Isoviolanthin Extracted from Dendrobium officinale Reverses TGF-β1-Mediated Epithelial–Mesenchymal Transition in Hepatocellular Carcinoma Cells via Deactivating the TGF-β/Smad and PI3K/Akt/mTOR Signaling Pathways


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
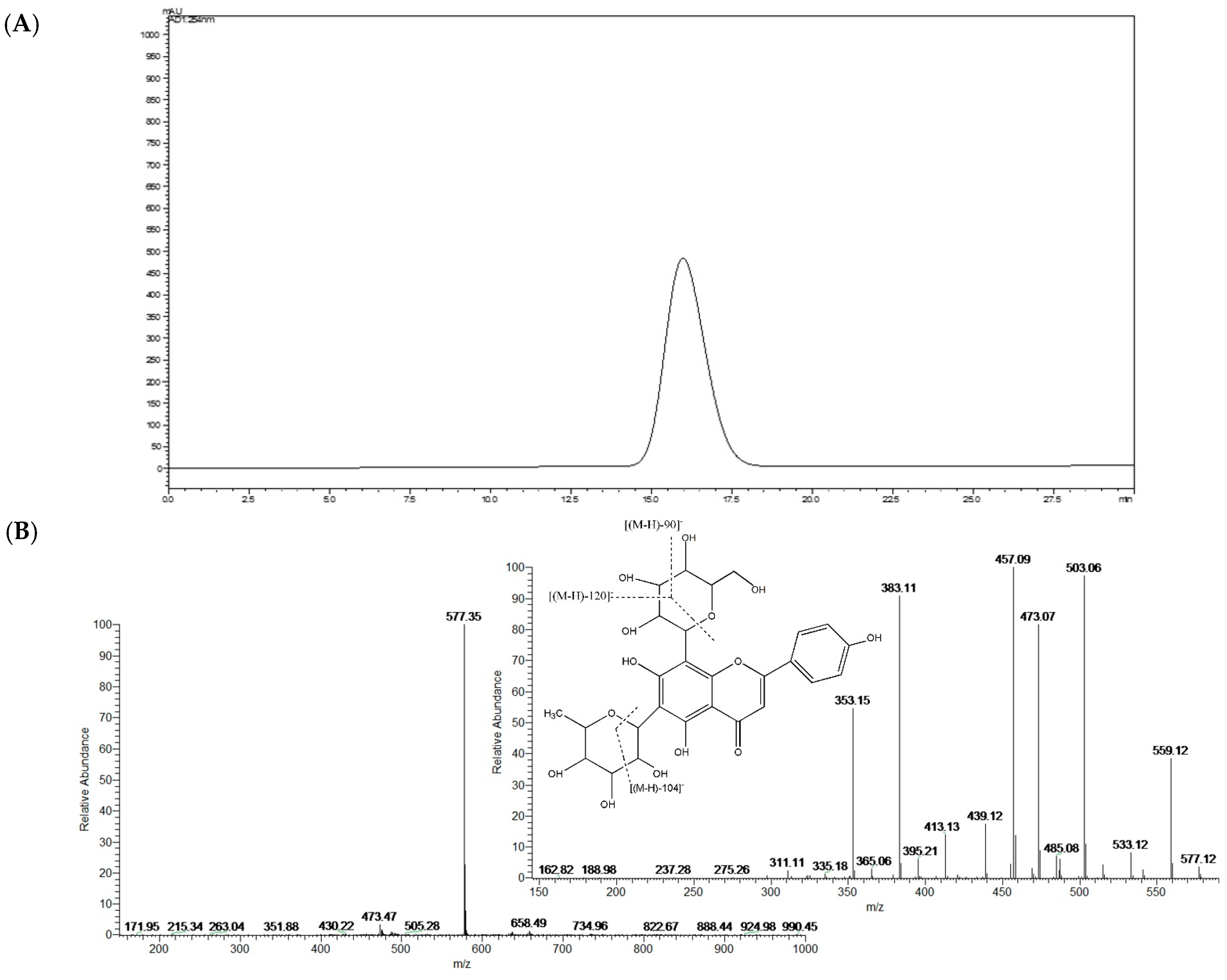
2.1. Structural Identification of Isolated Isoviolanthin
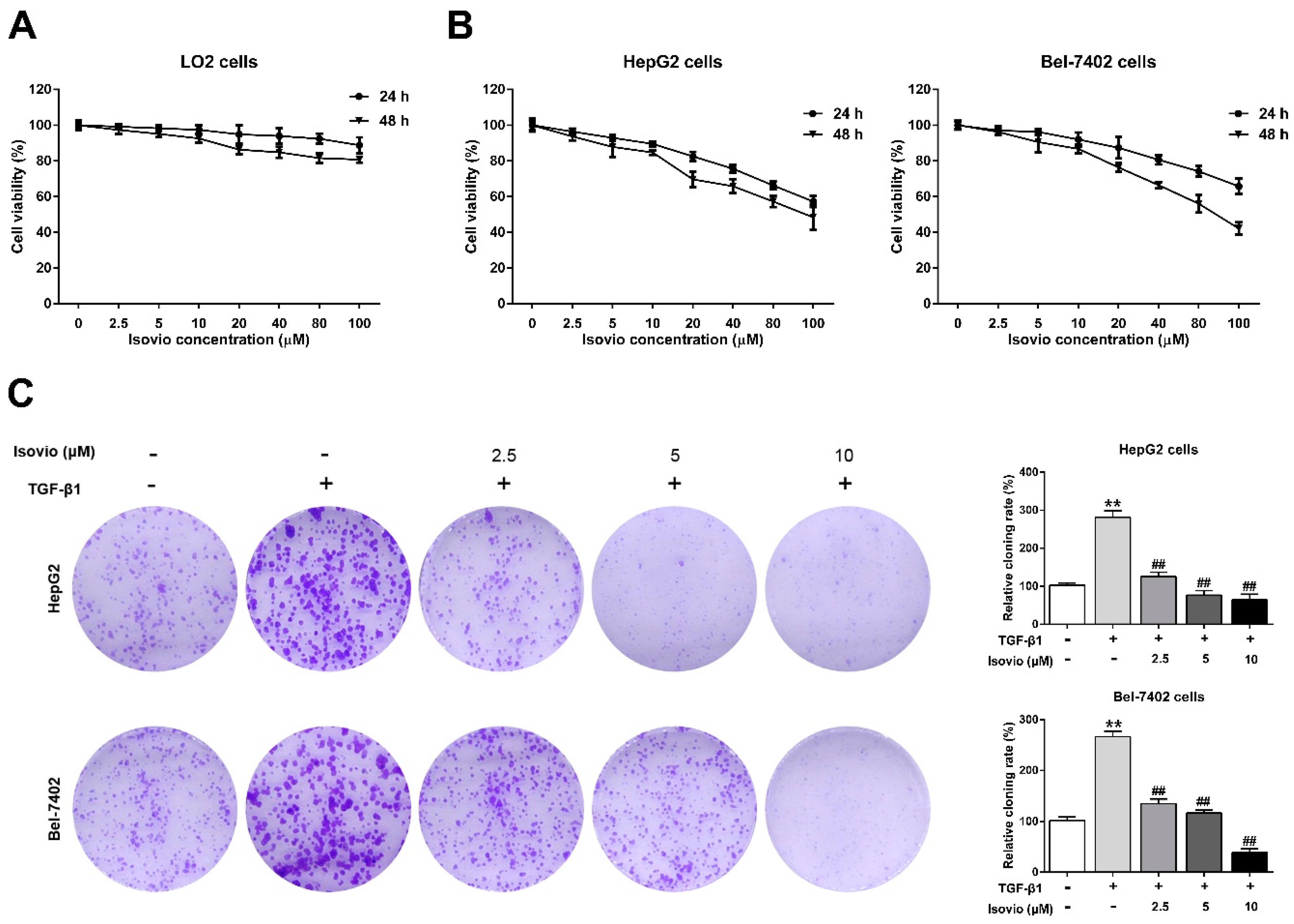
2.2. The Effect of Isoviolanthin on the Growth of Hepatocellular Carcinoma (HCC) and LO2 Cells
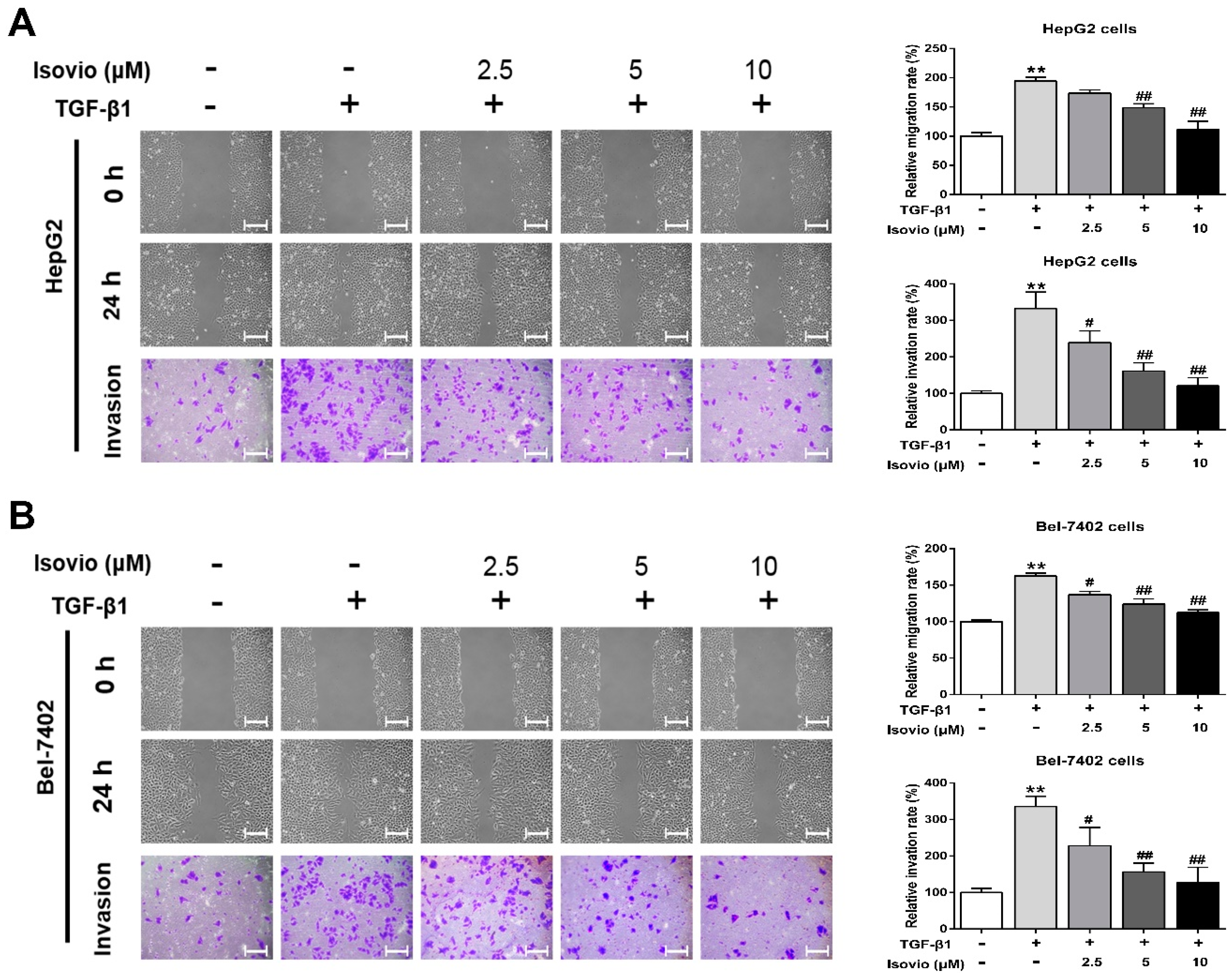
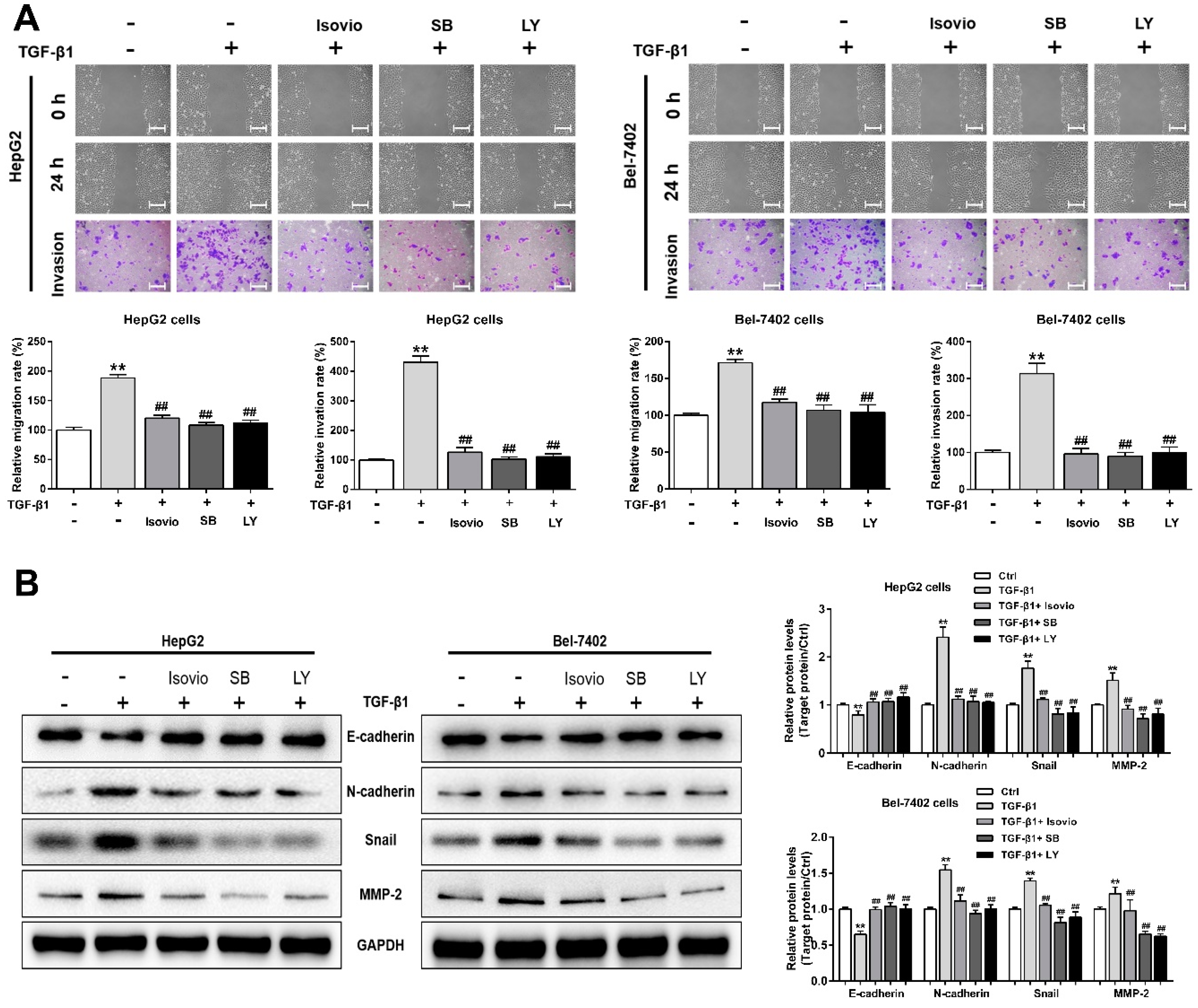
2.3. Isoviolanthin Inhibited the Transforming Growth Factor (TGF)-β1-Induced Migration and Invasion of HCC Cells
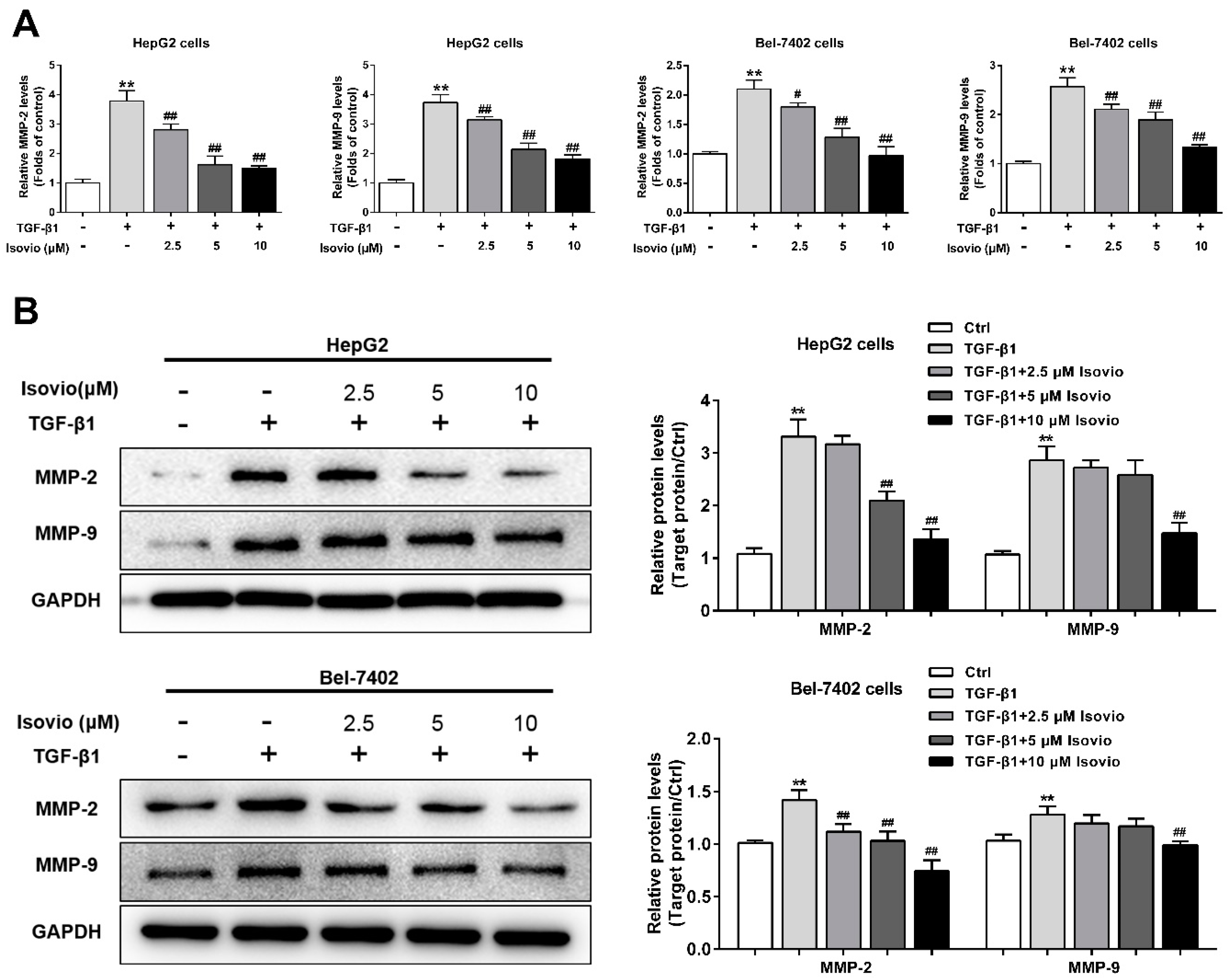
2.4. Isoviolanthin Inhibited TGF-β1-Induced MMP-2 and -9 in HCC Cells
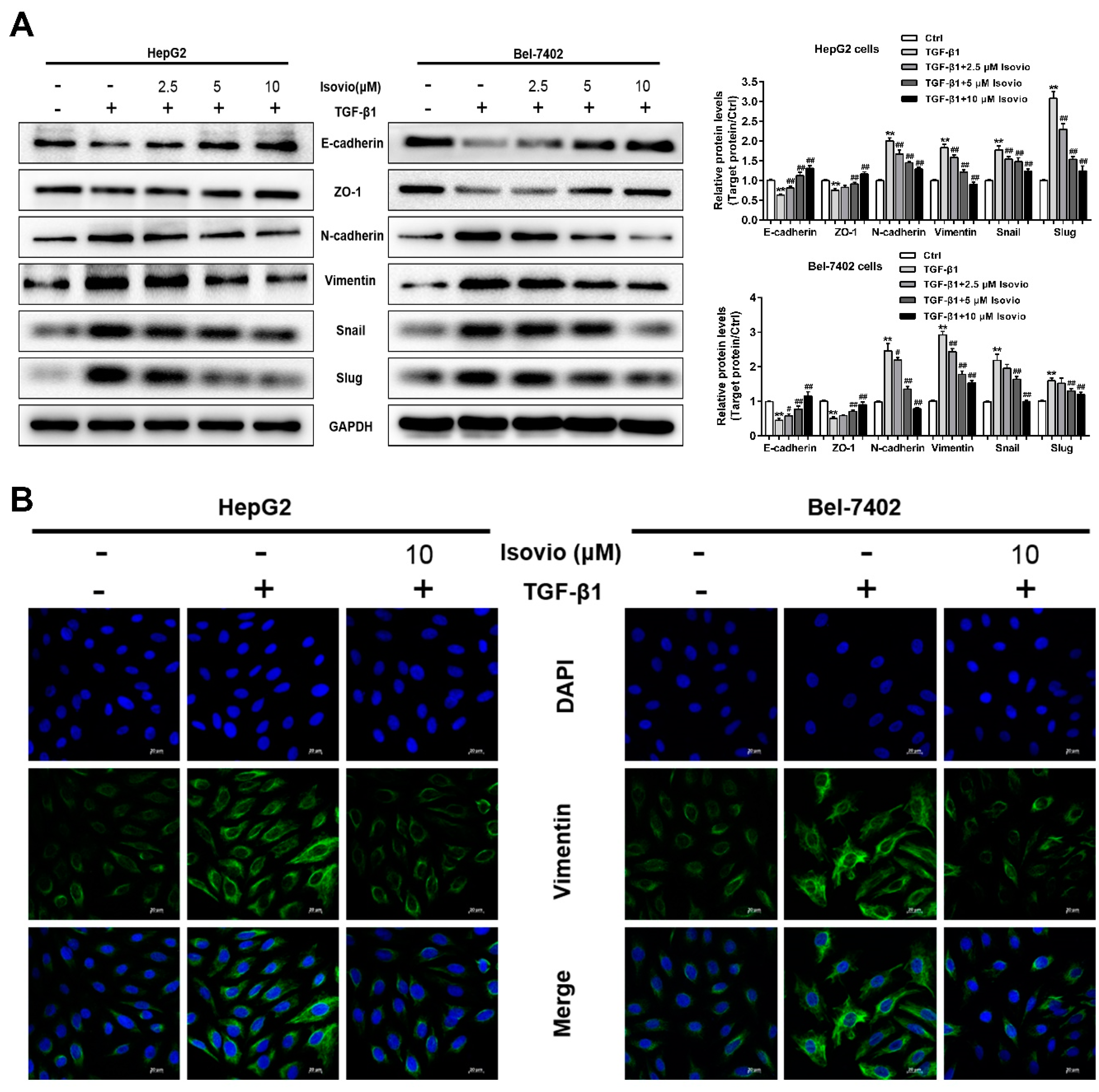
2.5. Isoviolanthin Reversed the Induction of EMT Biomarkers by TGF-β1 in HCC Cells
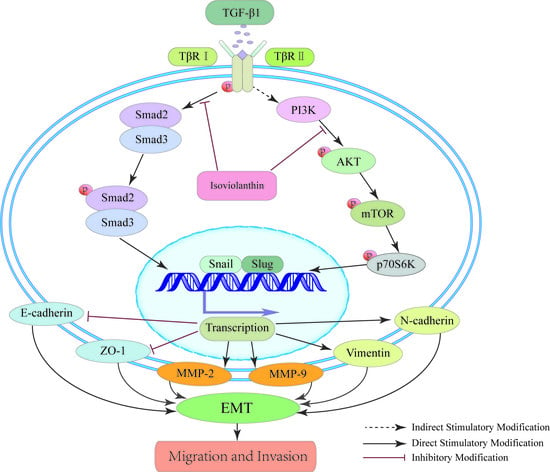
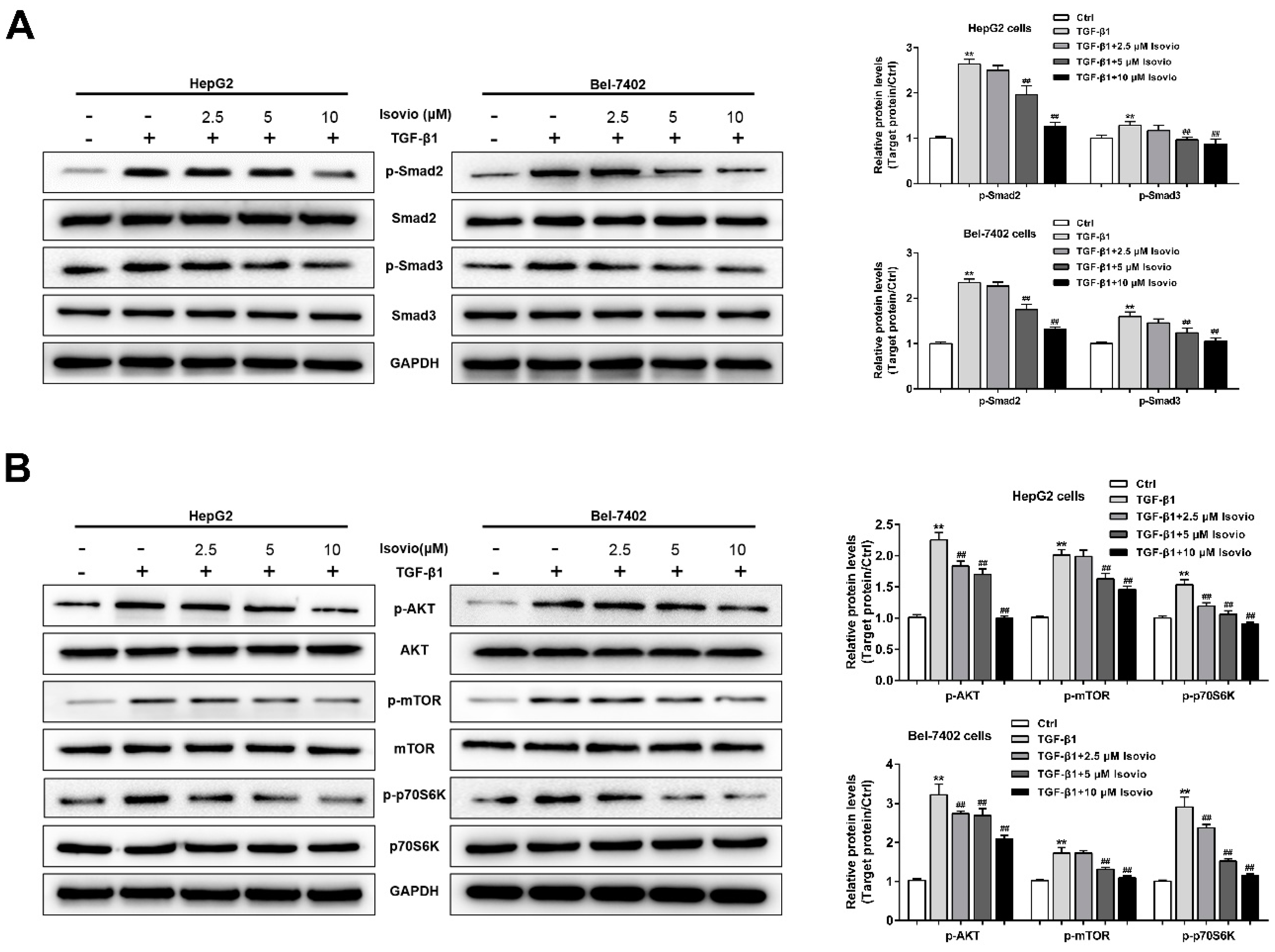
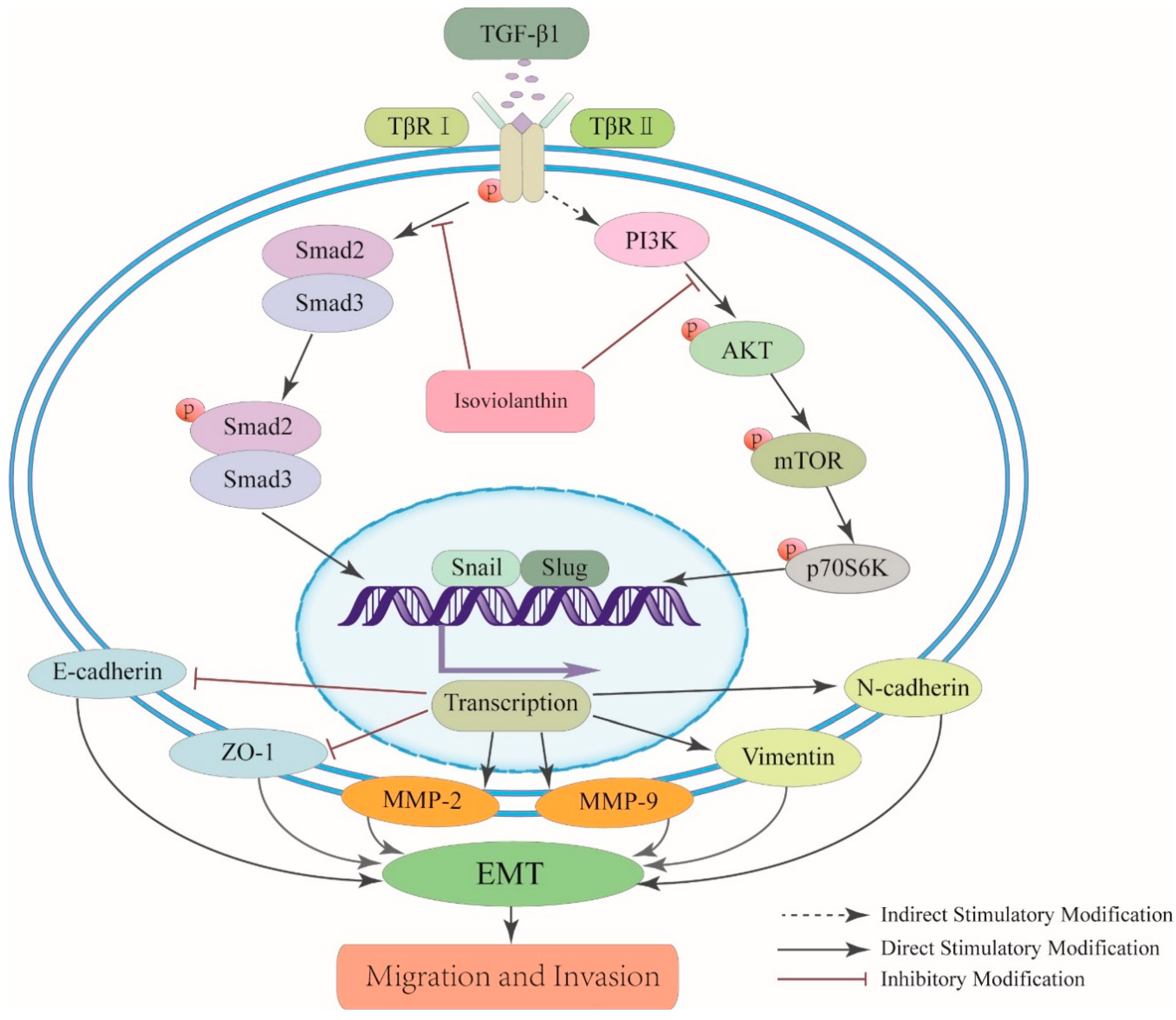
2.6. Isoviolanthin Inhibited TGF-β1-Induced EMT in HCC Cells via Regulating the TGF-β/Smad and PI3K/Akt/mTOR Pathways
3. Discussion
4. Materials and Methods
4.1. Plant Material and Chemicals
4.2. Extraction of Crude Isoviolanthin
4.3. Purification of Isoviolanthin
4.4. Structural Analysis of Isoviolanthin
4.5. Cell Culture and Treatment
4.6. Influence of Isoviolanthin on Cell Proliferation
4.7. ELISA
4.8. Colony Forming Assay
4.9. Immunofluorescence Analysis
4.10. Wound Healing and Transwell Assays
4.11. Western Blot Analysis
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| UV | Ultraviolet Radiation |
| HPLC | High-Performance Liquid Chromatography |
| Pre-HPLC | Preparative High-Performance Liquid Chromatography |
| LC-MS | Liquid Chromatography-Mass spectrometry |
| NMR | Nuclear Magnetic Resonance |
| HCC | Hepatocellular Carcinoma |
| TGF-β1 | Transforming Growth Factor-β1 |
| EMT | Epithelial–Mesenchymal Transition |
| MMPs | Matrix Metalloproteinases |
| PI3K | Phosphatidylinositol-3-Kinase |
| Akt | Protein Kinase B |
| TGF-β1 | Transforming Growth Factor-β1 |
| P70S6K | p70 Ribosomal Protein S6 Kinase |
| mTOR | Mammalian Target of Rapamycin |
References
- White, D.L.; Thrift, A.P.; Kanwal, F.; Davila, J.; El-Serag, H.B. Incidence of hepatocellular carcinoma in all 50 united states, from 2000 through 2012. Gastroenterology 2017, 152, 812–820. [Google Scholar] [CrossRef] [PubMed]
- Waziry, R.; Grebely, J.; Amin, J.; Alavi, M.; Hajarizadeh, B.; George, J.; Matthews, G.V.; Law, M.; Dore, G.J. Trends in hepatocellular carcinoma among people with HBV or HCV notification in Australia (2000–2014). J. Hepatol. 2016, 65, 1086–1093. [Google Scholar] [CrossRef] [PubMed]
- Bertuccio, P.; Turati, F.; Carioli, G.; Rodriguez, T.; La Vecchia, C.; Malvezzi, M.; Negri, E. Global trends and predictions in hepatocellular carcinoma mortality. J. Hepatol. 2017, 67, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Gray, W.H.; Patel, R.; Baker, C.J.; Starnes, V.A.; Bowdish, M.E. Resection of metastatic hepatocellular carcinoma in the ventricular septum causing left ventricular outflow tract obstruction. Ann. Thorac. Surg. 2018, 105, E107–E108. [Google Scholar] [CrossRef] [PubMed]
- Na, S.K.; Lee, D.; Yim, H.J.; Kim, S.Y.; Suh, S.J.; Hyun, J.J.; Jung, S.W.; Jung, Y.K.; Koo, J.S.; Kim, J.H.; et al. Subclassification of Barcelona clinic liver cancer stage C hepatocellular carcinoma by major vessel invasion and distant metastasis. Gastroenterology 2016, 150, S1128. [Google Scholar] [CrossRef]
- Harding, J.J.; Abu-Zeinah, G.; Chou, J.F.; Owen, D.H.; Ly, M.; Lowery, M.A.; Capanu, M.; Do, R.; Kemeny, N.E.; O’Reilly, E.M.; et al. Frequency, morbidity, and mortality of bone metastases in advanced hepatocellular carcinoma. J. Natl. Compr. Cancer Netw. 2018, 16, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Peroukides, S.; Alexopoulos, A.; Kalofonos, H.; Papadaki, H. The role of epithelial to mesenchymal transition (EMT) in human hepatocellular carcinoma (HCC). Ann. Oncol. 2010, 21, 53. [Google Scholar]
- Giannelli, G.; Koudelkova, P.; Dituri, F.; Mikulits, W. Role of epithelial to mesenchymal transition in hepatocellular carcinoma. J. Hepatol. 2016, 65, 798–808. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.Y.; Hao, M. Metformin inhibits TGF-β1-induced epithelial-to-mesenchymal transition via Pkm2 relative-mTOR/p70s6k signaling pathway in cervical carcinoma cells. Int. J. Mol. Sci. 2016, 17, 2000. [Google Scholar] [CrossRef] [PubMed]
- Li, D.P.; Fan, J.; Wu, Y.J.; Xie, Y.F.; Zha, J.M.; Zhou, X.M. Mir-155 up-regulated by TGF- β promotes epithelial-mesenchymal transition, invasion and metastasis of human hepatocellular carcinoma cells in vitro. Am. J. Transl. Res. 2017, 9, 2956–2965. [Google Scholar] [PubMed]
- Zhou, S.J.; Liu, F.Y.; Zhang, A.H.; Liang, H.F.; Wang, Y.; Ma, R.; Jiang, Y.H.; Sun, N.F. Microrna-199b-5p attenuates TGF-β1-induced epithelial-mesenchymal transition in hepatocellular carcinoma. Br. J. Cancer 2017, 117, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Lv, P.; Sun, Z.; Han, L.; Zhou, W. 14-3-3β promotes migration and invasion of human hepatocellular carcinoma cells by modulating expression of MMP2 and MMP9 through PI3K/AKT/NF-κB pathway. PLoS ONE 2016, 11, e0146070. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Ding, F.; Liu, Q.; Yao, Y. Knockdown of macc1 expression suppressed hepatocellular carcinoma cell migration and invasion and inhibited expression of MMP2 and MMP9. Mol. Cell. Biochem. 2013, 376, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Massague, J.; Seoane, J.; Wotton, D. Smad transcription factors. Gene Dev. 2005, 19, 2783–2810. [Google Scholar] [CrossRef] [PubMed]
- Miyazono, K. Transforming growth factor-β signaling in epithelial-mesenchymal transition and progression of cancer. Proc. Jpn. Acad. B Phys. 2009, 85, 314–323. [Google Scholar] [CrossRef]
- Bakin, A.V.; Tomlinson, A.K.; Bhowmick, N.A.; Moses, H.L.; Arteaga, C.L. Phosphatidylinositol 3-kinase function is required for transforming growth factor β-mediated epithelial to mesenchymal transition and cell migration. J. Biol. Chem. 2000, 275, 36803–36810. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Hebert, M.C.; Zhang, Y.E. TGF-β receptor-activated p38 MAP Kinase mediates Smad-independent TGF-β responses. Embo J. 2002, 21, 3749–3759. [Google Scholar] [CrossRef] [PubMed]
- Bhowmick, N.A.; Ghiassi, M.; Bakin, A.; Aakre, M.; Lundquist, C.A.; Engel, M.E.; Arteaga, C.L.; Moses, H.L. Transforming growth factor-β 1 mediates epithelial to mesenchymal transdifferentiation through a Rhoa-dependent mechanism. Mol. Biol. Cell. 2001, 12, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Hamidi, A.; Song, J.; Thakur, N.; Itoh, S.; Marcusson, A.; Bergh, A.; Heldin, C.H.; Landstrom, M. TGF-β promotes PI3K-AKT signaling and prostate cancer cell migration through the traf6-mediated ubiquitylation of p85α. Sci. Signal. 2017, 10, eaal4186. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.H.; Ko, J.H.; Lee, J.H.; Kim, C.; Lee, H.; Nam, D.; Lee, J.; Lee, S.G.; Yang, W.M.; Um, J.Y.; et al. Ginkgolic acid inhibits invasion and migration and TGF-induced EMT of lung cancer cells through PI3K/Akt/mTOR inactivation. J. Cell. Physiol. 2017, 232, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.X.; Zhao, T.W.; Sheng, Y.J.; Zheng, T.; Fu, L.Z.; Zhang, Y.S. Dendrobium officinale Kimura et migo: A review on its ethnopharmacology, phytochemistry, pharmacology, and industrialization. Evid.-Based Complement. Altern. Med. 2017, 2017, 7436259. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Sun, P.; Qian, Y.; Suo, H.Y.D. Candidum has in vitro anticancer effects in HCT-116 cancer cells and exerts in vivo anti-metastatic effects in mice. Nutr. Res. Pract. 2014, 8, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.Z.; Sun, S.R.; Zhang, Y.M.; Sun, S.; Cao, Z.W. Inhibitory effect of Dendrobium officinale polysaccharide on growth of human breast cancer MCF-7 cells and the related mechanism. Biomed. Res. 2017, 28, 1922–1926. [Google Scholar]
- Zhao, Y.; Liu, Y.; Lan, X.M.; Xu, G.L.; Sun, Y.Z.; Li, F.; Liu, H.N. Effect of Dendrobium officinale extraction on gastric carcinogenesis in rats. Evid.-Based Complement. Altern. Med. 2016, 2016, 1213090. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, Y.L.; Lin, S.Q. HPLC analysis of flavonoids in the root of six Glycyrrhiza species. Acta Pharm. Sin. 1990, 25, 840–848. [Google Scholar]
- Ye, Z.; Dai, J.R.; Zhang, C.G.; Lu, Y.; Wu, L.L.; Gong, A.G.W.; Xu, H.; Tsim, K.W.K.; Wang, Z.T. Chemical differentiation of Dendrobium officinale and Dendrobium devonianum by using UPLC fingerprints, HPLC-ESI-MS, and HPTLC analyses. Evid.-Based Complement. Altern. Med. 2017, 2017, 8647212. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, C.F.; Wang, Z.T.; Zhang, M.; Xu, L.S. Five new compounds from Dendrobium crystallinum. J. Asian Nat. Prod. Res. 2009, 11, 903–911. [Google Scholar] [CrossRef] [PubMed]
- Flamini, G. Flavonoids and other compounds from the aerial parts of Viola etrusca. Chem. Biodivers. 2007, 4, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Deng, G.L.; Zeng, S.; Ma, J.L.; Zhang, Y.; Qu, Y.L.; Han, Y.; Yin, L.; Cai, C.J.; Guo, C.; Shen, H. The anti-tumor activities of Neferine on cell invasion and Oxaliplatin sensitivity regulated by EMT via Snail signaling in hepatocellular carcinoma. Sci. Rep. 2017, 7, 41616. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef] [PubMed]
- Lo, U.G.; Lee, C.F.; Lee, M.S.; Hsieh, J.T. The role and mechanism of epithelial-to-mesenchymal transition in prostate cancer progression. Int. J. Mol. Sci. 2017, 18, 2079. [Google Scholar] [CrossRef] [PubMed]
- Budhathoki, N.; Shah, B.K. Survival of elderly patients with advanced hepatocellular carcinoma with distant metastasis in pre- and post-sorafenib era: A population based study. J. Clin. Oncol. 2017, 35, e15643. [Google Scholar]
- Tovoli, F.; De Lorenzo, S.; Dekel, B.G.S.; Piscaglia, F.; Benevento, F.; Brandi, G.; Bolondi, L. Oral Oxycodone/Naloxone for pain control in cirrhosis: Observational study in patients with symptomatic metastatic hepatocellular carcinoma. Liver Int. 2018, 38, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Nakaya, Y.; Sheng, G.J. EMT in developmental morphogenesis. Cancer Lett. 2013, 341, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Kogure, T.; Kondo, Y.; Kakazu, E.; Ninomiya, M.; Kimura, O.; Iwata, T.; Morosawa, T.; Shimosegawa, T. Involvement of Mir-29a in epigenetic regulation of E-cadherin during TGF-β-induced epithelial-mesenchymal transition in hepatocellular carcinoma. Hepatology 2012, 56, 615A. [Google Scholar]
- Liu, X.N.; Wang, S.; Yang, Q.; Wang, Y.J.; Chen, D.X.; Zhu, X.X. ESC reverses epithelial mesenchymal transition induced by transforming growth factor-β via inhibition of Smad signal pathway in HepG2 liver cancer cells. Cancer Cell Int. 2015, 15, 114. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Sun, P.; Li, G.J.; Zhu, K.; Wang, C.; Zhao, X. Inhibitory effects of Dendrobium candidum wall ex lindl. On azoxymethane- and dextran sulfate sodium-induced colon carcinogenesis in c57bl/6 mice. Oncol. Lett. 2014, 7, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Ru, N.Y.; Wu, J.; Chen, Z.N.; Bian, H.J. Hab18g/CD147 is involved in TGF-β-induced epithelial-mesenchymal transition and hepatocellular carcinoma invasion. Cell Biol. Int. 2015, 39, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Malfettone, A.; Soukupova, J.; Fernando, J.; Koudelkova, P.; Bertran, E.; Fabra, A.; Grubinger, M.; Rani, B.; Giannelli, G.; Mikulits, W.; et al. Crosstalk between TGF-β-induced epithelial-mesenchymal transition and stemness in hepatocellular carcinoma. J. Hepatol. 2016, 64, S557. [Google Scholar] [CrossRef]
- Jiao, D.M.; Wang, J.; Lu, W.; Tang, X.L.; Chen, J.; Mou, H.; Chen, Q.Y. Curcumin inhibited TGF-induced EMT and angiogenesis through regulating c-met dependent PI3K/Akt/mTOR signaling pathways in lung cancer. Mol. Ther.-Oncolytics 2016, 3, 16018. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; He, Y.C.; Qi, L.S.; Chen, L.; Luo, Y.; Chen, L.W.; Li, Y.M.; Zhang, N.; Guo, H. cPLA2α activates PI3K/Akt and inhibits Smad2/3 during epithelial-mesenchymal transition of hepatocellular carcinoma cells. Cancer Lett. 2017, 403, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.C.; Liang, Y.J.; Zheng, T.S.; Song, R.P.; Wang, J.B.; Shi, H.W.; Sun, B.S.; Xie, C.M.; Li, Y.J.; Han, J.H.; et al. Fcn2 inhibits epithelial-mesenchymal transition-induced metastasis of hepatocellular carcinoma via TGF-β/Smad signaling. Cancer Lett. 2016, 378, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.H.; Xing, S.P.; Wang, K.; Yi, H.; Du, B.Y. Paeonol attenuates aging MRC-5 cells and inhibits epithelial-mesenchymal transition of premalignant HaCaT cells induced by aging MRC-5 cell-conditioned medium. Mol. Cell. Biochem. 2018, 439, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.W.; Li, Y.R.; Tao, S.C.; Wei, G.; Huang, Y.C.; Chen, D.F.; Wu, C.F. Purification, characterization and biological activity of polysaccharides from Dendrobium officinale. Molecules 2016, 21, 701. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.H.; Huang, Y.C.; Huang, J.B.; Lin, L.Z.; Wei, G. Gigantol attenuates the proliferation of human liver cancer HepG2 cells through the PI3K/Akt/NF-κB signaling pathway. Oncol. Rep. 2017, 37, 865–870. [Google Scholar] [CrossRef] [PubMed]








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xing, S.; Yu, W.; Zhang, X.; Luo, Y.; Lei, Z.; Huang, D.; Lin, J.; Huang, Y.; Huang, S.; Nong, F.; et al. Isoviolanthin Extracted from Dendrobium officinale Reverses TGF-β1-Mediated Epithelial–Mesenchymal Transition in Hepatocellular Carcinoma Cells via Deactivating the TGF-β/Smad and PI3K/Akt/mTOR Signaling Pathways. Int. J. Mol. Sci. 2018, 19, 1556. https://doi.org/10.3390/ijms19061556
Xing S, Yu W, Zhang X, Luo Y, Lei Z, Huang D, Lin J, Huang Y, Huang S, Nong F, et al. Isoviolanthin Extracted from Dendrobium officinale Reverses TGF-β1-Mediated Epithelial–Mesenchymal Transition in Hepatocellular Carcinoma Cells via Deactivating the TGF-β/Smad and PI3K/Akt/mTOR Signaling Pathways. International Journal of Molecular Sciences. 2018; 19(6):1556. https://doi.org/10.3390/ijms19061556
Chicago/Turabian StyleXing, Shangping, Wenxia Yu, Xiaofeng Zhang, Yingyi Luo, Zhouxi Lei, Dandan Huang, Ji Lin, Yuechun Huang, Shaowei Huang, Feifei Nong, and et al. 2018. "Isoviolanthin Extracted from Dendrobium officinale Reverses TGF-β1-Mediated Epithelial–Mesenchymal Transition in Hepatocellular Carcinoma Cells via Deactivating the TGF-β/Smad and PI3K/Akt/mTOR Signaling Pathways" International Journal of Molecular Sciences 19, no. 6: 1556. https://doi.org/10.3390/ijms19061556