Salicylic Acid Induces Resistance in Rubber Tree against Phytophthora palmivora

Abstract
:
1. Introduction
2. Results
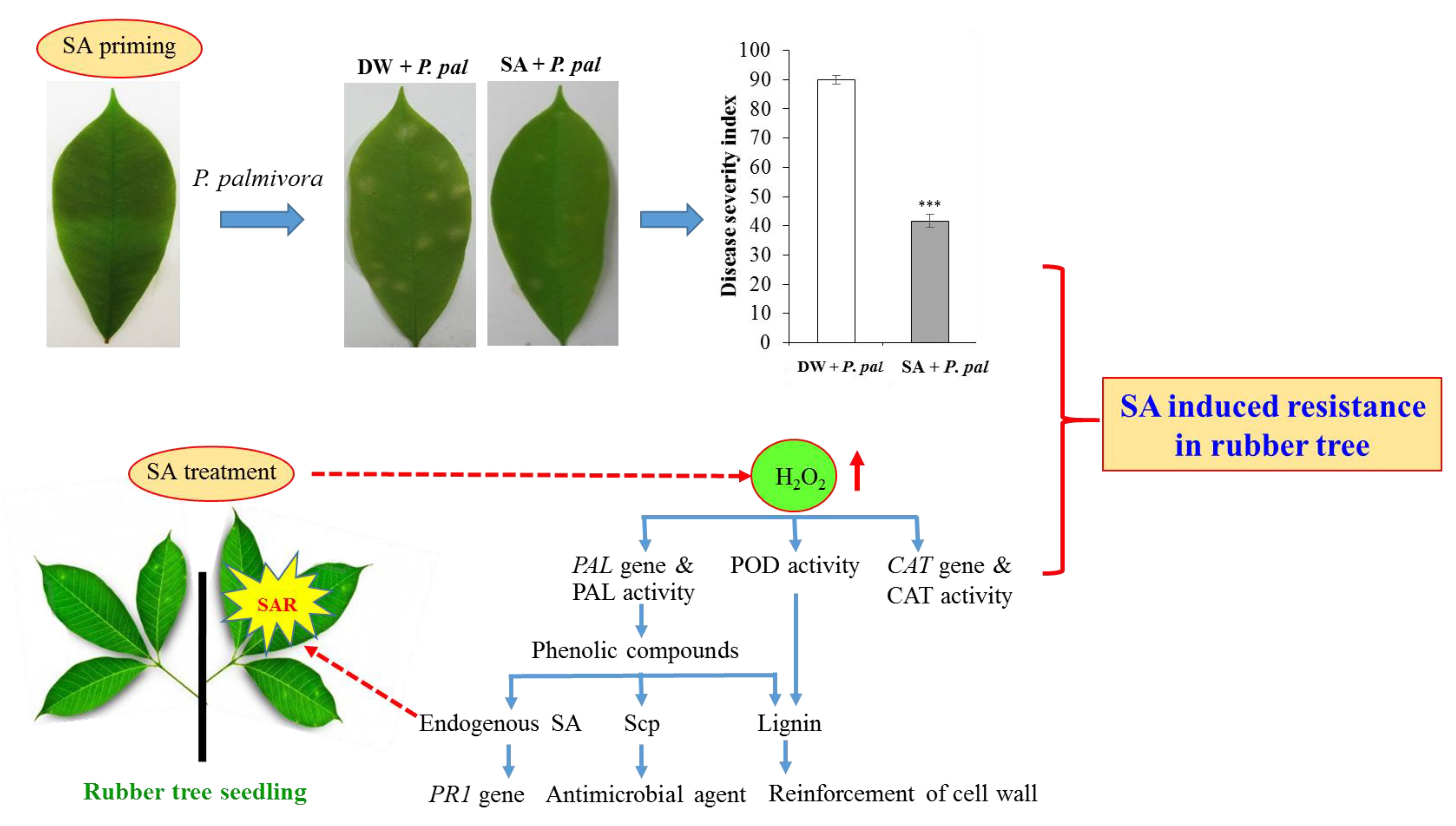
2.1. Effects of SA Priming on Resistance of Rubber Tree
2.1.1. Induced Resistance of SA-Pretreated Rubber Tree against P. palmivora
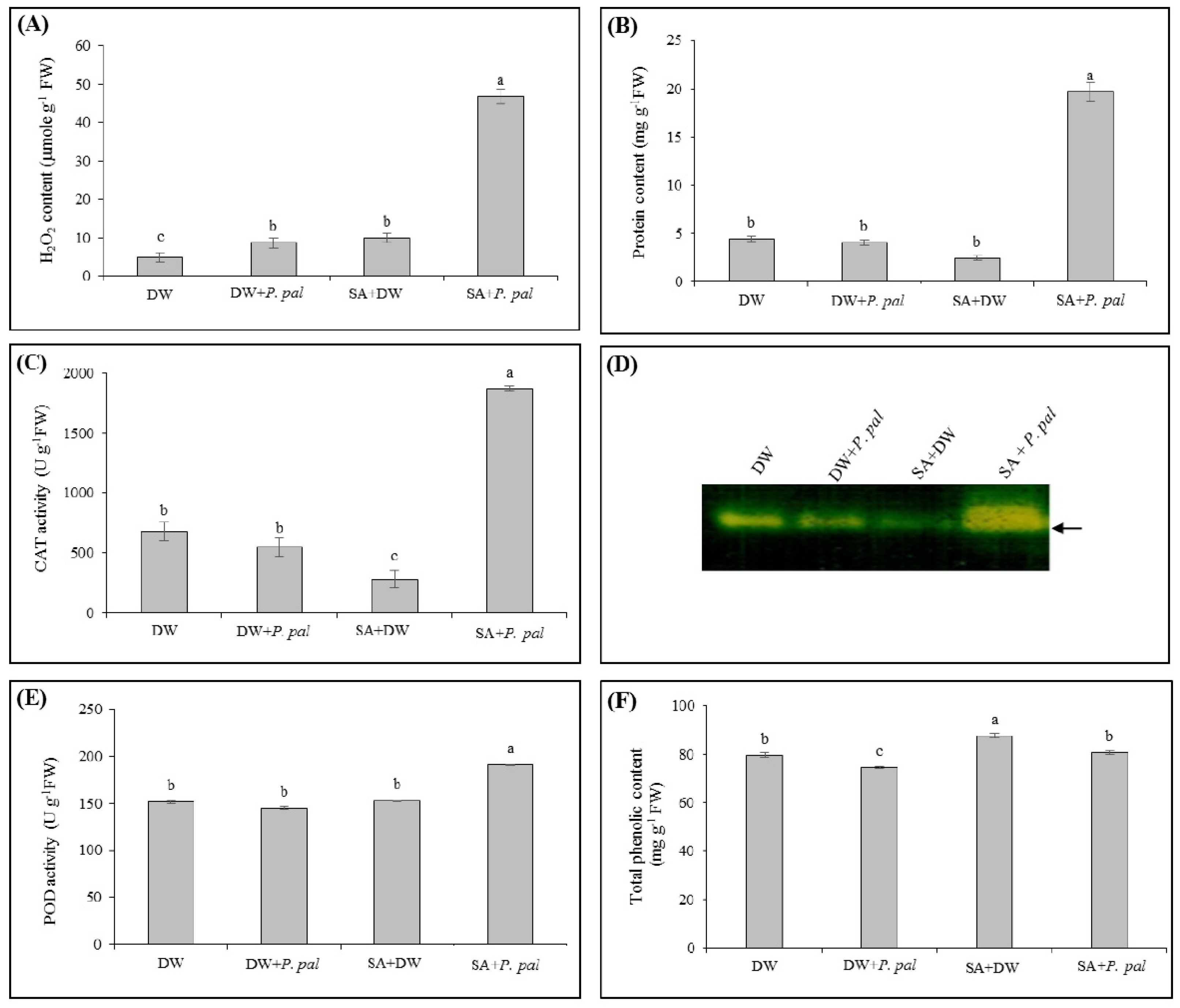
2.1.2. Effect of SA Priming on H2O2 Content, Total Protein Content, CAT and POD Activities and Total Phenolic Content in Rubber Tree Leaves after P. palmivora Challenge
2.2. Effect of SA on Kinetics of H2O2 Content, CAT and PAL Activities, SA and Scp Contents in Rubber Tree Seedling Leaves
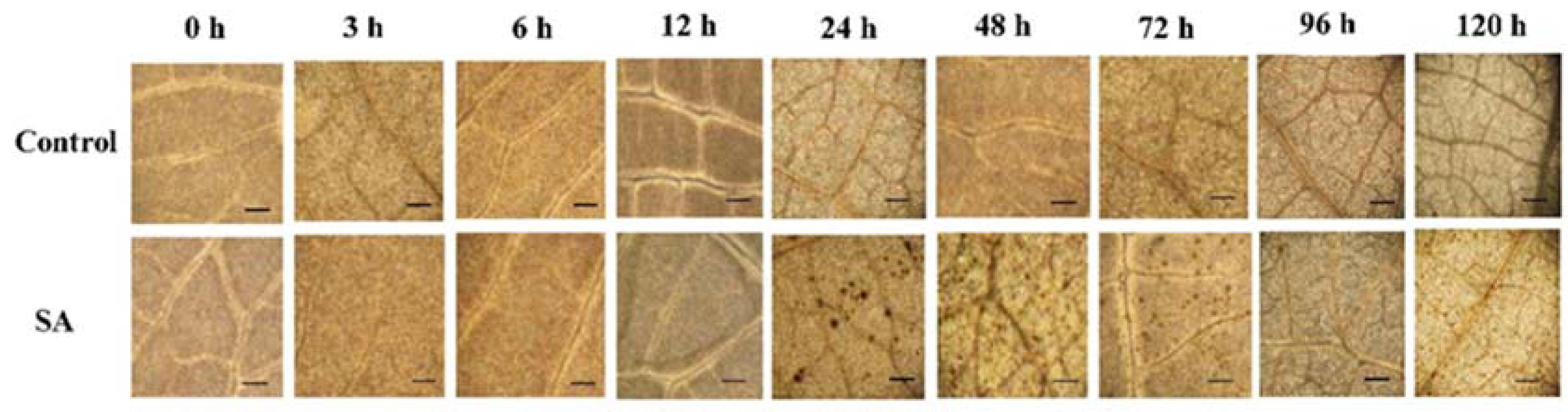
2.2.1. Effect of SA on H2O2 Content
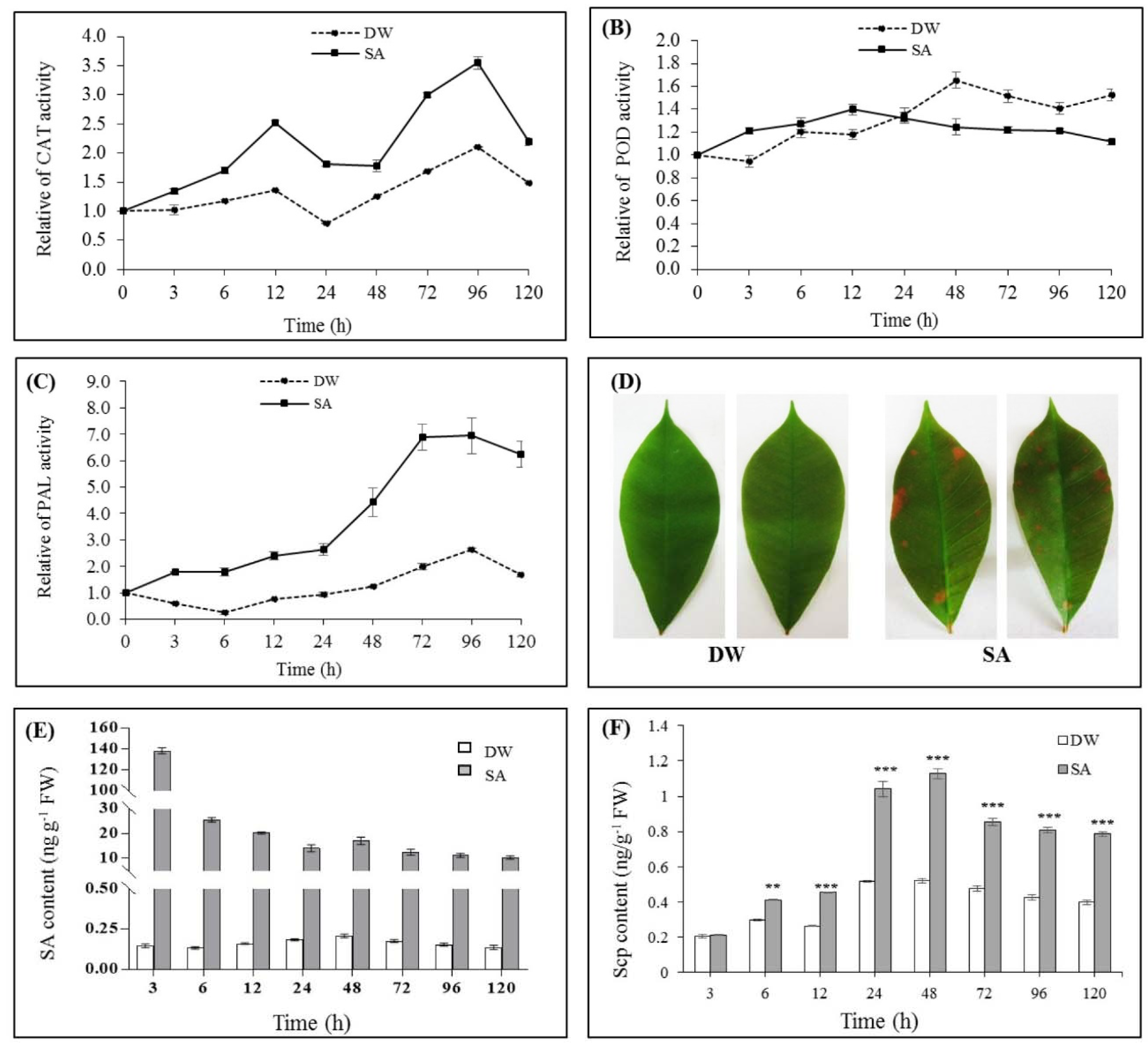
2.2.2. Effect of SA on CAT, POD and PAL Activities
2.2.3. Effect of SA on Lignin Deposition
2.2.4. Effect of SA on Endogenous SA and Scp Levels
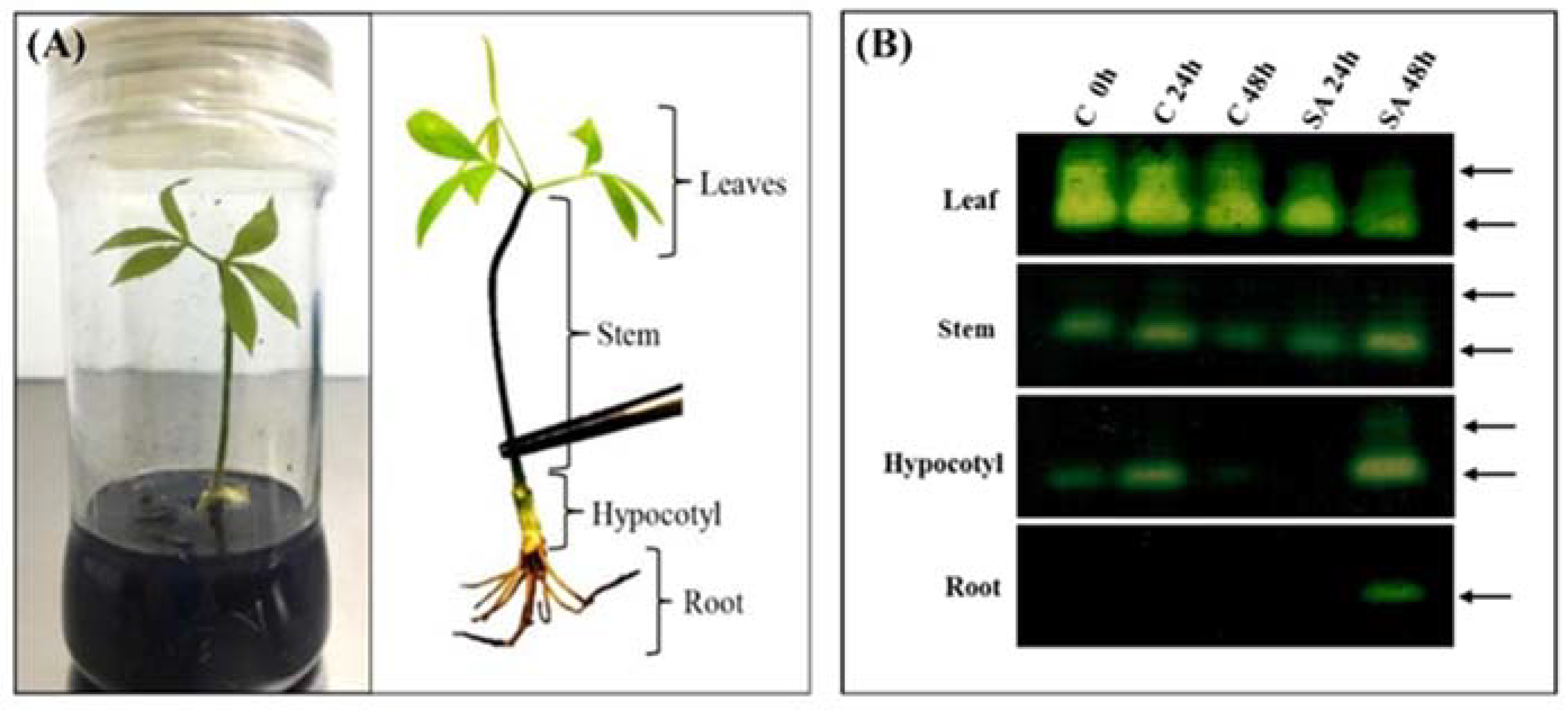
2.3. Effect of SA on CAT Activity in Different Rubber Tree Organs
2.4. Cloning and Sequence Analysis of Three Partial cDNAs Encoding Catalases
2.5. Cloning and Sequence Analysis of a Partial cDNA Encoding Phenylalanine Ammonia Lyase
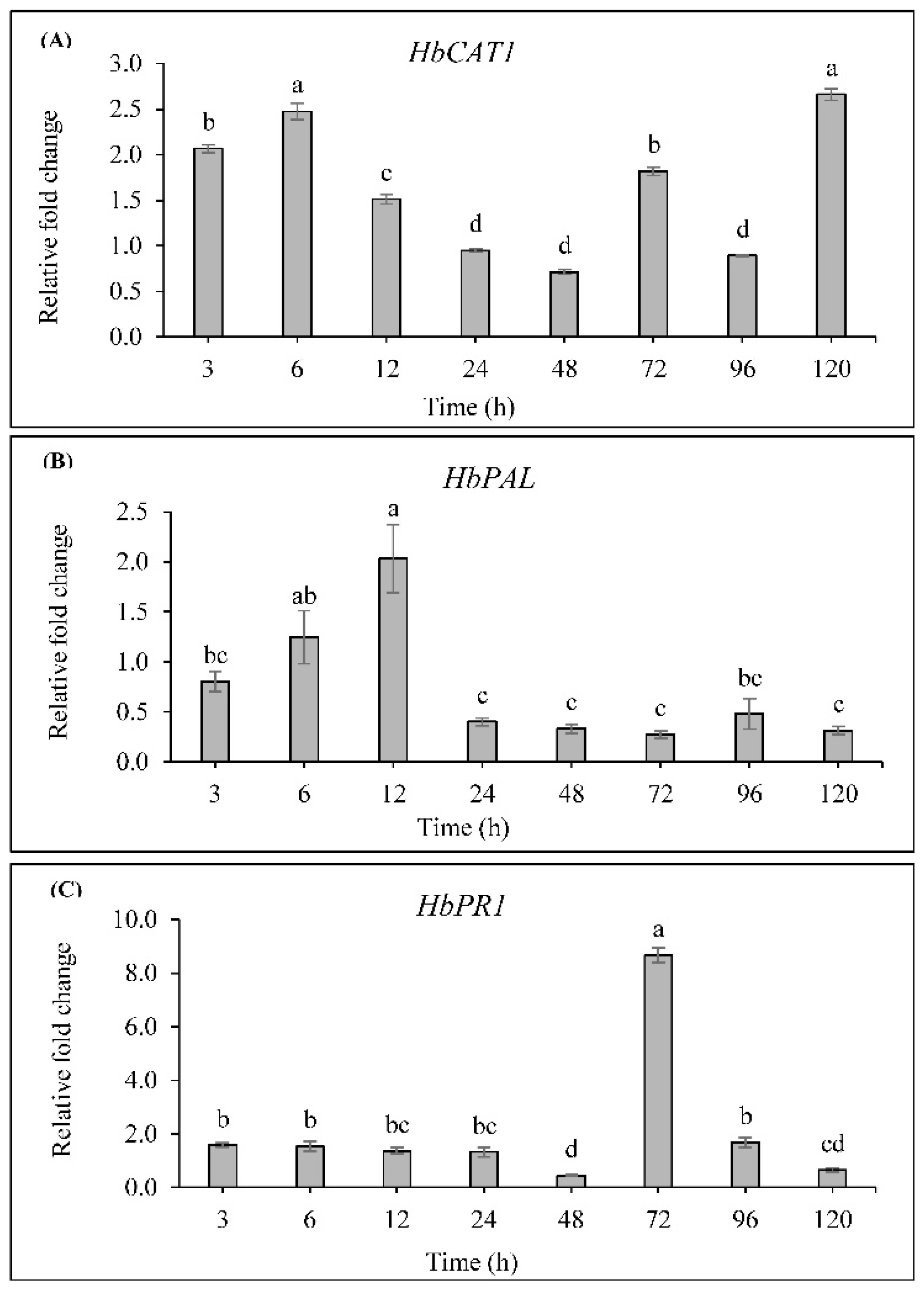
2.6. Effect of SA on HbCAT1, HbPAL and HbPR1 Expressions in H. brasiliensis
3. Discussion
4. Materials and Methods
4.1. Phytophthora palmivora Culture
4.2. Plant Materials and Treatments
4.3. Induced Resistance Bioassays
= {[Σ(n × 0) + (n × 1) + (n × 2) + (n × 3) + (n × 4)] ×100}/(N × Z)
4.4. Protein Extraction, H2O2 Content and Enzyme Activity Assays
4.5. Histochemical detection of H2O2
4.6. CAT Activity Staining after Native Polyacrylamide Gel Electrophoresis (Native-PAGE)
4.7. Total Phenolic Content and Lignin Detection
4.8. SA and Scp Measurements
4.9. Total RNA Isolation and cDNA Synthesis
4.10. Isolation of Partial cDNAs of HbCAT and HbPAL
4.11. Gene Expression Analyses of HbCAT1, HbPR1 and HbPAL By Semi-Quantitative Reverse Transcription Polymerase Chain Reaction (Semi-qRT-PCR)
4.12. Statistical analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Widmer, T.L. Phytophthora palmivora. For. Phytophthoras 2014, 4. [Google Scholar] [CrossRef]
- Wojtaszek, P. Oxidative burst: An early plant response to pathogen infection. Biochem. J. 1997, 322, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Levine, A.; Tenhaken, R.; Dixon, R.; Lamb, C. H2O2 from the oxidative burst orchestrates the plant hypersensitive disease resistance response. Cell 1994, 79, 583–593. [Google Scholar] [CrossRef]
- Dat, J.; Vandenabeele, S.; Vranová, E.; Van Montagu, M.; Inzé, D.; Van Breusegem, F. Dual action of the active oxygen species during plant stress responses. Cell. Mol. Life Sci. 2000, 57, 779–795. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed]
- Bienert, G.P.; Møller, A.L.B.; Kristiansen, K.A.; Schulz, A.; Møller, I.M.; Schjoerring, J.K.; Jahn, T.P. Specific aquaporins facilitate the diffusion of hydrogen peroxide across membranes. J. Biol. Chem. 2007, 282, 1183–1192. [Google Scholar] [CrossRef] [PubMed]
- Neill, S.J.; Desikan, R.; Clarke, A.; Hurst, R.D.; Hancock, J.T. Hydrogen peroxide and nitric oxide as signalling molecules in plants. J. Exp. Bot. 2002, 53, 1237–1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Silva, H.; Klessig, D.F. Active oxygen species in the induction of plant systemic acquired resistance by salicylic acid. Science 1993, 262, 1883–1886. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Breusegem, F.V. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Gratao, P.L.; Polle, A.; Lea, P.J.; Azevedo, R.A. Making the life of heavy metal-stressed plants a little easier. Funct. Plant Biol. 2005, 32, 481–494. [Google Scholar] [CrossRef]
- Radwan, D.E.M.; Fayez, K.A.; Mahmoud, S.Y.; Lu, G. Modifications of antioxidant activity and protein composition of bean leaf due to Bean yellow mosaic virus infection and salicylic acid treatments. Acta Physiol. Plant. 2010, 32, 891–904. [Google Scholar] [CrossRef]
- Mullen, P.E.; Pathé, M.; Purcell, R.; Stuart, G.W. Study of Stalkers. Am. J. Psychiatry 1999, 156, 1244–1249. [Google Scholar] [PubMed]
- Mhamdi, A.; Queval, G.; Chaouch, S.; Vanderauwera, S.; Breusegem, F.V.; Noctor, G. Catalase function in plants: A focus on Arabidopsis mutants as stress-mimic models. J. Exp. Bot. 2010, 61, 4197–4220. [Google Scholar] [CrossRef] [PubMed]
- Banci, L. Structural properties of peroxidases. J. Biotechnol. 1997, 53, 253–263. [Google Scholar] [CrossRef]
- Passardi, F.; Cosio, C.; Penel, C.; Dunand, C. Peroxidases have more functions than a Swiss army knife. Plant Cell Rep. 2005, 24, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Cosio, C.; Dunand, C. Specific functions of individual class III peroxidase genes. J. Exp. Bot. 2009, 60, 391–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seevers, P.M.; Daly, J.M.; Catedral, F.F. The Role of peroxidase isozymes in resistance to wheat stem rust disease. Plant Physiol. 1971, 48, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Kumar, L.; Mahatma, M.K.; Kalariya, K.A.; Bishi, S.K.; Mann, A. Plant phenolics: Important bio-weapon against pathogens and insect herbivores. Pop. Kheti 2014, 2, 149–152. [Google Scholar]
- Dempsey, D.A.; Vlotb, A.C.; Wildermuth, M.C.; Klessig, D.F. SA biosynthesis and metabolism. Arabidopsis Book 2011, 9, e0156. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Wan, G.; Liang, Z. Accumulation of salicylic acid-induced phenolic compounds and raised activities of secondary metabolic and antioxidative enzymes in Salvia miltiorrhiza cell culture. J. Biotechnol. 2010, 148, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Ahl-Goy, P.; Signer, H.; Reist, R.; Aichholz, R.; Blum, W.; Schmidt, E.; Kessmann, H. Accumulation of scopoletin is associated with the high disease resistance of the hybrid Nicotiana glutinosa × Nicotiana debneyi. Planta 1993, 191, 200–206. [Google Scholar] [CrossRef]
- Kuyyogsuy, A.; Deenamo, N.; Khompatara, K.; Ekchaweng, K.; Churngchow, N. Chitosan enhances resistance in rubber tree (Hevea brasiliensis), through the induction of abscisic acid (ABA). Physiol. Mol. Plant Pathol. 2018, 102, 67–78. [Google Scholar] [CrossRef]
- Bagal, U.R.; Leebens-Mack, J.H.; Lorenz, W.W.; Dean, J.F.D. The phenylalanine ammonia lyase (PAL) gene family shows a gymnosperm-specific lineage. BMC Genom. 2012, 13 (Suppl. 3), S1. [Google Scholar] [CrossRef]
- Kumar, A.; Ellis, B.E. The phenylalanine ammonia-lyase gene family in raspberry. Structure, expression, and evolution. Plant Physiol. 2001, 127, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Gu, M.; Lai, Z.; Fan, B.; Shi, K.; Zhou, Y.H.; Yu, J.Q.; Chen, Z. Functional analysis of the Arabidopsis PAL gene family in plant growth, development, and response to environmental stress. Plant Physiol. 2010, 153, 1526–1538. [Google Scholar] [CrossRef] [PubMed]
- Durrant, W.E.; Dong, X. Systemic acquired resistance. Annu. Rev. Phytopathol. 2004, 42, 185–209. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.C.; Kidd, B.N.; Carvalhais, L.C.; Schenk, P.M. Molecular defense responses in roots and the rhizosphere against Fusarium oxysporum. Plant Signal. Behav. 2014, 9, e977710. [Google Scholar] [CrossRef] [PubMed]
- Metraux, J.P. Systemic acquired resistance and salicylic acid: Current state of knowledge. Eur. J. Plant Pathol. 2001, 107, 13–18. [Google Scholar] [CrossRef]
- Mur, L.A.J.; Naylor, G.; Warner, S.A.J.; Sugars, J.M.; White, R.F.; Draper, J. Salicylic acid potentiates defence gene expression in tissue exhibiting acquired resistance to pathogen attack. Plant J. 1996, 9, 559–571. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Shi, X.P.; Li, B.H.; Zhang, Q.M.; Liang, W.X.; Wang, C.X. Salicylic acid confers enhanced resistance to Glomerella leaf spot in apple, Plant Physiol. Biochem. 2016, 106, 64–72. [Google Scholar] [CrossRef]
- Conrath, U.; Pieterse, C.M.J.; Mauch-Mani, B. Priming in plant–pathogen interactions. Trends Plant Sci. 2002, 7, 210–216. [Google Scholar] [CrossRef] [Green Version]
- Mandal, S.; Mallick, N.; Mitra, A. Salicylic acid-induced resistance to Fusarium oxysporum f. sp. lycopersici in tomato. Plant Physiol. Biochem. 2009, 47, 642–649. [Google Scholar] [CrossRef] [PubMed]
- Bernsdorff, F.; Döring, A.C.; Gruner, K.; Schuck, S.; Bräutigam, A.; Zeier, J. Pipecolic acid orchestrates plant systemic acquired resistance and defense priming via salicylic acid-dependent and -independent pathways. Plant Cell 2016, 28, 102–129. [Google Scholar] [CrossRef] [PubMed]
- Bastas, K.K. Importance of Reactive Oxygen Species in Plants-Pathogens Interactions. Selcuk J. Agric. Food Sci. 2014, 28, 11–21. [Google Scholar]
- Bhattacharjee, S. Reactive oxygen species and oxidative burst: Roles in stress, senescence and signal transduction in plants. Curr. Sci. 2005, 89, 1113–1121. [Google Scholar]
- Mejía-Teniente, L.; de Dalia Duran-Flores, F.; Chapa-Oliver, A.M.; Torres-Pacheco, I.; Cruz-Hernández, A.; González-Chavira, M.M.; Ocampo-Velázquez, R.V.; Guevara-González, R.G. Oxidative and molecular responses in Capsicum annuum L. after hydrogen peroxide, salicylic acid and chitosan foliar applications. Int. J. Mol. Sci. 2013, 14, 10178–10196. [Google Scholar] [CrossRef] [PubMed]
- Tomas-Barberan, F.A.; Espın, J.C. Phenolic compounds and related enzymes as determinants of quality in fruits and vegetables. J. Sci. Food Agric. 2001, 81, 853–876. [Google Scholar] [CrossRef]
- Tsai, C.J.; Harding, S.A.; Tschaplinski, T.J.; Lindroth, R.L.; Yuan, Y. Genome-wide analysis of the structural genes regulating defense phenylpropanoid metabolism in Populus. New Phytol. 2006, 172, 47–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandal, S.; Kar, I.; Mukherjee, A.K.; Acharya, P. Elicitor-Induced Defense Responses in Solanum lycopersicum against Ralstonia solanacearum. Sci. World J. 2013, 2013, 561056. [Google Scholar] [CrossRef] [PubMed]
- De Gara, L.; de Pinto, M.C.; Tommasi, F. The antioxidant systems vis-à-vis reactive oxygen species during plant–pathogen interaction. Plant Physiol. Biochem. 2003, 41, 863–870. [Google Scholar] [CrossRef]
- Hao, W.; Guo, H.; Zhang, J.; Hu, G.; Yao, Y.; Dong, J. Hydrogen peroxide is involved in salicylic acid-elicited rosmarinic acid production in Salvia miltiorrhiza cell cultures. Sci. World J. 2014, 2014, 843764. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Davis, L.C.; Verpoorte, R. Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol. Adv. 2005, 23, 283–333. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S. Induction of phenolics, lignin and key defense enzymes in eggplant (Solanum melongena L.) roots in response to elicitors. Afr. J. Biotechnol. 2010, 9, 8038–8047. [Google Scholar] [CrossRef]
- Churngchow, N.; Rattarasarn, M. Biosynthesis of scopoletin in Hevea brasiliensis leaves inoculated with Phytophthora palmivora. J. Plant Physiol. 2001, 158, 875–882. [Google Scholar] [CrossRef]
- Dorey, S.; Kopp, M.; Geoffroy, P.; Fritig, B.; Kauffmann, S. Hydrogen peroxide from the oxidative burst is neither necessary nor sufficient for hypersensitive cell death induction, phenylalanine ammonia lyase stimulation, salicylic acid accumulation, or scopoletin consumption in cultured tobacco cells treated with elicitin. Plant Physiol. 1999, 121, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Ananieva, E.A.; Christov, K.N.; Popova, L.P. Exogenous treatment with salicylic acid leads to increased antioxidant capacity in leaves of barley plants exposed to paraquat. J. Plant Physiol. 2004, 161, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Shim, I.S.; Momose, Y.; Yamamoto, A.; Kim, D.W.; Usui, K. Inhibition of catalase activity by oxidative stress and its relationship to salicylic acid accumulation in plants. Plant Growth Regul. 2003, 39, 285–292. [Google Scholar] [CrossRef]
- Rüffer, M.; Steipe, B.; Zenk, M.H. Evidence against specific binding of salicylic acid to plant catalase. FEBS Lett. 1995, 377, 175–180. [Google Scholar] [CrossRef] [Green Version]
- Conrath, U.; Chen, Z.; Ricigliano, J.R.; Klessig, D.F. Two inducers of plant defense responses, 2,6-dichloroisonicotinic acid and salicylic acid, inhibit catalase activity in tobacco. Proc. Natl. Acad. Sci. USA 1995, 92, 7143–7147. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.V.; Paliyath, G.; Ormrod, D.P.; Murr, D.P.; Watkins, C.B. Influence of salicylic acid on H2O2 production, oxidative stress, and H2O2-metabolizing enzymes. Plant Physiol. 1997, 115, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Dat, J.F.; Lopez-Delgado, H.; Foyer, C.H.; Scotth, I.M. Effects of salicylic acid on oxidative stress and thermotolerance in tobacco. J. Plant Physiol. 2000, 156, 659–665. [Google Scholar] [CrossRef]
- Durner, J.; Klessig, D.F. Salicylic acid is a modulator of tobacco and mammalian catalases. J. Biol. Chem. 1996, 271, 28492–28501. [Google Scholar] [CrossRef] [PubMed]
- Gayatridevi, S.; Jayalakshmi, S.K.; Mulimani, V.H.; Sreeramulu, K. Salicylic acid and salicylic acid sensitive and insensitive catalases in different genotypes of chickpea against Fusarium oxysporum f. sp. ciceri. Physiol. Mol. Biol. Plants 2013, 19, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Wutipraditkul, N.; Boonkomrat, S.; Buaboocha, T. Cloning and characterization of catalase from rice, Oryza sativa L. Biosci. Biotechnol. Biochem. 2011, 75, 1900–1906. [Google Scholar] [CrossRef] [PubMed]
- Guan, L.; Scandalios, J.G. Developmentally related responses of maize catalase genes to salicylic acid. Proc. Natl. Acad. Sci. USA 1995, 92, 5930–5934. [Google Scholar] [CrossRef] [PubMed]
- Gayatridevi, S.; Jayalakshmi, S.K.; Sreeramulu, K. Salicylic acid is a modulator of catalase isozymes in chickpea plants infected with Fusarium oxysporum f. sp. ciceri. Plant Physiol. Biochem. 2012, 52, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Drory, A.; Woodson, W.R. Molecular cloning and nucleotide sequence of a cDNA encoding catalase from tomato. Plant Physiol. 1992, 100, 1605–1606. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Deng, G.; Cheng, S.; Zhang, W.; Huang, X.; Li, L.; Cheng, H.; Rong, X.; Li, J. Molecular cloning, characterization and expression of the phenylalanine ammonia-lyase gene from Juglans regia. Molecules 2012, 17, 7810–7823. [Google Scholar] [CrossRef] [PubMed]
- Belkadhi, A.; De Haro, A.; Soengas, P.; Obregon, S.; Cartea, M.E.; Chaibi, W.; Djebali, W. Salicylic acid increases tolerance to oxidative stress induced by hydrogen peroxide accumulation in leaves of cadmium-exposed flax (Linum usitatissimum L.). J. Plant Interact. 2014, 9, 647–654. [Google Scholar] [CrossRef] [Green Version]
- Kwon, S.I.; Lee, H.; An, C.S. Differential expression of three catalase genes in the small radish (Rhaphanus sativus L. var. sativus). Mol. Cells 2007, 24, 37–44. [Google Scholar] [PubMed]
- Lee, S.H.; An, C.S. Differential expression of three catalase genes in hot pepper (Capsicum annuum L.). Mol. Cells 2005, 20, 247–255. [Google Scholar] [PubMed]
- Jeong, M.-J.; Choi, B.S.; Bae, D.W.; Shin, S.C.; Park, S.U.; Lim, H.-S.; Kim, J.; Kim, J.B.; Cho, B.-K.; Bae, H. Differential expression of kenaf phenylalanine ammonia-lyase (PAL) ortholog during developmental stages and in response to abiotic stresses. POJ 2012, 5, 392–399. [Google Scholar]
- Kolahi, M.; Jonoubi, P.; Majd, A.; Tabandeh, M.R.; Hashemitabar, M. Differential expression of phenylalanine ammonia-lyase in different tissues of sugarcane (Saccharum officinarum L.) during development. Bioresources 2013, 8, 4912–4922. [Google Scholar] [CrossRef]
- Chen, Z.; Zheng, Z.; Huang, J.; Lai, Z.; Fan, B. Biosynthesis of salicylic acid in plants. Plant Signal. Behav. 2009, 4, 493–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, P.F.; Chen, J.Y.; Kong, W.F.; Pan, Q.H.; Wan, S.B.; Huang, W.D. Salicylic acid induced the expression of phenylalanine ammonia-lyase gene in grape berry. Plant Sci. 2005, 169, 928–934. [Google Scholar] [CrossRef]
- Molinari, S.; Fanelli, E.; Leonetti, P. Expression of tomato salicylic acid (SA)-responsive pathogenesis-related genes in Mi-1-mediated and SA-induced resistance to root-knot nematodes. Mol. Plant Pathol. 2014, 15, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.S.; Liu, R.; Zheng, L. Expression analysis of wheat PR-1, PR-2, PR-5 activated by Bgt and SA, and powdery mildew resistance. J. Triticeae Crops 2007, 27, 1132–1137. [Google Scholar]
- Grellet-Bournonville, C.F.; Martinez-Zamora, M.G.; Castagnaro, A.P.; Díaz-Ricci, J.C. Temporal accumulation of salicylic acid activates the defense response against Colletotrichum in strawberry. Plant Physiol. Biochem. 2012, 54, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Glazebrook, J. Genes controlling expression of defense responses in Arabidopsis—2001 status. Curr. Opin. Plant Biol. 2001, 4, 301–308. [Google Scholar] [CrossRef]
- Ekchaweng, K.; Khunjun, U.; Churngchow, N. Molecular cloning and characterization of three novel subtilisin-like serine protease genes from Hevea brasiliensis, Physiol. Mol. Plant Pathol. 2017, 97, 79–95. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Kranz, J. Measuring plant disease. In Experimental Techniques in Plant Disease Epidemiology, 1st ed.; Kranz, J., Rotem, J., Eds.; Springer: Berlin/Heidelberg, Germany, 1988; pp. 35–50. ISBN 978-3-642-95534-1. [Google Scholar]
- Hadwan, M.H.; Abed, H.N. Data supporting the spectrophotometric method for the estimation of catalase activity. Data Brief 2016, 6, 194–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Jiang, W.; Bi, Y.; Luo, Y. Postharvest BTH treatment induces resistance of peach (Prunus persica L. cv. Jiubao) fruit to infection by Penicillium expansum and enhances activity of fruit defense mechanisms. Postharvest Biol. Technol. 2005, 35, 263–269. [Google Scholar] [CrossRef]
- D’Cunha, G.B.; Satyanarayan, V.; Nair, P.M. Stabilization of phenylalanine ammonia lyase containing Rhodotorula glutinis cells for the continuous synthesis of l-phenylalanine methyl ester/96. Enzym. Microb. Technol. 1996, 19, 421–427. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Thordal-Christensen, H.; Zhang, Z.; Wei, Y.; Collinge, D.B. Subcellular localization of H2O2 in plants. H2O2 accumulation in papillae and hypersensitive response during the barley powdery mildew interaction. Plant J. 1997, 11, 1187–1194. [Google Scholar] [CrossRef]
- Davis, B.J. Disc electrophoresis II: Method and application to human serum protein. Ann. N. Y. Acad. Sci. 1964, 121, 404–427. [Google Scholar] [CrossRef] [PubMed]
- Woodbury, W.; Spencer, A.K.; Stahman, M.A. An improved procedure for using ferricyanide for detecting catalase isozymes. Anal. Biochem. 1971, 44, 301–305. [Google Scholar] [CrossRef]
- Torres, A.M.; Mau-Lastovicka, T.; Rezaaiyan, R. Total phenolics and high performance liquid chromatography of phenolic acids of avocado. J. Agric. Food Chem. 1987, 35, 921–925. [Google Scholar] [CrossRef]
- Jensen, W.A. Botanical Histochemistry: Principles and Practice; W.H. Freeman & Co.: San Francisco, CA, USA, 1962; Volume viii, 408p, ISBN 978-938-328-525-9. [Google Scholar]
- Ederli, L.; Madeo, L.; Calderini, O.; Gehring, C.; Moretti, C.; Buonaurio, R.; Paolocci, F.; Pasqualini, S. The Arabidopsis thaliana cysteine-rich receptor-like kinase CRK20 modulates host responses to Pseudomonas syringae cv. tomato DC3000 infection. J. Plant Physiol. 2011, 168, 1784–1794. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Qin, Y.; Xiao, X.; Tang, C. Screening of valid reference genes for real-time RT-PCR data normalization in Hevea brasiliensis and expression validation of a sucrose transporter gene HbSUT3. Plant Sci. 2011, 181, 132–139. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Names | Sequence (5′→3′) | Tm (°C) | Base Pairs (bp) |
|---|---|---|---|
| CAT-F | ACWTGTGCTGATTTCCTTCGAG | 55.0–60.3 | 22 |
| CAT-R | ATGGTGATTGTTGTGATGAGCACAC | 58.6–64.2 | 25 |
| PAL-F | CATTTGGATGARGTGAARARAATGGT | 51.7–56.4 | 26 |
| PAL-R | AGTTSACRTCTTGGTTGTGTTGCTC | 56.0–57.7 | 25 |
| Genes | Primer Names | Sequence (5′→3′) | Amplicon Size (bp) | GenBank Accession No. |
|---|---|---|---|---|
| HbCAT1 | HbCAT1-F | CCTGTCATTGTCCGTTTCTCCACTG | 495 | MF383167 |
| HbCAT1-R | CCAGCGGCAATGGAGTCATATAAATCC | |||
| HbPAL | HbPAL-F | TGAACGAGGGAACTGATAGCTATGGTG | 736 | MG992015 |
| HbPAL-R | GGTTTCTGCAAGGGATCCATTTCATGC | |||
| HbPR1 | HbPR1-F | ATGCCCATAACCAAGCACGAGCAG | 364 | KM514666 |
| HbPR1-R | CCAGGAGGGTCGTAGTTGCATCCA | |||
| HbMitosis | HbMito-F | TGGGCTGTTGATCAGGCAATCTTGGC | 577 | HQ323250 |
| HbMito-R | TGTCAGATACATTGCTGCACACAAGGC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deenamo, N.; Kuyyogsuy, A.; Khompatara, K.; Chanwun, T.; Ekchaweng, K.; Churngchow, N. Salicylic Acid Induces Resistance in Rubber Tree against Phytophthora palmivora. Int. J. Mol. Sci. 2018, 19, 1883. https://doi.org/10.3390/ijms19071883
Deenamo N, Kuyyogsuy A, Khompatara K, Chanwun T, Ekchaweng K, Churngchow N. Salicylic Acid Induces Resistance in Rubber Tree against Phytophthora palmivora. International Journal of Molecular Sciences. 2018; 19(7):1883. https://doi.org/10.3390/ijms19071883
Chicago/Turabian StyleDeenamo, Nuramalee, Arnannit Kuyyogsuy, Khemmikar Khompatara, Thitikorn Chanwun, Kitiya Ekchaweng, and Nunta Churngchow. 2018. "Salicylic Acid Induces Resistance in Rubber Tree against Phytophthora palmivora" International Journal of Molecular Sciences 19, no. 7: 1883. https://doi.org/10.3390/ijms19071883
APA StyleDeenamo, N., Kuyyogsuy, A., Khompatara, K., Chanwun, T., Ekchaweng, K., & Churngchow, N. (2018). Salicylic Acid Induces Resistance in Rubber Tree against Phytophthora palmivora. International Journal of Molecular Sciences, 19(7), 1883. https://doi.org/10.3390/ijms19071883