Protective Effects of Xyloglucan in Association with the Polysaccharide Gelose in an Experimental Model of Gastroenteritis and Urinary Tract Infections

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
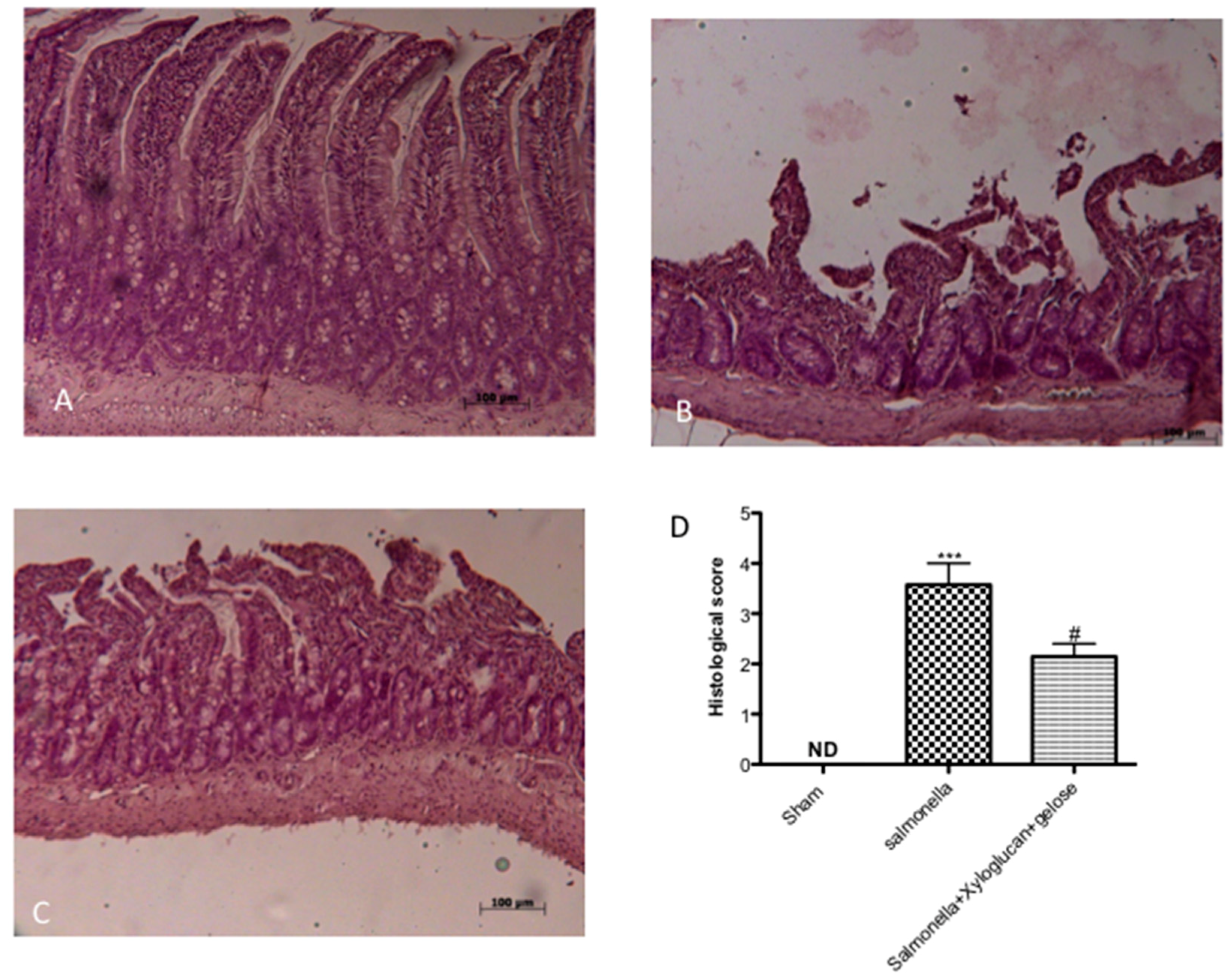
2.1. Effect of Xyloglucan-Gelose on Intestinal Damage and Neutrophil Infiltration
2.2. Effect of Xyloglucan-Gelose on TJ Permeability
2.3. Bactericidal or Bacteriostatic Effect of Xyloglucan-Gelose
2.4. Effect of Xyloglucan-Gelose on Urine Volume and pH
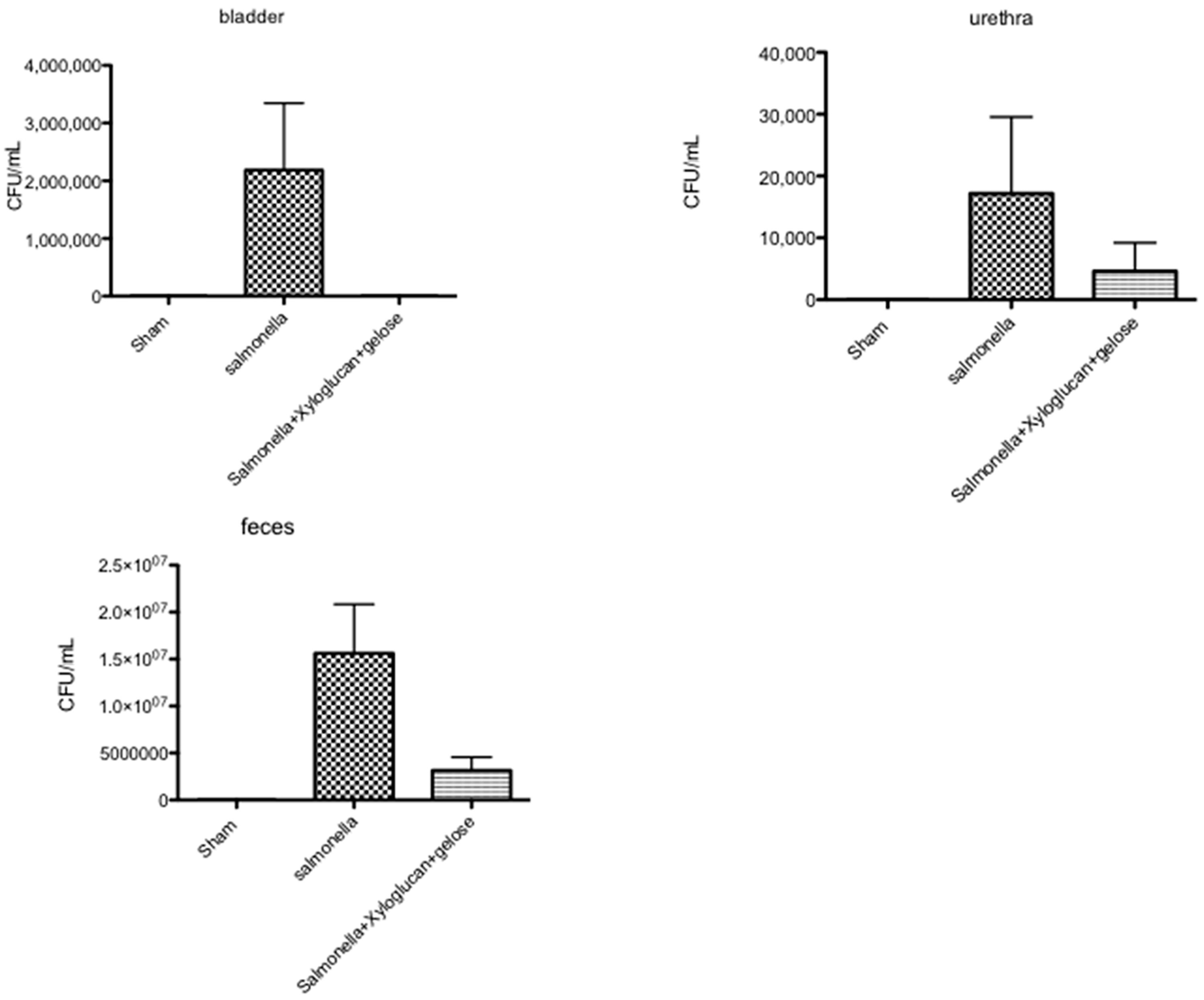
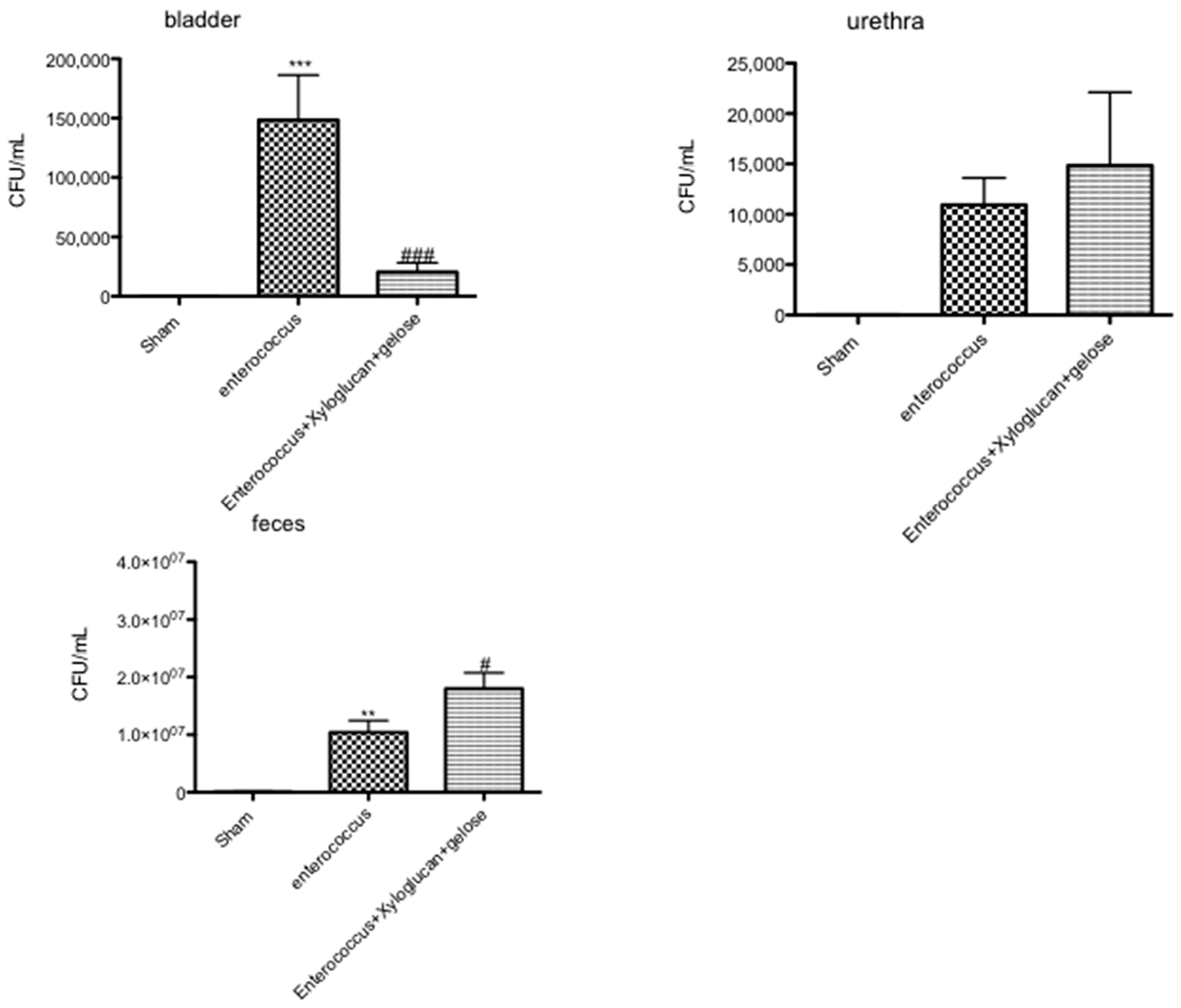
2.5. Effect of Xyloglucan-Gelose on Bacterial Infection of the Urinary Tract
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animals
4.3. Salmonella enterica and Enterococcus hirae Infection
- -
- Feces were collected for bacterial analysis.
- -
- Urine was collected for pH measurement, bacterial counts, volume evaluation, and chemical analysis.
- -
- Bladder and the low urethra were collected for primary cultures evaluating bacterial count.
- -
- Intestine was collected for histological evaluation, myeloperoxidase assay, and immunostaining of tight junctions.
4.4. Experimental Groups
- (1)
- Control group (vehicle)—no S. enterica
- (2)
- Control group (vehicle)—no E. hirae
- (3)
- S. enterica group (2 × 109 CFU/mL)
- (4)
- E. hirae group (3 × 108 CFU/mL)
- (5)
- S. enterica + xyloglucan (10 mg/kg daily) + gelose (5 mg/kg daily)
- (6)
- E. hirae + xyloglucan (10 mg/kg daily) + gelose (5 mg/kg daily)
4.5. Histological Examination
4.6. Myeloperoxidase Assay
4.7. Immunohistochemical Localization of ZO-1 and Occludin
4.8. Bactericidal or Bacteriostatic Study
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cortes-Penfield, N.W.; Trautner, B.W.; Jump, R.L.P. Urinary Tract Infection and Asymptomatic Bacteriuria in Older Adults. Infect. Dis. Clin. N. Am. 2017, 31, 673–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skyum, F.; Andersen, V.; Chen, M.; Pedersen, C.; Mogensen, C.B. Infectious Gastroenteritis and the Need for Strict Contact Precaution Procedures in Adults Presenting in the Emergency Department—A Danish Register-based Study. J. Hosp. Infect. 2018, 98, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Finlay, B.B.; Brumell, J.H. Salmonella interactions with host cells: In vitro to in vivo. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2000, 355, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Eckmann, L.; Kagnoff, M.F.; Fierer, J. Epithelial cells secrete the chemokine interleukin-8 in response to bacterial entry. Infect. Immun. 1993, 61, 4569–4574. [Google Scholar] [PubMed]
- Jung, H.C.; Eckmann, L.; Yang, S.K.; Panja, A.; Fierer, J.; Morzycka-Wroblewska, E.; Kagnoff, M.F. A distinct array of proinflammatory cytokines is expressed in human colon epithelial cells in response to bacterial invasion. J. Clin. Investig. 1995, 95, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Gorelik, Y.; Paul, M.; Geffen, Y.; Khamaisi, M. Urinary Tract Infections Due to Nontyphoidal Salmonella. Am. J. Med. Sci. 2017, 353, 529–532. [Google Scholar] [CrossRef] [PubMed]
- Gulcan, A.; Bayram, P.; Levent, B.; Gulcan, E. A case of urinary tract infection due to Salmonella enterica serovar Virchow and review of the related literature. Acta Microbiol. Immunol. Hung. 2012, 59, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Leung, A.K.; Kao, C.P.; Robson, W.L. Urinary tract infection due to Salmonella stanleyville in an otherwise healthy child. J. Natl. Med. Assoc. 2005, 97, 281–283. [Google Scholar] [PubMed]
- Gilad, J.; Borer, A.; Riesenberg, K.; Peled, N.; Shnaider, A.; Schlaeffer, F. Enterococcus hirae septicemia in a patient with end-stage renal disease undergoing hemodialysis. Eur. J. Clin. Microbiol. Infect. Dis. 1998, 17, 576–577. [Google Scholar] [CrossRef] [PubMed]
- Etheridge, M.E.; Yolken, R.H.; Vonderfecht, S.L. Enterococcus hirae implicated as a cause of diarrhea in suckling rats. J. Clin. Microbiol. 1988, 26, 1741–1744. [Google Scholar] [PubMed]
- Bourafa, N.; Loucif, L.; Boutefnouchet, N.; Rolain, J.M. Enterococcus hirae, an unusual pathogen in humans causing urinary tract infection in a patient with benign prostatic hyperplasia: First case report in Algeria. New Microbes New Infect. 2015, 8, 7–9. [Google Scholar] [CrossRef] [PubMed]
- Di Lodovico, S.; Cataldi, V.; Di Campli, E.; Ancarani, E.; Cellini, L.; Di Giulio, M. Enterococcus hirae biofilm formation on hospital material surfaces and effect of new biocides. Environ. Health Prev. Med. 2017, 22, 63. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Dwivedi, M.; Mahdi, A.A.; Nagana Gowda, G.A.; Khetrapal, C.L.; Bhandari, M. Inhibition of adherence of multi-drug resistant E. coli by proanthocyanidin. Urol. Res. 2012, 40, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Howell, A.B. Cranberry proanthocyanidins and the maintenance of urinary tract health. Crit. Rev. Food Sci. Nutr. 2002, 42 (Suppl. 3), 273–278. [Google Scholar] [CrossRef] [PubMed]
- Howell, A.B.; Botto, H.; Combescure, C.; Blanc-Potard, A.B.; Gausa, L.; Matsumoto, T.; Tenke, P.; Sotto, A.; Lavigne, J.P. Dosage effect on uropathogenic Escherichia coli anti-adhesion activity in urine following consumption of cranberry powder standardized for proanthocyanidin content: A multicentric randomized double blind study. BMC Infect. Dis. 2010, 10, 94. [Google Scholar] [CrossRef] [PubMed]
- Chisholm, A.H. Probiotics in Preventing Recurrent Urinary Tract Infections in Women: A Literature Review. Urol. Nurs. 2015, 35, 18–21, 29. [Google Scholar] [PubMed]
- De Servi, B.; Ranzini, F.; Pique, N. Effect of Utipro((R)) (containing gelatin-xyloglucan) against Escherichia coli invasion of intestinal epithelial cells: Results of an in vitro study. Future Microbiol. 2016, 11, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Salvatorelli, N.; Garcia-Larrosa, A.; Allegrini, A.; Pavone, D. A New Approach to the Treatment of Uncomplicated Cystitis: Results of a Randomized Placebo-Controlled Clinical Trial. Urol. Int. 2016, 97, 347–351. [Google Scholar] [CrossRef] [PubMed]
- Gnessi, L.; Bacarea, V.; Marusteri, M.; Pique, N. Xyloglucan for the treatment of acute diarrhea: Results of a randomized, controlled, open-label, parallel group, multicentre, national clinical trial. BMC Gastroenterol. 2015, 15, 153. [Google Scholar] [CrossRef] [PubMed]
- Plesea Condratovici, C.; Bacarea, V.; Pique, N. Xyloglucan for the Treatment of Acute Gastroenteritis in Children: Results of a Randomized, Controlled, Clinical Trial. Gastroenterol. Res. Pract. 2016, 2016, 6874207. [Google Scholar] [CrossRef] [PubMed]
- Pique, N.; Gomez-Guillen, M.D.C.; Montero, M.P. Xyloglucan, a Plant Polymer with Barrier Protective Properties over the Mucous Membranes: An Overview. Int. J. Mol. Sci. 2018, 19, 673. [Google Scholar] [CrossRef] [PubMed]
- Fraile, B.; Alcover, J.; Royuela, M.; Rodriguez, D.; Chaves, C.; Palacios, R.; Pique, N. Xyloglucan, hibiscus and propolis for the prevention of urinary tract infections: Results of in vitro studies. Future Microbiol. 2017, 12, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Dong, M.; Guo, M.; Wang, X.; Zhou, J.; Lei, J.; Guo, C.; Qin, C. Agar/gelatin bilayer gel matrix fabricated by simple thermo-responsive sol-gel transition method. Mater. Sci. Eng. C Mater. Biol. Appl. 2017, 77, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Zhao, J.; Hu, F.; Zheng, Y.; Yang, H.; Pan, S.; Shi, S.; Chen, X.; Wang, S. Design of injectable agar-based composite hydrogel for multi-mode tumor therapy. Carbohydr. Polym. 2018, 180, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, F.; Scaldaferri, F.; Riccioni, M.E.; Casagranda, I.; Forte, E.; Gerardi, V.; Cordischi, C.; Antonini, S.; Tortora, A.; Di Rienzo, T.; et al. Management of acute dyarrhea: Current and future trends. Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 2065–2069. [Google Scholar] [PubMed]
- Ruszczynski, M.; Urbanska, M.; Szajewska, H. Gelatin tannate for treating acute gastroenteritis: A systematic review. Ann. Gastroenterol. 2014, 27, 121–124. [Google Scholar] [PubMed]
- Alexea, O.; Bacarea, V.; Pique, N. The combination of oligo- and polysaccharides and reticulated protein for the control of symptoms in patients with irritable bowel syndrome: Results of a randomised, placebo-controlled, double-blind, parallel group, multicentre clinical trial. United Eur. Gastroenterol. J. 2016, 4, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.D.; Falkow, S. Salmonellosis: Host immune responses and bacterial virulence determinants. Annu. Rev. Immunol. 1996, 14, 533–561. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.; Howlader, D.R.; Mukherjee, P.; Rai, S.; Nag, D.; Koley, H. Retinoic acid decreases the severity of Salmonella enterica serovar Typhimurium mediated gastroenteritis in a mouse model. Immunobiology 2016, 221, 839–844. [Google Scholar] [CrossRef] [PubMed]
- Barthel, M.; Hapfelmeier, S.; Quintanilla-Martinez, L.; Kremer, M.; Rohde, M.; Hogardt, M.; Pfeffer, K.; Russmann, H.; Hardt, W.D. Pretreatment of mice with streptomycin provides a Salmonella enterica serovar Typhimurium colitis model that allows analysis of both pathogen and host. Infect. Immun. 2003, 71, 2839–2858. [Google Scholar] [CrossRef] [PubMed]
- Finlay, B.B.; Falkow, S. Salmonella interactions with polarized human intestinal Caco-2 epithelial cells. J. Infect. Dis. 1990, 162, 1096–1106. [Google Scholar] [CrossRef] [PubMed]
- Jepson, M.A.; Collares-Buzato, C.B.; Clark, M.A.; Hirst, B.H.; Simmons, N.L. Rapid disruption of epithelial barrier function by Salmonella typhimurium is associated with structural modification of intercellular junctions. Infect. Immun. 1995, 63, 356–359. [Google Scholar] [PubMed]
- Tafazoli, F.; Magnusson, K.E.; Zheng, L. Disruption of epithelial barrier integrity by Salmonella enterica serovar typhimurium requires geranylgeranylated proteins. Infect. Immun. 2003, 71, 872–881. [Google Scholar]
- Fasano, A. Toxins and the gut: Role in human disease. Gut 2002, 50 (Suppl. 3), III9–III14. [Google Scholar] [CrossRef] [PubMed]
- Hecht, G. Microbes and microbial toxins: Paradigms for microbial-mucosal interactions. VII. Enteropathogenic Escherichia coli: Physiological alterations from an extracellular position. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 281, G1–G7. [Google Scholar] [CrossRef] [PubMed]
- Tzipori, S.; Hayes, J.; Sims, L.; Withers, M. Streptococcus durans: An unexpected enteropathogen of foals. J. Infect. Dis. 1984, 150, 589–593. [Google Scholar] [CrossRef] [PubMed]
- Hoover, D.; Bendele, S.A.; Wightman, S.R.; Thompson, C.Z.; Hoyt, J.A. Streptococcal enteropathy in infant rats. Lab. Anim. Sci. 1985, 35, 635–641. [Google Scholar] [PubMed]
- Boopathy, V.; Periyasamy, S.; Alexander, T.; Balasubramanian, P. Typhoid fever with caecal ulcer bleed: Managed conservatively. BMJ Case Rep. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Ten Bruggencate, S.J.; Bovee-Oudenhoven, I.M.; Lettink-Wissink, M.L.; Katan, M.B.; Van Der Meer, R. Dietary fructo-oligosaccharides and inulin decrease resistance of rats to salmonella: Protective role of calcium. Gut 2004, 53, 530–535. [Google Scholar] [CrossRef] [PubMed]
- Bovee-Oudenhoven, I.M.; Termont, D.S.; Heidt, P.J.; Van der Meer, R. Increasing the intestinal resistance of rats to the invasive pathogen Salmonella enteritidis: Additive effects of dietary lactulose and calcium. Gut 1997, 40, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Mullane, K.M.; Kraemer, R.; Smith, B. Myeloperoxidase activity as a quantitative assessment of neutrophil infiltration into ischemic myocardium. J. Pharmacol. Methods 1985, 14, 157–167. [Google Scholar] [CrossRef]
- Esposito, E.; Impellizzeri, D.; Mazzon, E.; Paterniti, I.; Cuzzocrea, S. Neuroprotective activities of palmitoylethanolamide in an animal model of Parkinson’s disease. PLoS ONE 2012, 7, e41880. [Google Scholar] [CrossRef] [PubMed]









© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esposito, E.; Campolo, M.; Casili, G.; Lanza, M.; Franco, D.; Filippone, A.; Peritore, A.F.; Cuzzocrea, S. Protective Effects of Xyloglucan in Association with the Polysaccharide Gelose in an Experimental Model of Gastroenteritis and Urinary Tract Infections. Int. J. Mol. Sci. 2018, 19, 1844. https://doi.org/10.3390/ijms19071844
Esposito E, Campolo M, Casili G, Lanza M, Franco D, Filippone A, Peritore AF, Cuzzocrea S. Protective Effects of Xyloglucan in Association with the Polysaccharide Gelose in an Experimental Model of Gastroenteritis and Urinary Tract Infections. International Journal of Molecular Sciences. 2018; 19(7):1844. https://doi.org/10.3390/ijms19071844
Chicago/Turabian StyleEsposito, Emanuela, Michela Campolo, Giovanna Casili, Marika Lanza, Domenico Franco, Alessia Filippone, Alessio F. Peritore, and Salvatore Cuzzocrea. 2018. "Protective Effects of Xyloglucan in Association with the Polysaccharide Gelose in an Experimental Model of Gastroenteritis and Urinary Tract Infections" International Journal of Molecular Sciences 19, no. 7: 1844. https://doi.org/10.3390/ijms19071844