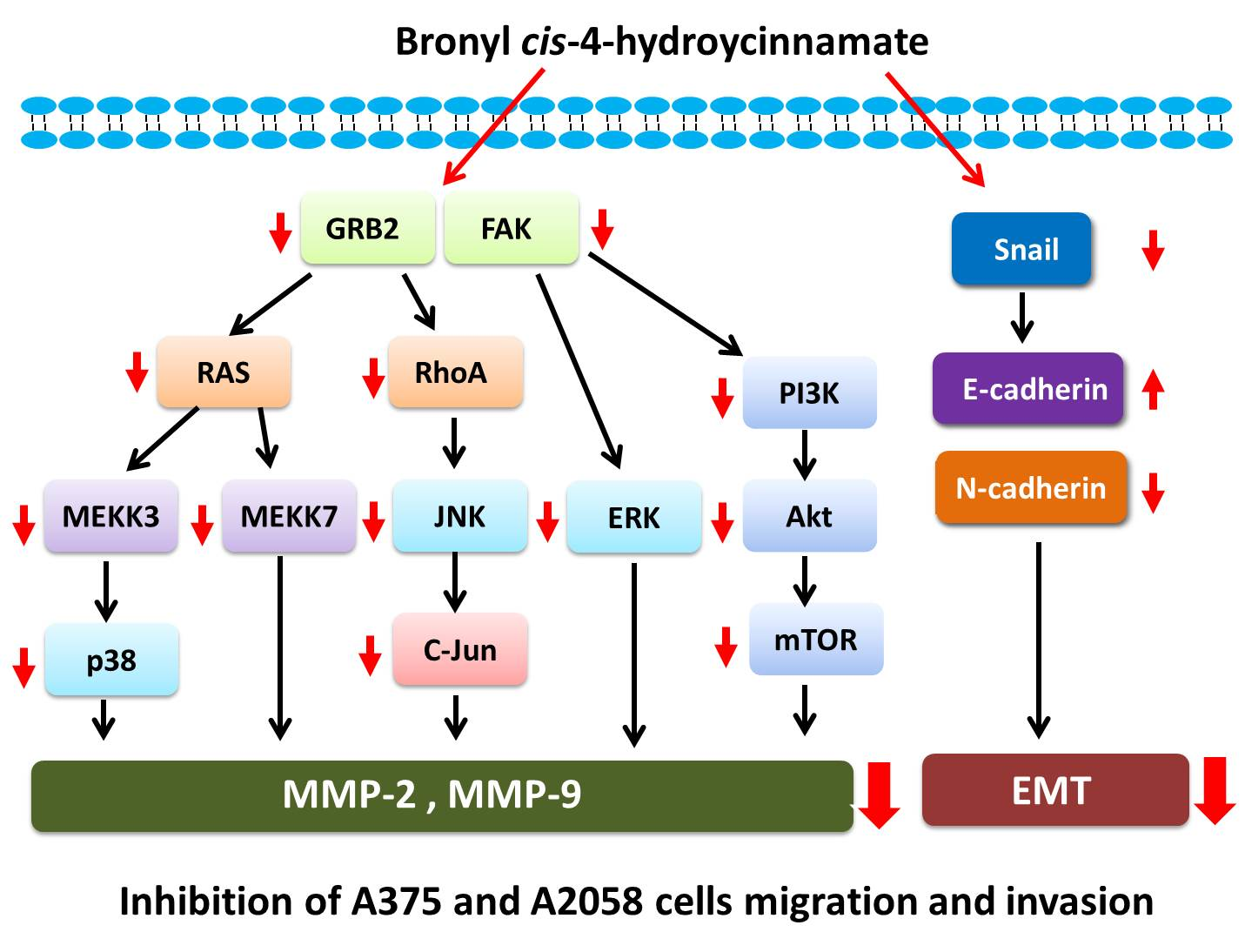
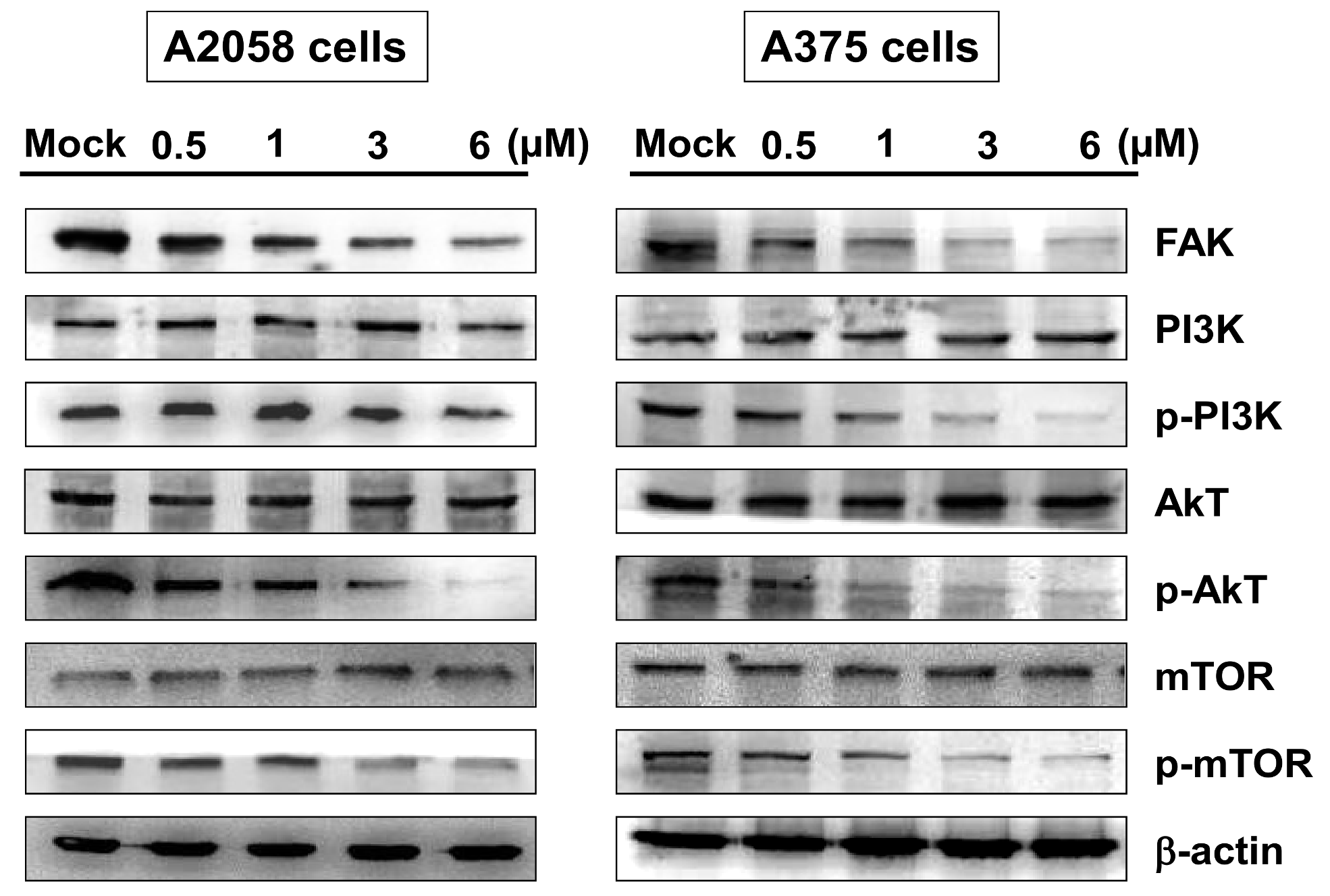
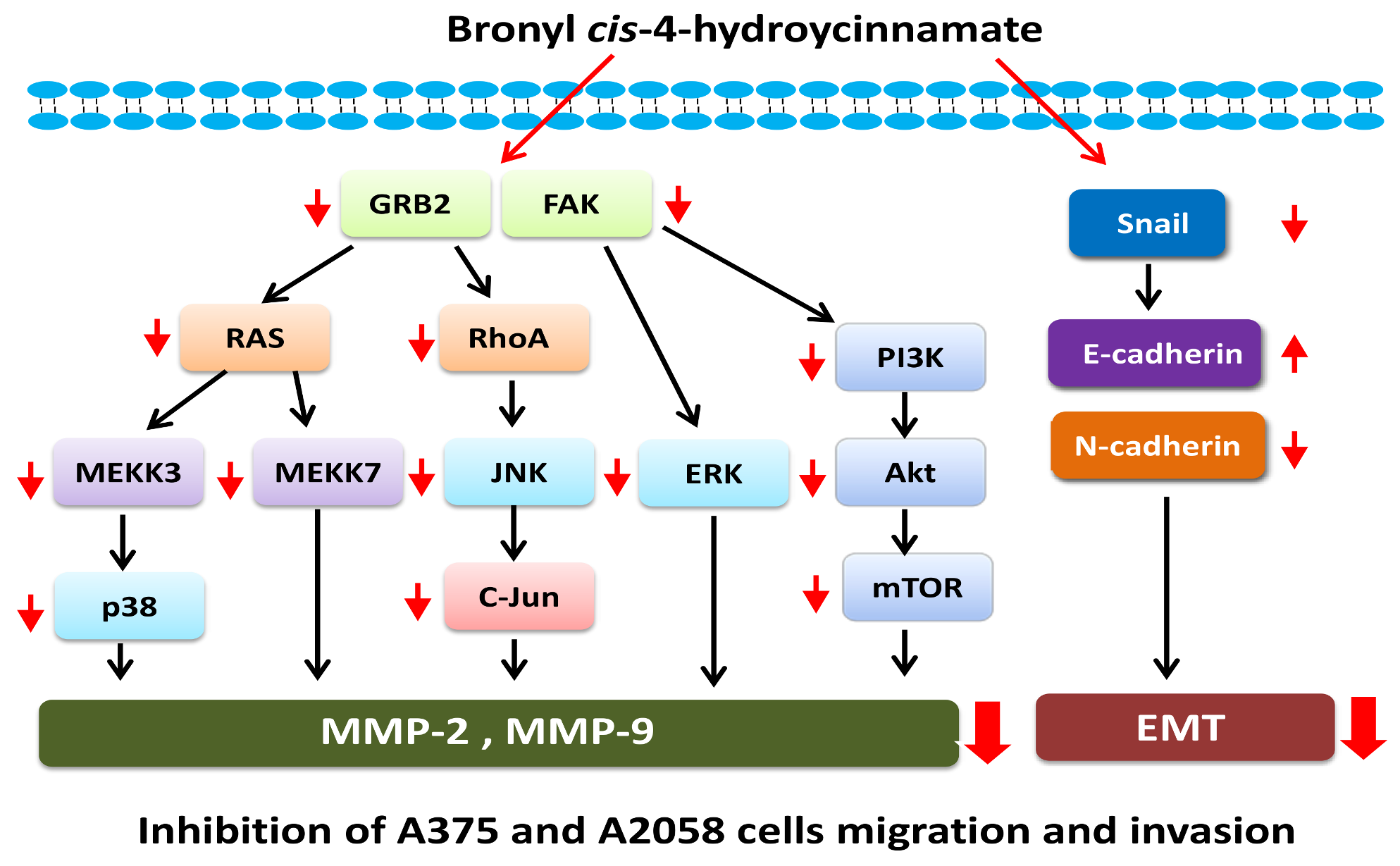
2.4. Bornyl cis-4-Hydroxycinnamate Inhibited FAK/PI3K/Akt/mTOR Signaling Pathway-Associated Proteins
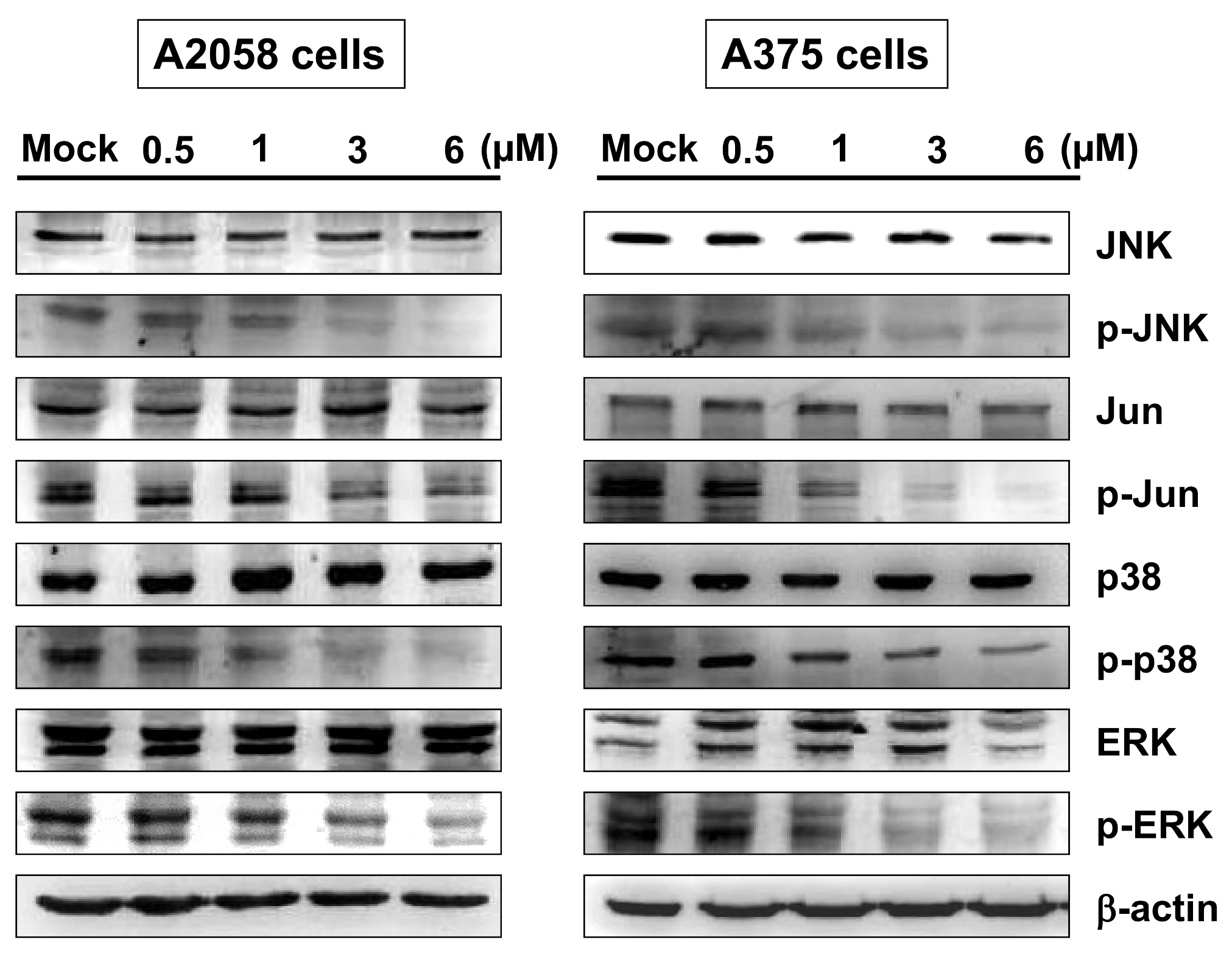
2.5. Bornyl cis-4-Hydroxycinnamate Inhibited MAPK Signaling Pathway-Related Molecules
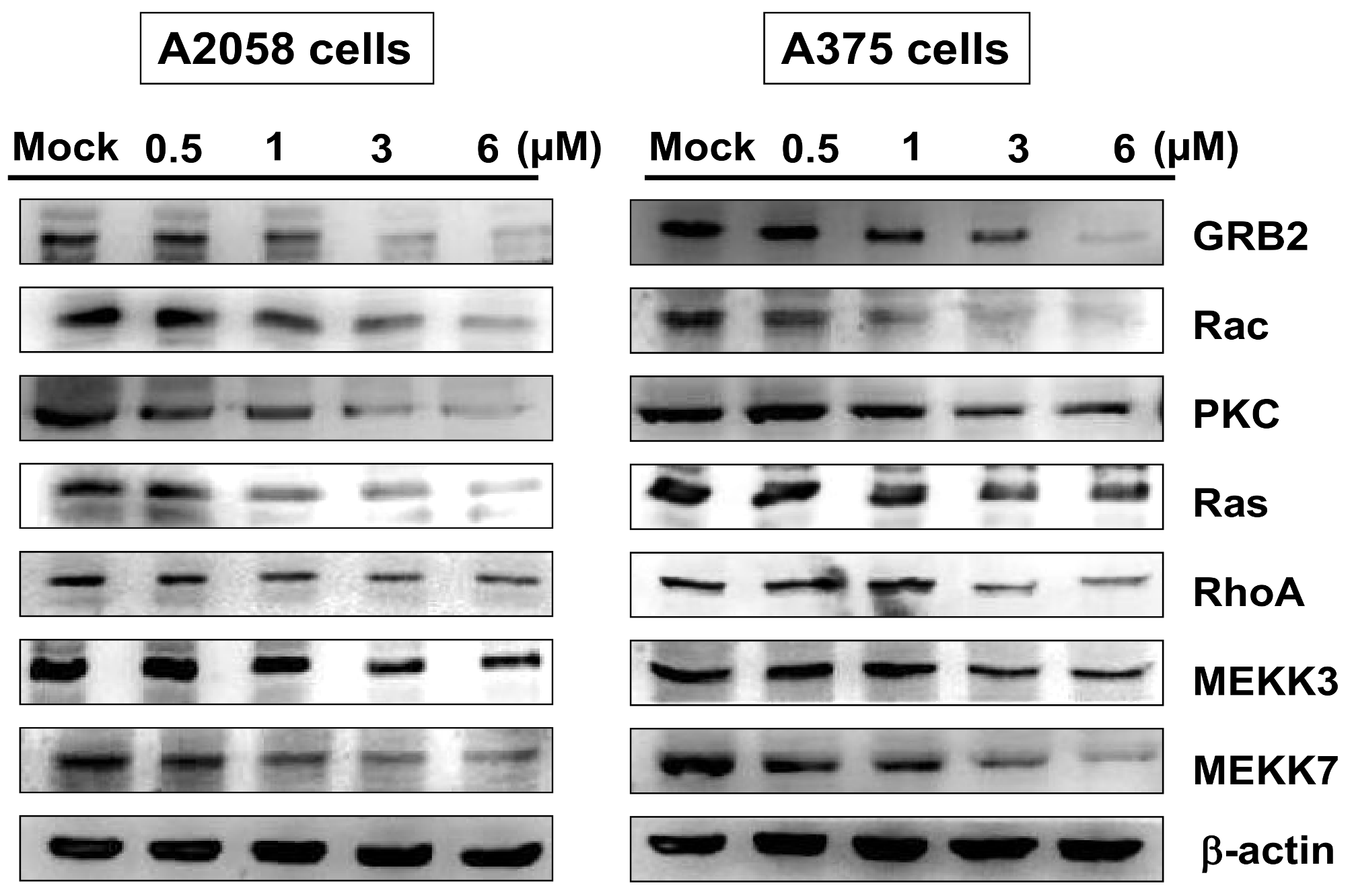
2.6. Bornyl cis-4-Hydroxycinnamate Inhibited the GRB2 Signaling Pathway
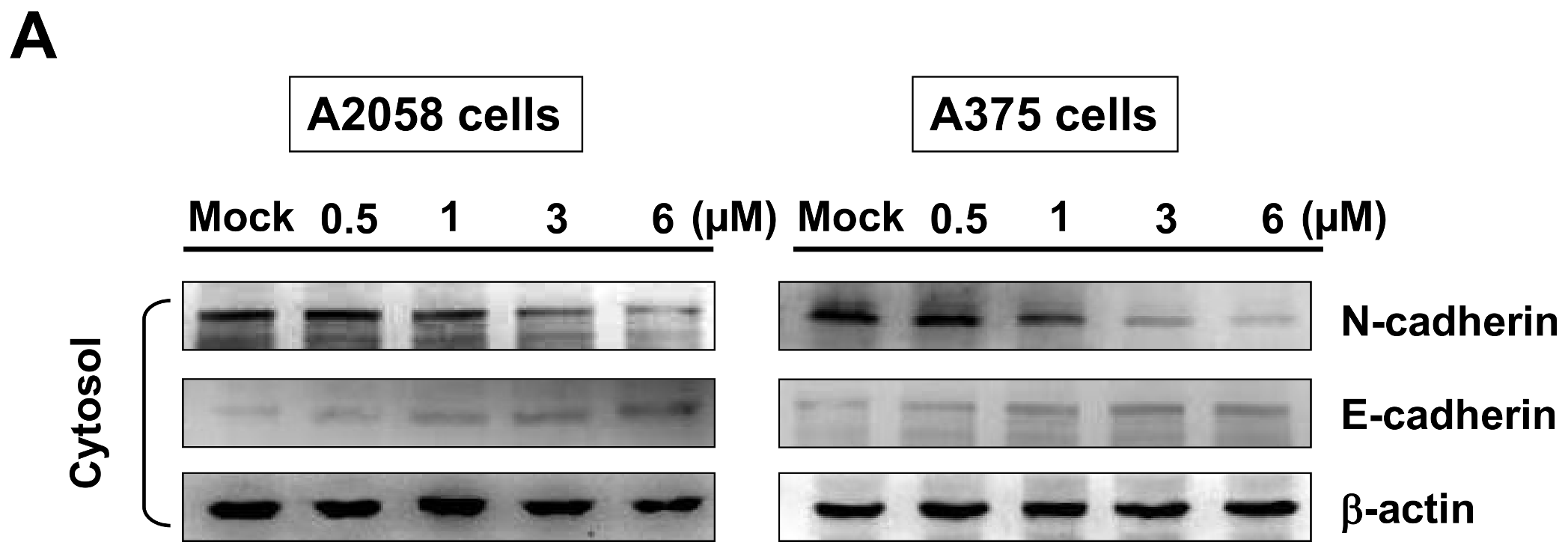
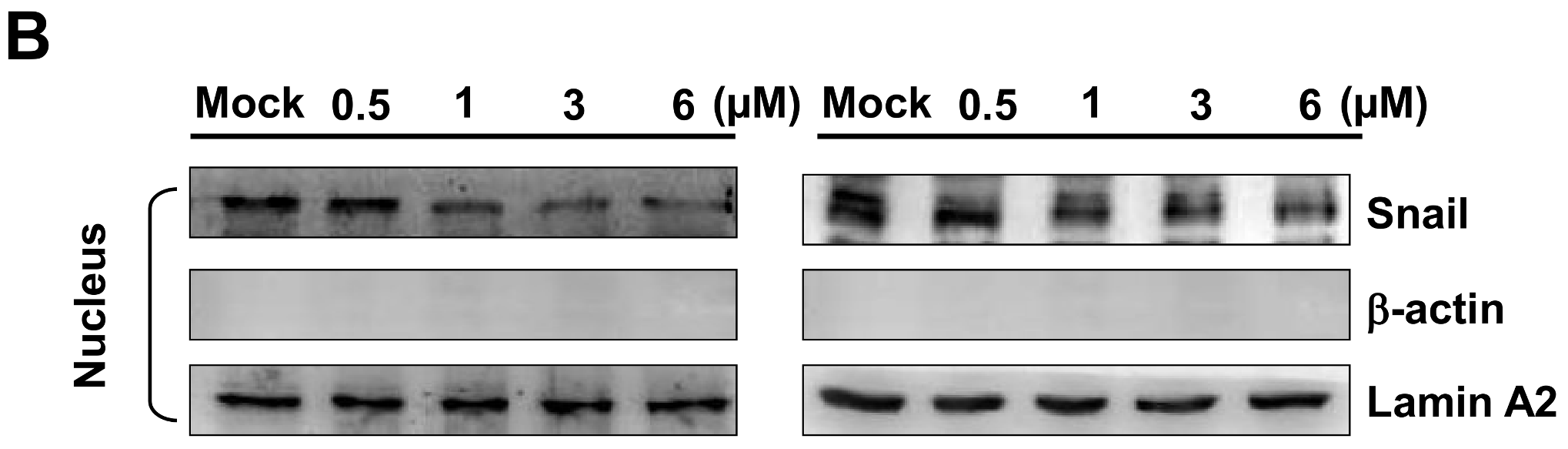
2.7. Bornyl cis-4-Hydroxycinnamate Inhibited Epithelial to Mesenchymal Transition (EMT)
3. Discussion
4. Methods
4.1. Materials and Chemical Reagents
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. Cell Migration Assay
4.5. Cell Invasion Assay
4.6. Gelatin Zymography Assay
4.7. Western Blot Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Parkin, D.M.; Bray, F.; Ferlay, J.; Pisani, P. Global cancer statistics, 2002. CA Cancer J. Clin. 2005, 55, 74–108. [Google Scholar] [CrossRef] [PubMed]
- Chang, J. Cutaneous melanoma: Taiwan experience and literature review. Chang Gung Med. J. 2010, 33, 602–612. [Google Scholar] [PubMed]
- Essner, R. Surgical treatment of malignant melanoma. Surg. Clin. 2003, 83, 109–156. [Google Scholar] [CrossRef]
- Bleehen, N.; Newlands, E.; Lee, S.M.; Thatcher, N.; Selby, P.; Calvert, A.; Rustin, G.; Brampton, M.; Stevens, M. Cancer research campaign phase II trial of temozolomide in metastatic melanoma. J. Clin. Oncol. 1995, 13, 910–913. [Google Scholar] [CrossRef] [PubMed]
- Treisman, J.; Garlie, N. Systemic therapy for cutaneous melanoma. Clin. Plast Surg. 2010, 37, 127–146. [Google Scholar] [CrossRef] [PubMed]
- Acquavella, N.; Kluger, H.; Rhee, J.; Farber, L.; Tara, H.; Ariyan, S.; Narayan, D.; Kelly, W.; Sznol, M. Toxicity and activity of a twice daily high-dose bolus interleukin 2 regimen in patients with metastatic melanoma and metastatic renal cell cancer. J. Immunother. 2008, 31, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, N.; Haluska, F.G. Molecular pathogenesis of cutaneous melanocytic neoplasms. Annu. Rev. Pathol.-Mech. 2009, 4, 551–579. [Google Scholar] [CrossRef] [PubMed]
- Fuglede, N.; Brinck-Claussen, U.; Deltour, I.; Boesen, E.; Dalton, S.; Johansen, C. Incidence of cutaneous malignant melanoma in Denmark, 1978–2007. Brit. J. Dermatol. 2011, 165, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Sasse, A.D.; Sasse, E.C.; Clark, L.; Ulloa, L.; Clark, O. Chemoimmunotherapy versus chemotherapy for metastatic malignant melanoma. Cochrane Database Syst. Rev. 2007, 24, CD005413. [Google Scholar] [CrossRef] [PubMed]
- Tawbi, H.A.; Buch, S.C. Chemotherapy resistance abrogation in metastatic melanoma. Clin. Adv. Hematol. Oncol. 2010, 8, 259–266. [Google Scholar] [PubMed]
- Khan, J.A.; Kumar, N. Evaluation of antibacterial properties of extracts of Piper betel leaf. J. Pharmaceut. Biomed. Sci. 2011, 11, 1–3. [Google Scholar]
- Sarkar, A.; Sen, R.; Saha, P.; Ganguly, S.; Mandal, G.; Chatterjee, M. An ethanolic extract of leaves of Piper betle (PAAN) linn mediates its antileishmanial activity via apoptosis. Parasitol. Res. 2008, 102, 1249–1255. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Shakya, S.; Soni, V.K.; Dangi, A.; Kumar, N.; Bhattacharya, S.-M. The n-hexane and chloroform fractions of Piper betle L. Trigger different arms of immune responses in BALB/c mice and exhibit antifilarial activity against human lymphatic filarid brugia malayi. Int. Immunopharmacol. 2009, 9, 716–728. [Google Scholar] [CrossRef] [PubMed]
- Himratul-Aznita, W.; Mohd-Al-Faisal, N.; Fathilah, A. Determination of the percentage inhibition of diameter growth (PIDG) of Piper betle crude aqueous extract against oral candida species. J. Med. Plants Res. 2011, 5, 878–884. [Google Scholar]
- Al-Adhroey, A.H.; Nor, Z.M.; Al-Mekhlafi, H.M.; Amran, A.A.; Mahmud, R. Antimalarial activity of methanolic leaf extract of Piper betle L. Molecules 2011, 16, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Wardhana, A.H.; Kumarasinghe, S.P.W.; Arawwawala, L.; Arambewela, L.S. Larvicidal efficacy of essential oil of betel leaf (Piper betle) on the larvae of the old world screwworm fly, chrysomya bezziana in vitro. Indian J. Dermatol. 2007, 52, 43–47. [Google Scholar]
- Fathilah, A.; Sujata, R.; Norhanom, A.; Adenan, M. Antiproliferative activity of aqueous extract of Piper betle L. And psidium guajava L. on KB and HeLa cell lines. J. Med. Plants Res. 2010, 4, 987–990. [Google Scholar]
- Rekha, V.; Kollipara, M.; Gupta, B.S.; Bharath, Y.; Pulicherla, K.K. A review on Piper betle L.: Natureâ s promising medicinal reservoir. Am. J. Ethnomed. 2014, 1, 276–289. [Google Scholar]
- Bhuvaneswari, S.; Sripriya, N.; Deepa, S.; Prakash, N.U. Studies on antioxidant activities of six cultivars of Piper betle linn. Int. J. Pharm. Pharmceut. Sci. 2014, 6, 270–273. [Google Scholar]
- Abrahim, N.N.; Kanthimathi, M.; Abdul-Aziz, A. Piper betle shows antioxidant activities, inhibits MCF-7 cell proliferation and increases activities of catalase and superoxide dismutase. BMC Complement. Altern. Med. 2012, 12, 220. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Misra, P.; Dube, A.; Bhattacharya, S.; Dikshit, M.; Ranade, S. Piper betle Linn. A maligned pan-asiatic plant with an array of pharmacological activities and prospects for drug discovery. Curr. Sci. 2010, 99, 922–932. [Google Scholar]
- Rai, M.P.; Thilakchand, K.R.; Palatty, P.L.; Rao, P.; Rao, S.; Bhat, H.P.; Baliga, M.S. Piper betel linn (betel vine), the maligned southeast asian medicinal plant possesses cancer preventive effects: Time to reconsider the wronged opinion. Asian Pac. J. Cancer Prev. 2011, 12, 2149–2156. [Google Scholar] [PubMed]
- Wu, P.-F.; Tseng, H.-C.; Chyau, C.-C.; Chen, J.-H.; Chou, F.-P. Piper betle leaf extracts induced human hepatocellular carcinoma Hep3B cell death via mapks regulating the p73 pathway in vitro and in vivo. Food Funct. 2014, 5, 3320–3328. [Google Scholar] [CrossRef] [PubMed]
- Young, S.-C.; Wang, C.-J.; Hsu, J.-D.; Hsu, J.-L.; Chou, F.-P. Increased sensitivity of Hep G2 cells toward the cytotoxicity of cisplatin by the treatment of Piper betel leaf extract. Arch. Toxicol. 2006, 80, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.-Y.; Wu, Y.-J.; Chang, C.-I.; Chiu, C.-C.; Wu, M.-L. The effect of bornyl cis-4-hydroxycinnamate on melanoma cell apoptosis is associated with mitochondrial dysfunction and endoplasmic reticulum stress. Int. J. Mol. Sci. 2018, 19, 1370. [Google Scholar] [CrossRef] [PubMed]
- Giubellino, A.; Burke, T.R.; Bottaro, D.P. GRB2 signaling in cell motility and cancer. Expert Opin. Ther. Targets 2008, 12, 1021–1033. [Google Scholar] [CrossRef] [PubMed]
- ATCC. A-375 [A375] (ATCC® CRL-1619™). Available online: www.atcc.org/Products/All/CRL-1619.aspx (accessed on 19 December 2017).
- ATCC. A2058 (ATCC® CRL-11147™). Available online: https://www.atcc.org/Products/All/CRL-11147.aspx (accessed on 19 December 2017).
- Jiao, Y.; Feng, X.; Zhan, Y.; Wang, R.; Zheng, S.; Liu, W.; Zeng, X. Matrix metalloproteinase-2 promotes αvβ3 integrin-mediated adhesion and migration of human melanoma cells by cleaving fibronectin. PLoS ONE 2012, 7, e41591. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, U.B.; Westphal, J.R.; van Muijen, G.N.; Ruiter, D.J. Matrix metalloproteinases in human melanoma. J. Investig. Dermatol. 2000, 115, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Redondo, P.; Lloret, P.; Idoate, M.; Inoges, S. Expression and serum levels of MMP-2 and MMP-9 during human melanoma progression. Clin. Exp. Dermatol. 2005, 30, 541–545. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Liu, L.; Liu, L.; Geng, J.; Zhou, Y.; Chen, L. β-elemene inhibits the metastasis of B16F10 melanoma cells by downregulation of the expression of uPA, uPAR, MMP-2, and MMP-9. Melanoma Res. 2014, 24, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Alpízar-Alpízar, W.; Christensen, I.J.; Santoni-Rugiu, E.; Skarstein, A.; Ovrebo, K.; Illemann, M.; Laerum, O.D. Urokinase plasminogen activator receptor on invasive cancer cells: A prognostic factor in distal gastric adenocarcinoma. Int. J. Cancer 2012, 131, E329–E336. [Google Scholar] [CrossRef] [PubMed]
- Sonoda, Y.; Hada, N.; Kaneda, T.; Suzuki, T.; Ohshio, T.; Takeda, T.; Kasahara, T. A synthetic glycosphingolipid-induced antiproliferative effect in melanoma cells is associated with suppression of FAK, AKT, and ERK activation. Biol. Pharm. Bull. 2008, 31, 1279–1283. [Google Scholar] [CrossRef] [PubMed]
- Thang, N.D.; Yajima, I.; Kumasaka, M.Y.; Iida, M.; Suzuki, T.; Kato, M. Deltex-3-like (DTX3L) stimulates metastasis of melanoma through FAK/PI3K/AKT but not MEK/ERK pathway. Oncotarget 2015, 6, 14290–14299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Guan, J.-L. Signal transduction by focal adhesion kinase in cancer. Cancer Metastasis Rev. 2009, 28, 35–49. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Prywes, R. Atf1 phosphorylation by the ERK MAPK pathway is required for epidermal growth factor-induced c-Jun expression. J. Biol. Chem. 2002, 277, 50550–50556. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.B.; Pollack, M.H.; Sosman, J.A. Emerging targeted therapies for melanoma. Expert Opin. Emerg. Drugs 2016, 21, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Hugo, H.; Ackland, M.L.; Blick, T.; Lawrence, M.G.; Clements, J.A.; Williams, E.D.; Thompson, E.W. Epithelial-mesenchymal and mesenchymal-epithelial transitions in carcinoma progression. J. Cell. Physiol. 2007, 213, 374–383. [Google Scholar] [CrossRef] [PubMed]
- Radisky, D.C. Epithelial-mesenchymal transition. J. Cell Sci. 2005, 118, 4325–4326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batlle, E.; Sancho, E.; Francí, C.; Domínguez, D.; Monfar, M.; Baulida, J.; de Herreros, A.G. The transcription factor snail is a repressor of e-cadherin gene expression in epithelial tumour cells. Nat. Cell Biol. 2000, 2, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Bolós, V.; Peinado, H.; Pérez-Moreno, M.A.; Fraga, M.F.; Esteller, M.; Cano, A. The transcription factor Slug represses E-cadherin expression and induces epithelial to mesenchymal transitions: A comparison with snail and E47 repressors. J. Cell Sci. 2003, 116, 499–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cano, A.; Pérez-Moreno, M.A.; Rodrigo, I.; Locascio, A.; Blanco, M.J.; del Barrio, M.G.; Portillo, F.; Nieto, M.A. The transcription factor Snail controls epithelial–mesenchymal transitions by repressing e-cadherin expression. Nat. Cell Biol. 2000, 2, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Nieto, M.A. The snail superfamily of zinc-finger transcription factors. Nat. Rev. Mol. Cell Biol. 2002, 3, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Haass, N.K.; Smalley, K.S.; Li, L.; Herlyn, M. Adhesion, migration and communication in melanocytes and melanoma. Pigment Cell Melanoma Res. 2005, 18, 150–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attis, M.G.; Burchette, J.L.; Selim, M.A.; Pham, T.; Soler, A.P. Differential expression of N-cadherin distinguishes a subset of metastasizing desmoplastic melanomas. Hum. Pathol. 2006, 37, 899–905. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Chen, N.; Wang, J.; Siu, C.-H. Transendothelial migration of melanoma cells involves n-cadherin-mediated adhesion and activation of the β-catenin signaling pathway. Mol. Biol. Cell 2005, 16, 4386–4397. [Google Scholar] [CrossRef] [PubMed]
- Smalley, K.S.; Brafford, P.; Haass, N.K.; Brandner, J.M.; Brown, E.; Herlyn, M. Up-regulated expression of zonula occludens protein-1 in human melanoma associates with N-cadherin and contributes to invasion and adhesion. Am. J. Pathol. 2005, 166, 1541–1554. [Google Scholar] [CrossRef]











© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, T.-Y.; Wu, M.-L.; Chang, C.-I.; Liu, C.-I.; Cheng, T.-C.; Wu, Y.-J. Bornyl cis-4-Hydroxycinnamate Suppresses Cell Metastasis of Melanoma through FAK/PI3K/Akt/mTOR and MAPK Signaling Pathways and Inhibition of the Epithelial-to-Mesenchymal Transition. Int. J. Mol. Sci. 2018, 19, 2152. https://doi.org/10.3390/ijms19082152
Yang T-Y, Wu M-L, Chang C-I, Liu C-I, Cheng T-C, Wu Y-J. Bornyl cis-4-Hydroxycinnamate Suppresses Cell Metastasis of Melanoma through FAK/PI3K/Akt/mTOR and MAPK Signaling Pathways and Inhibition of the Epithelial-to-Mesenchymal Transition. International Journal of Molecular Sciences. 2018; 19(8):2152. https://doi.org/10.3390/ijms19082152
Chicago/Turabian StyleYang, Tzu-Yen, Mei-Li Wu, Chi-I Chang, Chih-I Liu, Te-Chih Cheng, and Yu-Jen Wu. 2018. "Bornyl cis-4-Hydroxycinnamate Suppresses Cell Metastasis of Melanoma through FAK/PI3K/Akt/mTOR and MAPK Signaling Pathways and Inhibition of the Epithelial-to-Mesenchymal Transition" International Journal of Molecular Sciences 19, no. 8: 2152. https://doi.org/10.3390/ijms19082152