Polysialic Acid in Human Plasma Can Compensate the Cytotoxicity of Histones

 ,
,  , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
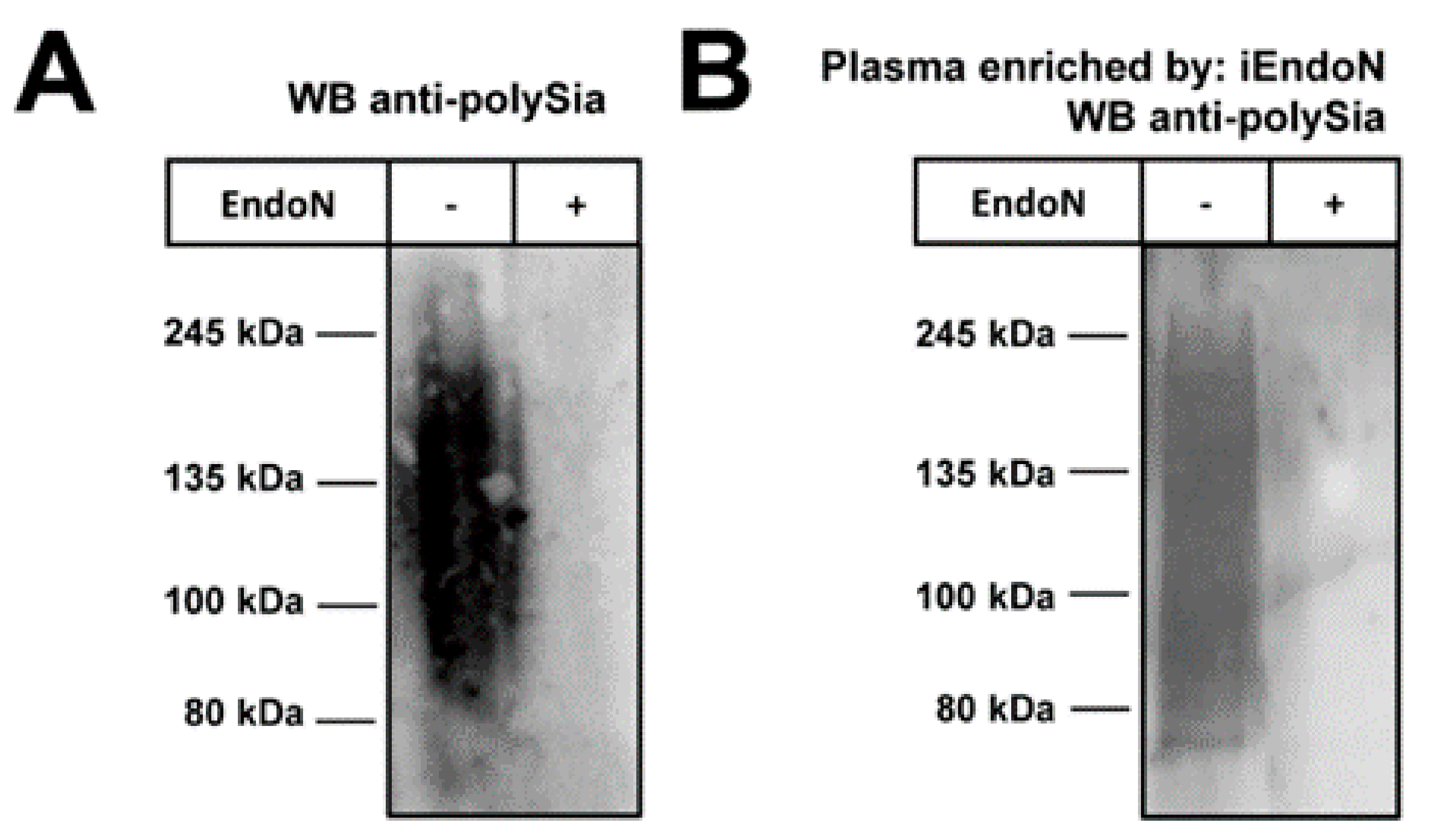
2.1. Plasma Contains Polysialic Acid
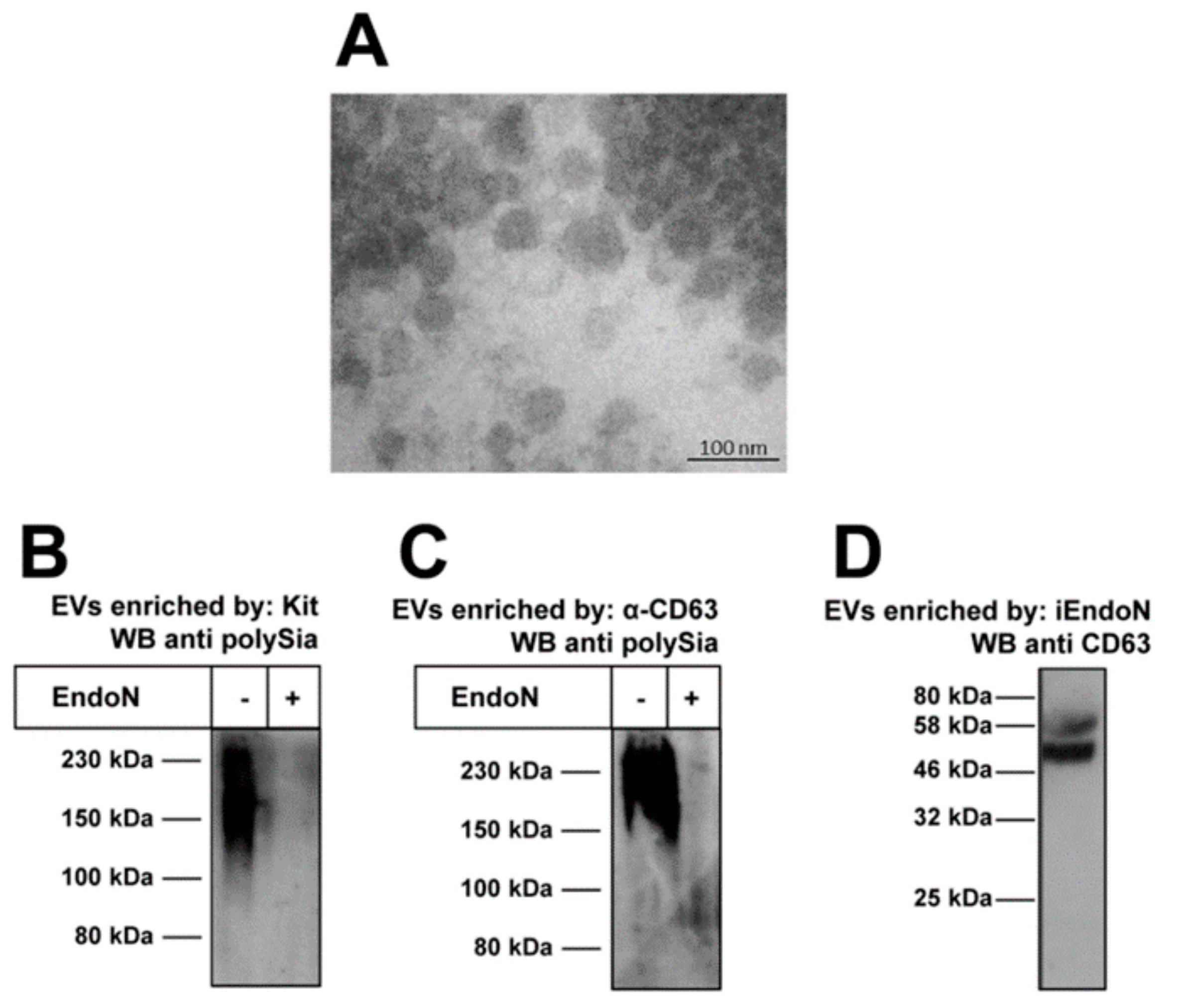
2.2. Extacellular Vesicles Can Contain PolySia
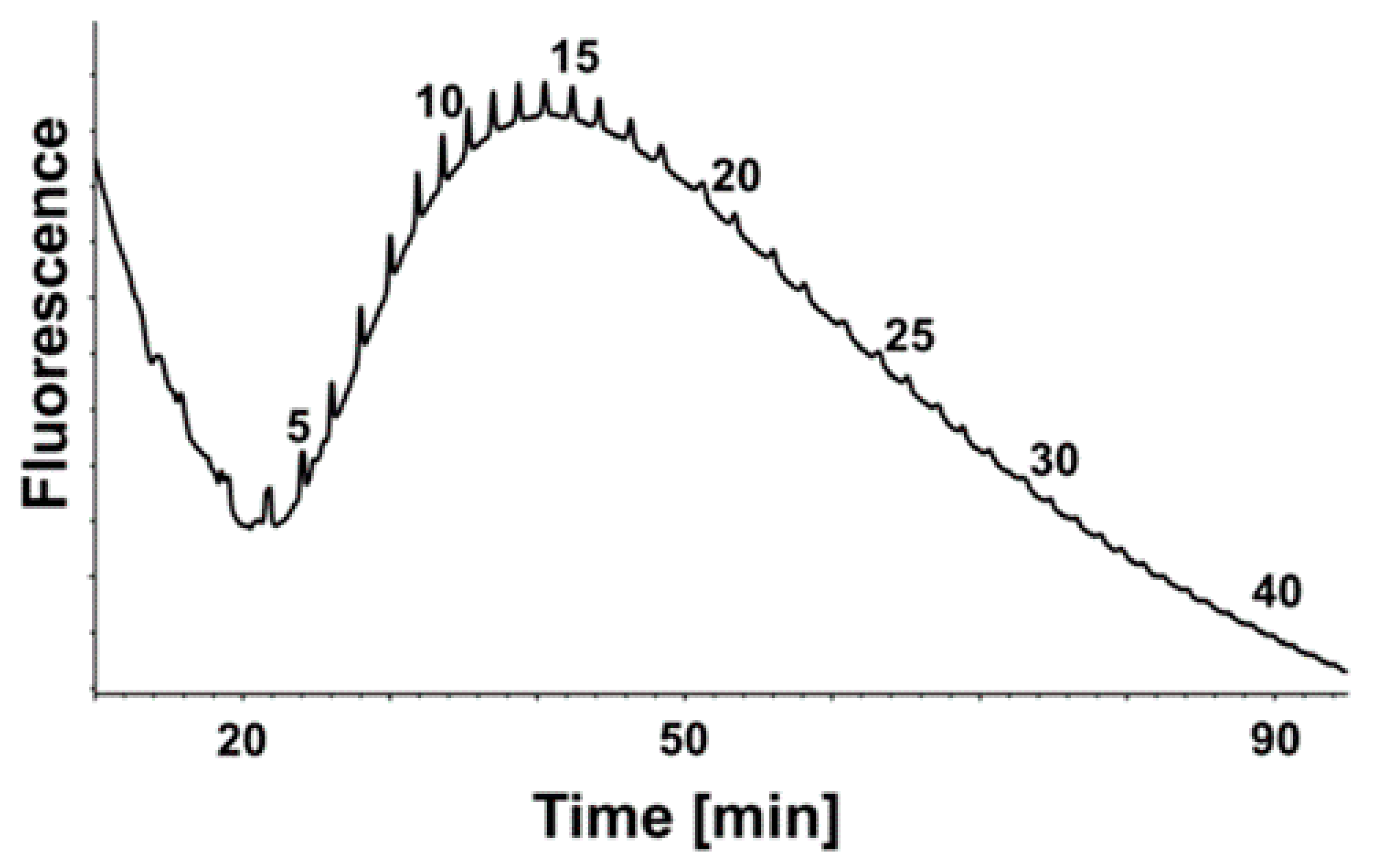
2.3. Chain Length Determination of PolySia from Plasma
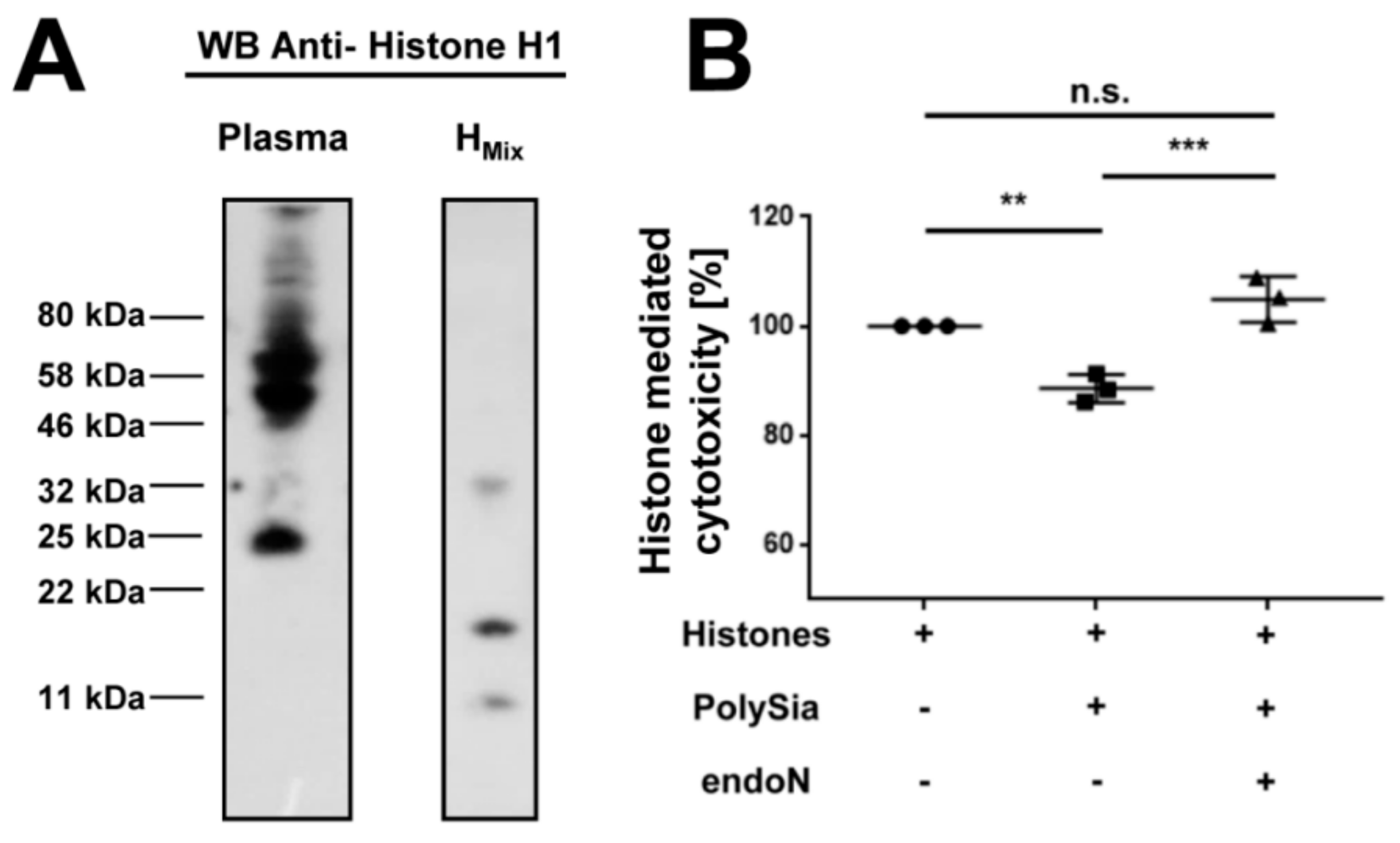
2.4. Histone H1 Is a Natural Binding Partner of PolySia in Human Plasma
2.5. PolySia in Plasma Can Inhibit the Cytotoxicity of Histones
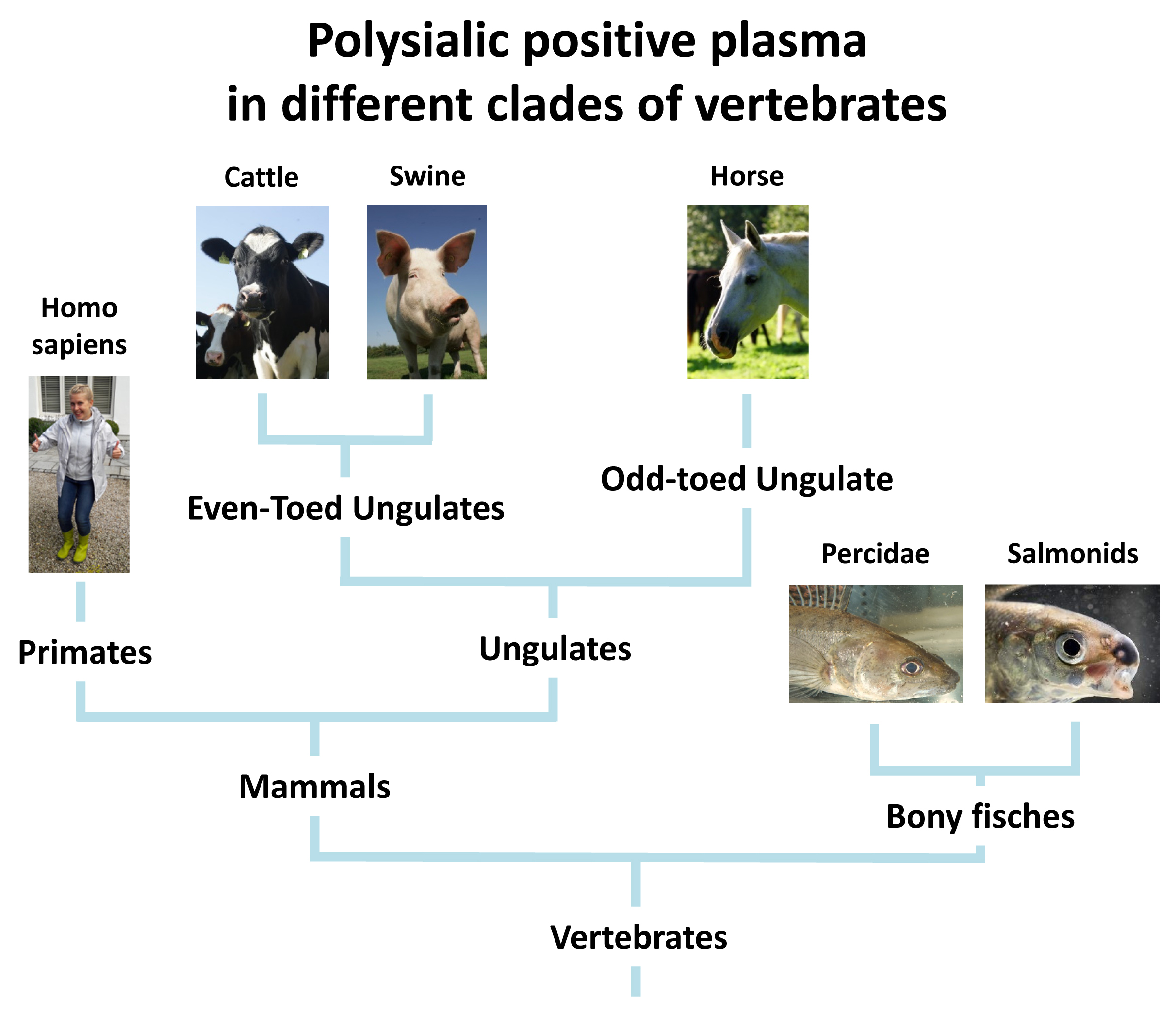
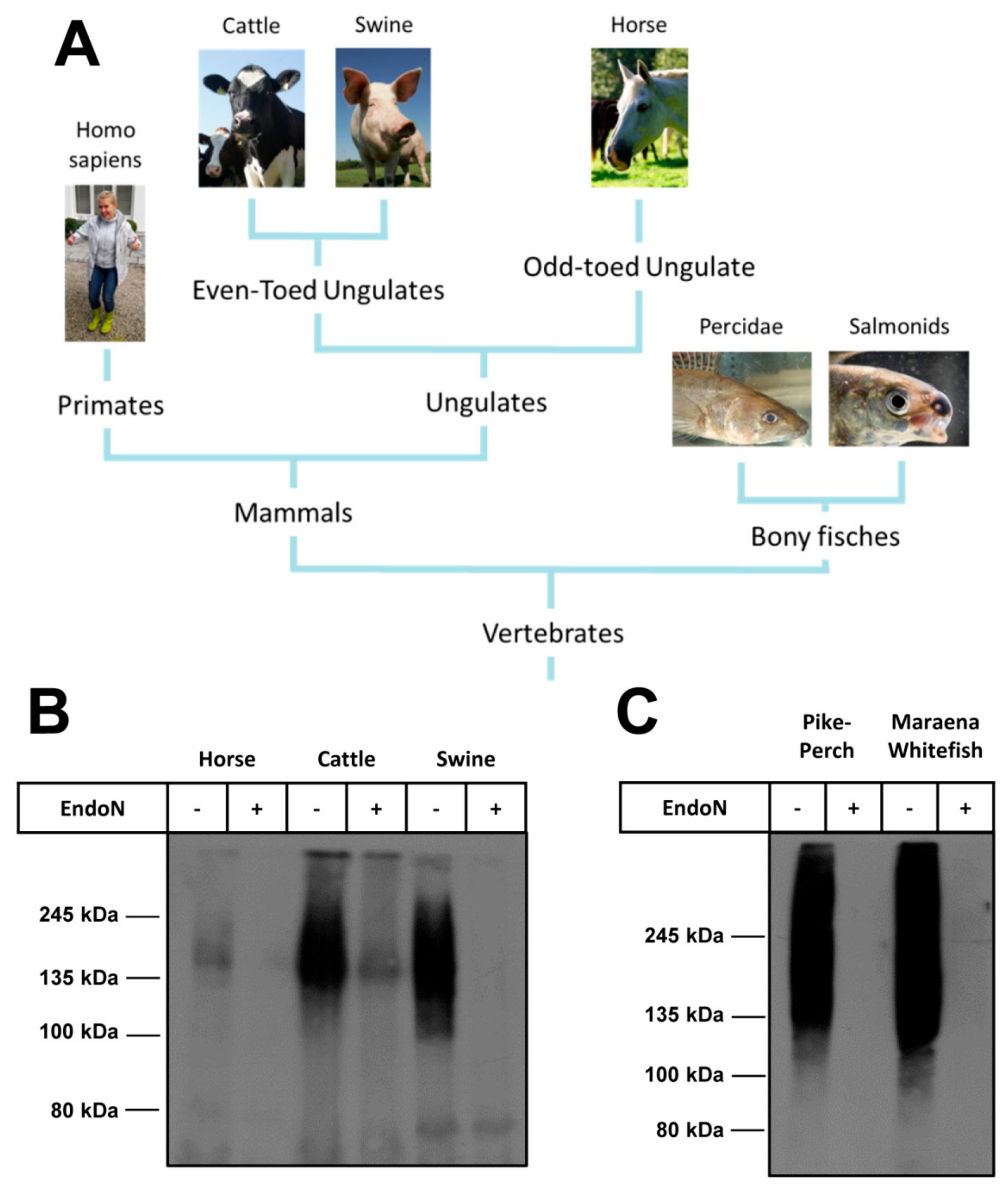
2.6. Several Branches of Vertebrates Have PolySia in Plasma
3. Materials and Methods
3.1. Materials
3.2. Enrichment of Polysialylated EVs and Western Blot Analysis
3.3. Transmission Electron Microscopy
3.4. Determination of Chain Length Distribution by HPLC
3.5. Cell Culture Experiments
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DMB | 4,5-methylene dioxybenzene |
| DP | degree of polymerization |
| endoN | endoneuraminidase |
| EVs | extracellular vesicles |
| LDH | lactate dehydrogenase |
| mAb | monoclonal antibody |
| NETs | neutrophil extracellular traps |
| PolySia | polysialic acid |
| TEM | transmission electron microscopy |
References
- Sato, C.; Kitajima, K. Disialic, oligosialic and polysialic acids: Distribution, functions and related disease. J. Biochem. 2013, 154, 115–136. [Google Scholar] [CrossRef] [PubMed]
- Colley, K.J.; Kitajima, K.; Sato, C. Polysialic acid: Biosynthesis, novel functions and applications. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 498–532. [Google Scholar] [CrossRef] [PubMed]
- Gascon, E.; Vutskits, L.; Kiss, J.Z. The role of psa-ncam in adult neurogenesis. Adv. Exp. Med. Biol. 2010, 663, 127–136. [Google Scholar] [PubMed]
- Rutishauser, U. Polysialic acid in the plasticity of the developing and adult vertebrate nervous system. Nat. Rev. Neurosci. 2008, 9, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Schnaar, R.L.; Gerardy-Schahn, R.; Hildebrandt, H. Sialic acids in the brain: Gangliosides and polysialic acid in nervous system development, stability, disease, and regeneration. Physiol. Rev. 2014, 94, 461–518. [Google Scholar] [CrossRef] [PubMed]
- Galuska, C.E.; Lütteke, T.; Galuska, S.P. Is polysialylated ncam not only a regulator during brain development but also during the formation of other organs? Biology 2017, 6, 27. [Google Scholar] [CrossRef] [PubMed]
- Husmann, M.; Pietsch, T.; Fleischer, B.; Weisgerber, C.; Bitter-Suermann, D. Embryonic neural cell adhesion molecules on human natural killer cells. Eur. J. Immunol. 1989, 19, 1761–1763. [Google Scholar] [CrossRef] [PubMed]
- Kiermaier, E.; Moussion, C.; Veldkamp, C.T.; Gerardy-Schahn, R.; Vries, I.D.; Williams, L.G.; Chaffee, G.R.; Phillips, A.J.; Freiberger, F.; Imre, R.; et al. Polysialylation controls dendritic cell trafficking by regulating chemokine recognition. Science 2016, 351, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Ermert, D.; Urban, C.F.; Laube, B.; Goosmann, C.; Zychlinsky, A.; Brinkmann, V. Mouse neutrophil extracellular traps in microbial infections. J. Innate Immun. 2009, 1, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, T.A.; Abed, U.; Goosmann, C.; Hurwitz, R.; Schulze, I.; Wahn, V.; Weinrauch, Y.; Brinkmann, V.; Zychlinsky, A. Novel cell death program leads to neutrophil extracellular traps. J. Cell Biol. 2007, 176, 231–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillipson, M.; Kubes, P. The neutrophil in vascular inflammation. Nat. Med. 2011, 17, 1381–1390. [Google Scholar] [CrossRef] [PubMed]
- Ma, A.C.; Kubes, P. Platelets, neutrophils, and neutrophil extracellular traps (nets) in sepsis. J. Thromb. Haemost. 2008, 6, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Alghamdi, A.S.; Foster, D.N. Seminal dnase frees spermatozoa entangled in neutrophil extracellular traps. Biol. Reprod. 2005, 73, 1174–1181. [Google Scholar] [CrossRef] [PubMed]
- Saffarzadeh, M.; Juenemann, C.; Queisser, M.A.; Lochnit, G.; Barreto, G.; Galuska, S.P.; Lohmeyer, J.; Preissner, K.T. Neutrophil extracellular traps directly induce epithelial and endothelial cell death: A predominant role of histones. PLoS ONE 2012, 7, e32366. [Google Scholar] [CrossRef] [PubMed]
- Saffarzadeh, M.; Preissner, K.T. Fighting against the dark side of neutrophil extracellular traps in disease: Manoeuvres for host protection. Curr. Opin. Hematol. 2013, 20, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Wildhagen, K.C.; Garcia de Frutos, P.; Reutelingsperger, C.P.; Schrijver, R.; Areste, C.; Ortega-Gomez, A.; Deckers, N.M.; Hemker, H.C.; Soehnlein, O.; Nicolaes, G.A. Non-anticoagulant heparin prevents histone-mediated cytotoxicity in vitro and improves survival in sepsis. Blood 2013, 123, 1098–1101. [Google Scholar] [CrossRef] [PubMed]
- Zlatina, K.; Lütteke, T.; Galuska, S. Individual impact of distinct polysialic acid chain lengths on the cytotoxicity of histone h1, h2a, h2b, h3 and h4. Polymers 2017, 9, 720. [Google Scholar] [CrossRef]
- Galuska, C.E.; Dambon, J.A.; Kühnle, A.; Bornhöfft, K.F.; Prem, G.; Zlatina, K.; Lütteke, T.; Galuska, S.P. Artificial polysialic acid chains as sialidase-resistant molecular-anchors to accumulate particles on neutrophil extracellular traps. Front. Immunol. 2017, 8, 1229. [Google Scholar] [CrossRef] [PubMed]
- Simon, P.; Bäumner, S.; Busch, O.; Röhrich, R.; Kaese, M.; Richterich, P.; Wehrend, A.; Müller, K.; Gerardy-Schahn, R.; Mühlenhoff, M.; et al. Polysialic acid is present in mammalian semen as a post-translational modification of the neural cell adhesion molecule ncam and the polysialyltransferase st8siaii. J. Biol. Chem. 2013, 288, 18825–18833. [Google Scholar] [CrossRef] [PubMed]
- Ulm, C.; Saffarzadeh, M.; Mahavadi, P.; Müller, S.; Prem, G.; Saboor, F.; Simon, P.; Middendorff, R.; Geyer, H.; Henneke, I.; et al. Soluble polysialylated ncam: A novel player of the innate immune system in the lung. Cell Mol. Life Sci. 2013, 70, 3695–3708. [Google Scholar] [CrossRef] [PubMed]
- Glüer, S.; Schelp, C.; Madry, N.; Schweinitz, D.; Eckhardt, M.; Gerardy-Schahn, R. Serum polysialylated neural cell adhesion molecule in childhood neuroblastoma. Br. J. Cancer 1998, 78, 106–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glüer, S.; Wunder, M.A.; Schelp, C.; Radtke, E.; Gerardy-Schahn, R. Polysialylated neural cell adhesion molecule serum levels in normal children. Pediatr. Res. 1998, 44, 915–919. [Google Scholar] [CrossRef] [PubMed]
- Piras, F.; Schiff, M.; Chiapponi, C.; Bossù, P.; Mühlenhoff, M.; Caltagirone, C.; Gerardy-Schahn, R.; Hildebrandt, H.; Spalletta, G. Brain structure, cognition and negative symptoms in schizophrenia are associated with serum levels of polysialic acid-modified ncam. Transl. Psychiatry 2015, 5, e658. [Google Scholar] [CrossRef] [PubMed]
- Jakobsson, E.; Jokilammi, A.; Aalto, J.; Ollikka, P.; Lehtonen, J.V.; Hirvonen, H.; Finne, J. Identification of amino acid residues at the active site of endosialidase that dissociate the polysialic acid binding and cleaving activities in escherichia coli k1 bacteriophages. Biochem. J. 2007, 405, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Buzas, E.I.; György, B.; Nagy, G.; Falus, A.; Gay, S. Emerging role of extracellular vesicles in inflammatory diseases. Nat. Rev. Rheumatol. 2014, 10, 356–364. [Google Scholar] [CrossRef] [PubMed]
- Kalra, H.; Simpson, R.J.; Ji, H.; Aikawa, E.; Altevogt, P.; Askenase, P.; Bond, V.C.; Borràs, F.E.; Breakefield, X.; Budnik, V.; et al. Vesiclepedia: A compendium for extracellular vesicles with continuous community annotation. PLoS Biol. 2012, 10, e1001450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Pol, E.; Böing, A.N.; Harrison, P.; Sturk, A.; Nieuwland, R. Classification, functions, and clinical relevance of extracellular vesicles. Pharmacol. Rev. 2012, 64, 676–705. [Google Scholar] [CrossRef] [PubMed]
- Silk, E.; Zhao, H.; Weng, H.; Ma, D. The role of extracellular histone in organ injury. Cell Death Dis. 2017, 8, e2812. [Google Scholar] [CrossRef] [PubMed]
- Dhaenens, M.; Glibert, P.; Meert, P.; Vossaert, L.; Deforce, D. Histone proteolysis: A proposal for categorization into ‘clipping’ and ‘degradation’. Bioessays 2015, 37, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Thorslund, T.; Ripplinger, A.; Hoffmann, S.; Wild, T.; Uckelmann, M.; Villumsen, B.; Narita, T.; Sixma, T.K.; Choudhary, C.; Bekker-Jensen, S.; et al. Histone h1 couples initiation and amplification of ubiquitin signalling after DNA damage. Nature 2015, 527, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Palic, D.; Ostojic, J.; Andreasen, C.B.; Roth, J.A. Fish cast nets: Neutrophil extracellular traps are released from fish neutrophils. Dev. Comp. Immunol. 2007, 31, 805–816. [Google Scholar] [CrossRef] [PubMed]
- Cedervall, J.; Hamidi, A.; Olsson, A.K. Platelets, nets and cancer. Thromb. Res. 2018, 164, S148–S152. [Google Scholar] [CrossRef] [PubMed]
- Frosch, M.; Görgen, I.; Boulnois, G.J.; Timmis, K.N.; Bitter-Suermann, D. Nzb mouse system for production of monoclonal antibodies to weak bacterial antigens: Isolation of an igg antibody to the polysaccharide capsules of escherichia coli k1 and group b meningococci. Proc. Natl. Acad. Sci. USA 1985, 82, 1194–1198. [Google Scholar] [CrossRef] [PubMed]
- Stummeyer, K.; Dickmanns, A.; Mühlenhoff, M.; Gerardy-Schahn, R.; Ficner, R. Crystal structure of the polysialic acid-degrading endosialidase of bacteriophage k1f. Nat. Struct. Mol. Biol. 2005, 12, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Schägger, H.; Jagow, G.V. Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kda. Anal. Biochem. 1987, 166, 368–379. [Google Scholar] [CrossRef]
- Nakata, D.; Troy, F.A. Degree of polymerization (dp) of polysialic acid (polysia) on neural cell adhesion molecules (n-cams): Development and application of a new strategy to accurately determine the dp of polysia chains on n-cams. J. Biol. Chem. 2005, 280, 38305–38316. [Google Scholar] [CrossRef] [PubMed]
- Galuska, S.P.; Geyer, H.; Bleckmann, C.; Röhrich, R.C.; Maass, K.; Bergfeld, A.K.; Mühlenhoff, M.; Geyer, R. Mass spectrometric fragmentation analysis of oligosialic and polysialic acids. Anal. Chem. 2010, 82, 2059–2066. [Google Scholar] [CrossRef] [PubMed]
- Galuska, S.P.; Galuska, C.E.; Tharmalingam, T.; Zlatina, K.; Prem, G.; Husejnov, F.C.O.; Rudd, P.M.; Vann, W.F.; Reid, C.; Vionnet, J.; et al. In vitro generation of polysialylated cervical mucins by bacterial polysialyltransferases to counteract cytotoxicity of extracellular histones. FEBS J. 2017, 284, 1688–1699. [Google Scholar] [CrossRef] [PubMed]
- Inoue, S.; Inoue, Y. Ultrasensitive analysis of sialic acids and oligo/polysialic acids by fluorometric high-performance liquid chromatography. In Recognition of Carbohydrates in Biological Systems; Lee, Y.C., Lee, R.T., Eds.; Academic Press: San Diego, CA, USA; London, UK, 2003; Volume 362, pp. 543–560. [Google Scholar]
- Galuska, S.P.; Geyer, H.; Weinhold, B.; Kontou, M.; Röhrich, R.C.; Bernard, U.; Gerardy-Schahn, R.; Reutter, W.; Münster-Kühnel, A.; Geyer, R. Quantification of nucleotide-activated sialic acids by a combination of reduction and fluorescent labeling. Anal. Chem. 2010, 82, 4591–4598. [Google Scholar] [CrossRef] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zlatina, K.; Saftenberger, M.; Kühnle, A.; Galuska, C.E.; Gärtner, U.; Rebl, A.; Oster, M.; Vernunft, A.; Galuska, S.P. Polysialic Acid in Human Plasma Can Compensate the Cytotoxicity of Histones. Int. J. Mol. Sci. 2018, 19, 1679. https://doi.org/10.3390/ijms19061679
Zlatina K, Saftenberger M, Kühnle A, Galuska CE, Gärtner U, Rebl A, Oster M, Vernunft A, Galuska SP. Polysialic Acid in Human Plasma Can Compensate the Cytotoxicity of Histones. International Journal of Molecular Sciences. 2018; 19(6):1679. https://doi.org/10.3390/ijms19061679
Chicago/Turabian StyleZlatina, Kristina, Max Saftenberger, Andrea Kühnle, Christina E. Galuska, Ulrich Gärtner, Alexander Rebl, Michael Oster, Andreas Vernunft, and Sebastian P. Galuska. 2018. "Polysialic Acid in Human Plasma Can Compensate the Cytotoxicity of Histones" International Journal of Molecular Sciences 19, no. 6: 1679. https://doi.org/10.3390/ijms19061679
APA StyleZlatina, K., Saftenberger, M., Kühnle, A., Galuska, C. E., Gärtner, U., Rebl, A., Oster, M., Vernunft, A., & Galuska, S. P. (2018). Polysialic Acid in Human Plasma Can Compensate the Cytotoxicity of Histones. International Journal of Molecular Sciences, 19(6), 1679. https://doi.org/10.3390/ijms19061679