High-Throughput Sequencing of microRNAs in Glucocorticoid Sensitive Paediatric Inflammatory Bowel Disease Patients

 ,
,  ,
,  , , and
, , and
Abstract
:
1. Introduction
2. Results
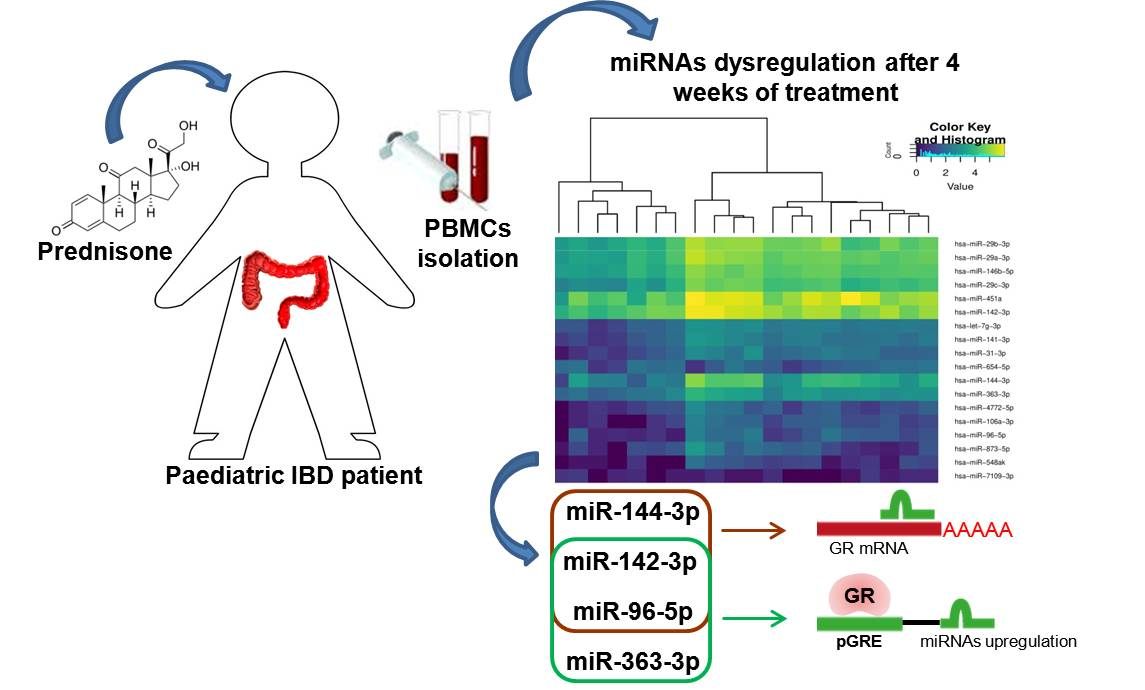
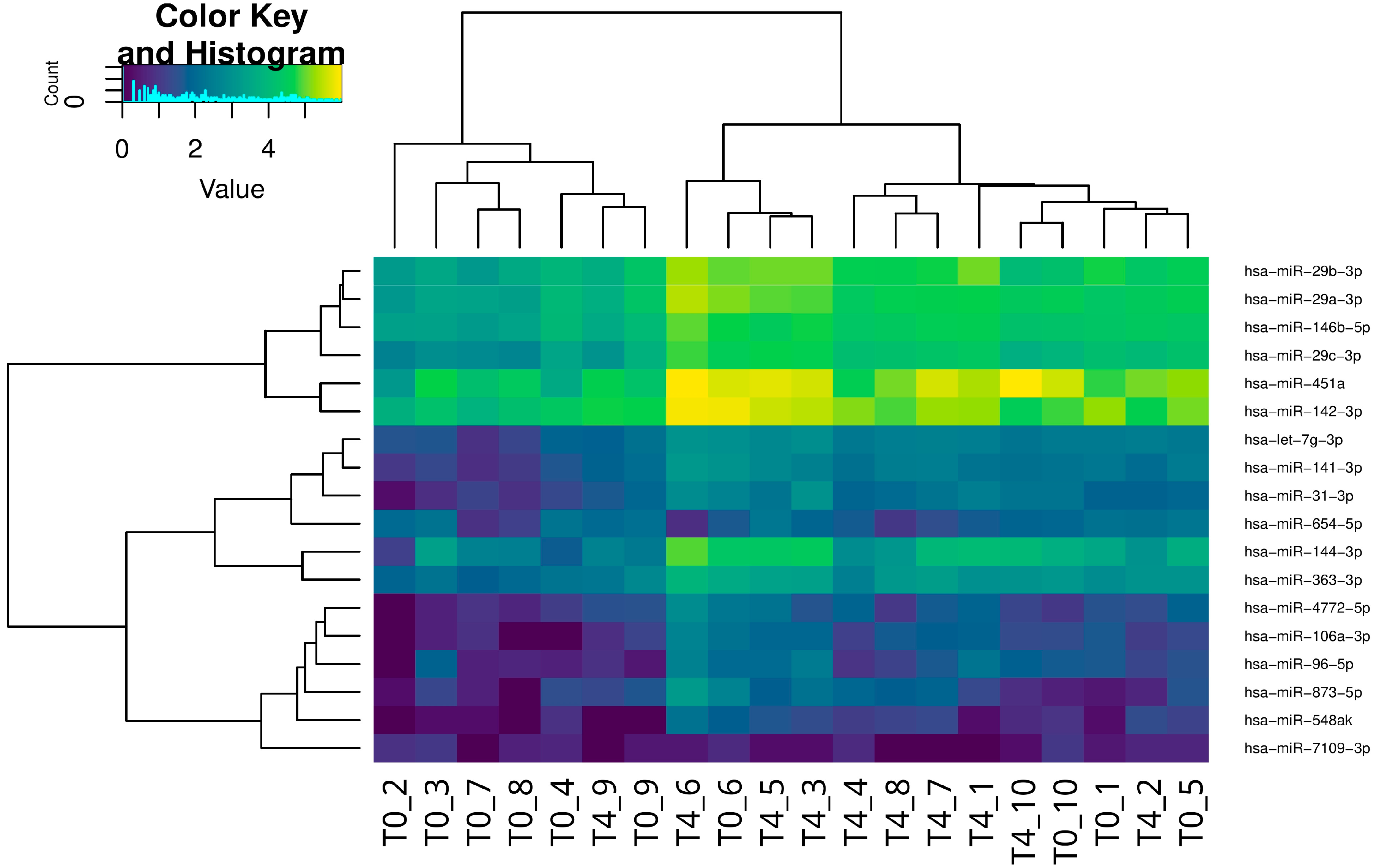
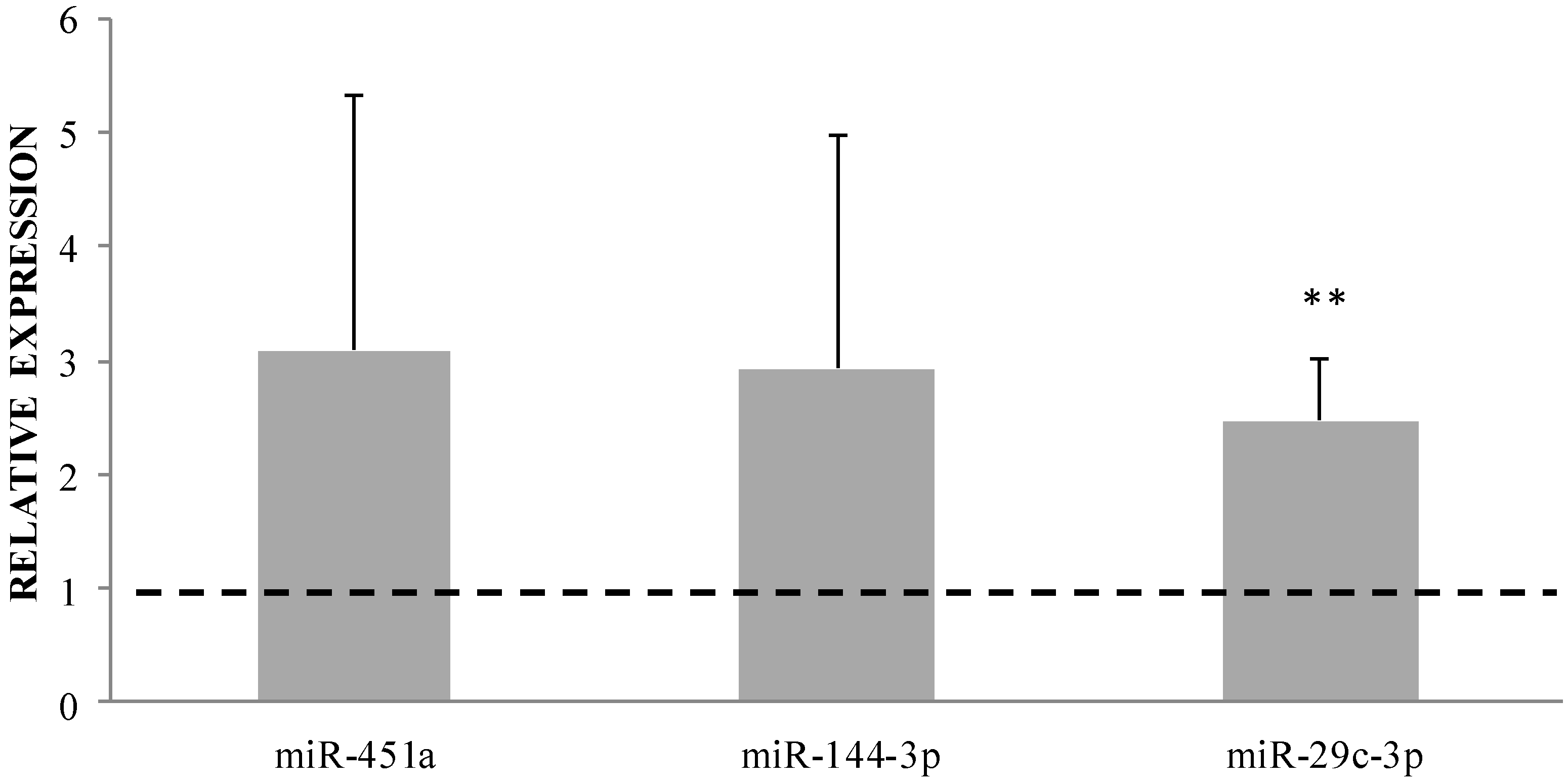
2.1. miRNA Analysis
2.2. mRNA Analysis
2.3. Integration between miRNA and mRNA Analysis
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. RNA Extraction
4.3. miRNA-seq
4.4. miRNAs Identification
4.5. Differential miRNA Expression Analysis
4.6. Validation of Selected Differentially Expressed miRNAs by qRT-PCR
4.7. TaqMan Gene Expression Analysis
4.8. Identification of Putative mRNA Targets
4.9. Identification of GC Responsive Elements (GRE)
4.10. Statistical Analysis
4.11. Ethical Considerations
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| GC | Glucocorticoid |
| IBD | Inflammatory bowel disease |
| miRNA | Micro RNA |
| GR | Glucocorticoid receptor |
| GRE | Glucocorticoid responsive elements |
References
- De Iudicibus, S.; Franca, R.; Martelossi, S.; Ventura, A.; Decorti, G. Molecular mechanism of glucocorticoid resistance in inflammatory bowel disease. World J. Gastroenterol. 2011, 17, 1095–1108. [Google Scholar] [CrossRef] [PubMed]
- Grad, I.; Picard, D. The glucocorticoid responses are shaped by molecular chaperones. Mol. Cell. Endocrinol. 2007, 275, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Gross, K.L.; Lu, N.Z.; Cidlowski, J.A. Molecular mechanisms regulating glucocorticoid sensitivity and resistance. Mol. Cell. Endocrinol. 2009, 300, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Raddatz, D.; Middel, P.; Bockemuhl, M.; Benohr, P.; Wissmann, C.; Schworer, H.; Ramadori, G. Glucocorticoid receptor expression in inflammatory bowel disease: Evidence for a mucosal down-regulation in steroid-unresponsive ulcerative colitis. Aliment. Pharmacol. Ther. 2004, 19, 47–61. [Google Scholar] [CrossRef] [PubMed]
- Matysiak, M.; Makosa, B.; Walczak, A.; Selmaj, K. Patients with multiple sclerosis resisted to glucocorticoid therapy: Abnormal expression of heat-shock protein 90 in glucocorticoid receptor complex. Mult. Scler. 2008, 14, 919–926. [Google Scholar] [CrossRef] [PubMed]
- Qian, X.; Zhu, Y.; Xu, W.; Lin, Y. Glucocorticoid receptor and heat shock protein 90 in peripheral blood mononuclear cells from asthmatics. Chin. Med. J. 2001, 114, 1051–1054. [Google Scholar] [PubMed]
- Guo, H.; Ingolia, N.T.; Weissman, J.S.; Bartel, D.P. Mammalian microRNAs predominantly act to decrease target mRNA levels. Nature 2010, 466, 835–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalal, S.R.; Kwon, J.H. The role of MicroRNA in inflammatory bowel disease. Gastroenterol. Hepatol. 2010, 6, 714–722. [Google Scholar]
- Kalla, R.; Ventham, N.T.; Kennedy, N.A.; Quintana, J.F.; Nimmo, E.R.; Buck, A.H.; Satsangi, J. MicroRNAs: New players in IBD. Gut 2015, 64, 504–517. [Google Scholar] [CrossRef] [PubMed]
- Iborra, M.; Bernuzzi, F.; Invernizzi, P.; Danese, S. MicroRNAs in autoimmunity and inflammatory bowel disease: Crucial regulators in immune response. Autoimmun. Rev. 2012, 11, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Archanioti, P.; Gazouli, M.; Theodoropoulos, G.; Vaiopoulou, A.; Nikiteas, N. Micro-RNAs as regulators and possible diagnostic bio-markers in inflammatory bowel disease. J. Crohns Colitis 2011, 5, 520–524. [Google Scholar] [CrossRef] [PubMed]
- Coskun, M.; Bjerrum, J.T.; Seidelin, J.B.; Nielsen, O.H. MicroRNAs in inflammatory bowel disease—Pathogenesis, diagnostics and therapeutics. World J. Gastroenterol. 2012, 18, 4629–4634. [Google Scholar] [CrossRef] [PubMed]
- Sionov, R.V. MicroRNAs and Glucocorticoid-Induced Apoptosis in Lymphoid Malignancies. ISRN hematol. 2013, 2013, 348212. [Google Scholar] [CrossRef] [PubMed]
- Riester, A.; Issler, O.; Spyroglou, A.; Rodrig, S.H.; Chen, A.; Beuschlein, F. ACTH-dependent regulation of microRNA as endogenous modulators of glucocorticoid receptor expression in the adrenal gland. Endocrinology 2014, 153, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Iwaya, T.; Yokobori, T.; Nishida, N.; Kogo, R.; Sudo, T.; Tanaka, F.; Shibata, K.; Sawada, G.; Takahashi, Y.; Ishibashi, M.; et al. Downregulation of miR-144 is associated with colorectal cancer progression via activation of mTOR signaling pathway. Carcinogenesis 2012, 33, 2391–2397. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Wu, J.; Zhao, J.; Wang, H.; Liu, Y.; Chen, T.; Kan, X.; Tao, Q.; Shen, X.; Yan, K.; et al. miR-29b and miR-29c are involved in Toll-like receptor control of glucocorticoid-induced apoptosis in human plasmacytoid dendritic cells. PLoS ONE 2013, 8, e69926. [Google Scholar] [CrossRef] [PubMed]
- Chuang, T.D.; Pearce, W.J.; Khorram, O. miR-29c induction contributes to downregulation of vascular extracellular matrix proteins by glucocorticoids. Am. J. Physiol. Cell Physiol. 2015, 309, C117–C125. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.K.; Shah, R.R.; Cidlowski, J.A. Glucocorticoids modulate microRNA expression and processing during lymphocyte apoptosis. J. Biol. Chem. 2010, 285, 36698–36708. [Google Scholar] [CrossRef] [PubMed]
- Palagani, A.; Op de Beeck, K.; Naulaerts, S.; Diddens, J.; Sekhar Chirumamilla, C.; Van Camp, G.; Laukens, K.; Heyninck, K.; Gerlo, S.; Mestdagh, P.; et al. Ectopic microRNA-150-5p transcription sensitizes glucocorticoid therapy response in MM1S multiple myeloma cells but fails to overcome hormone therapy resistance in MM1R cells. PLoS ONE 2014, 9, e113842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hafner, M.; Landthaler, M.; Burger, L.; Khorshid, M.; Hausser, J.; Berninger, P.; Rothballer, A.; Ascano, M. Jr.; Jungkamp, A.C.; Munschauer, M.; et al. Transcriptome-wide identification of RNA-binding protein and microRNA target sites by PAR-CLIP. Cell 2010, 141, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Fujioka, S.; Nakamichi, I.; Esaki, M.; Asano, K.; Matsumoto, T.; Kitazono, T. Serum microRNA levels in patients with Crohn’s disease during induction therapy by infliximab. J. Gastroenterol. Hepatol. 2014, 29, 1207–1214. [Google Scholar] [CrossRef] [PubMed]
- Heier, C.R.; Fiorillo, A.A.; Chaisson, E.; Gordish-Dressman, H.; Hathout, Y.; Damsker, J.M.; Hoffman, E.P.; Conklin, L.S. Identification of Pathway-Specific Serum Biomarkers of Response to Glucocorticoid and Infliximab Treatment in Children with Inflammatory Bowel Disease. Clin. Transl. Gastroenterol. 2016, 7, e192. [Google Scholar] [CrossRef] [PubMed]
- McBeth, L.; Nwaneri, A.C.; Grabnar, M.; Demeter, J.; Nestor-Kalinoski, A.; Hinds, T.D., Jr. Glucocorticoid receptor beta increases migration of human bladder cancer cells. Oncotarget 2016, 7, 27313–27324. [Google Scholar] [CrossRef] [PubMed]
- Meijsing, S.H.; Pufall, M.A.; So, A.Y.; Bates, D.L.; Chen, L.; Yamamoto, K.R. DNA binding site sequence directs glucocorticoid receptor structure and activity. Science 2009, 324, 407–410. [Google Scholar] [CrossRef] [PubMed]
- Vermeer, H.; Hendriks-Stegeman, B.I.; van der Burg, B.; van Buul-Offers, S.C.; Jansen, M. Glucocorticoid-induced increase in lymphocytic FKBP51 messenger ribonucleic acid expression: A potential marker for glucocorticoid sensitivity, potency, and bioavailability. J. Clin. Endocrinol. Metab. 2003, 88, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Freedman, L.P.; Luisi, B.F. On the mechanism of DNA binding by nuclear hormone receptors: A structural and functional perspective. J. Cell. Biochem. 1993, 51, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Yu, J.; Bi, J.; Qi, H.; Di, W.; Wu, L.; Wang, L.; Zha, J.; Lv, S.; Zhang, F.; et al. Glucocorticoids transcriptionally regulate miR-27b expression promoting body fat accumulation via suppressing the browning of white adipose tissue. Diabetes 2014, 64, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet.journal 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Risso, D.; Schwartz, K.; Sherlock, G.; Dudoit, S. GC-content normalization for RNA-Seq data. BMC Bioinform. 2011, 12, 480. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.D.; Tseng, Y.T.; Shrestha, S.; Lin, Y.L.; Khaleel, A.; Chou, C.H.; Chu, C.F.; Huang, H.Y.; Lin, C.M.; Ho, S.Y.; et al. miRTarBase update 2014: An information resource for experimentally validated miRNA-target interactions. Nucleic Acids Res. 2014, 42, D78–D85. [Google Scholar] [CrossRef] [PubMed]
- Grant, C.E.; Bailey, T.L.; Noble, W.S. FIMO: Scanning for occurrences of a given motif. Bioinformatics 2011, 27, 1017–1018. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Williams, N.; Misleh, C.; Li, W.W. MEME: Discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res. 2006, 34, W369–W373. [Google Scholar] [CrossRef] [PubMed]
- Matys, V.; Fricke, E.; Geffers, R.; Gossling, E.; Haubrock, M.; Hehl, R.; Hornischer, K.; Karas, D.; Kel, A.E.; Kel-Margoulis, O.V.; et al. TRANSFAC: Transcriptional regulation, from patterns to profiles. Nucleic Acids Res. 2003, 31, 374–378. [Google Scholar] [CrossRef] [PubMed]
- Surjit, M.; Ganti, K.P.; Mukherji, A.; Ye, T.; Hua, G.; Metzger, D.; Li, M.; Chambon, P. Widespread negative response elements mediate direct repression by agonist-liganded glucocorticoid receptor. Cell 2011, 145, 224–241. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
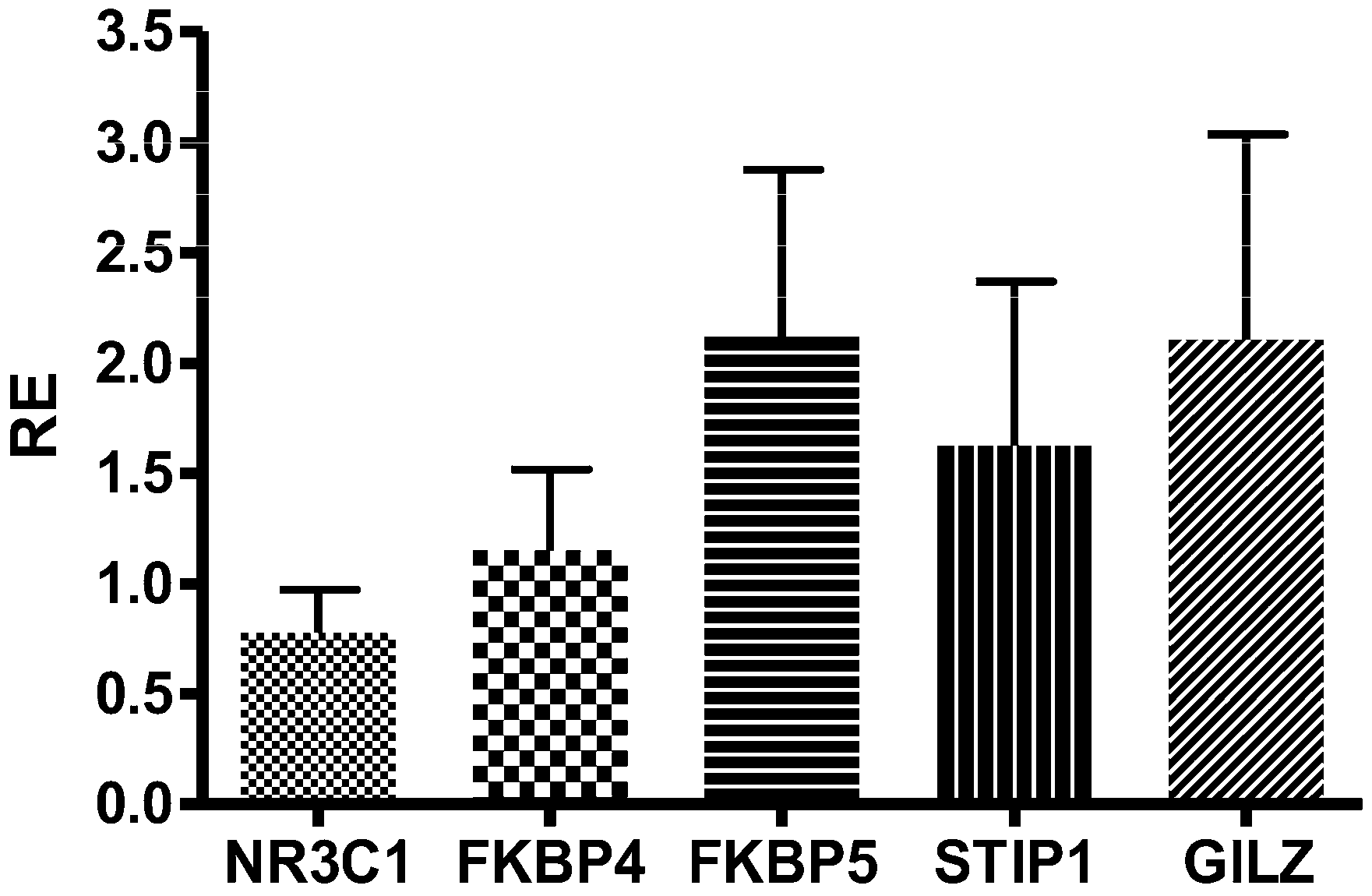{kind=link}
| Upregulated miRNAs | FC | FDR Corrected P-Value | Downregulated miRNAs | FC | FDR Corrected p-Value |
|---|---|---|---|---|---|
| hsa-miR-451a * [13] hsa-miR-144-3p * [14,15] hsa-miR-96-5p * [13,14] hsa-miR-29b-3p * [13] hsa-miR-142-3p * [14] hsa-miR-873-5p hsa-miR-29c-3p * [16,17] hsa-miR-29a-3p * [13] hsa-miR-363-3p hsa-miR-141-3p hsa-miR-548ak hsa-let-7g-3p* [18] hsa-miR-4772-5p hsa-miR-106a-3p hsa-miR-31-3p hsa-miR-146b-5p * [19] | 4.16 4.44 2.96 2.89 2.21 3.36 3.37 2.72 2.31 2.59 3.11 2.44 2.70 3.52 3.36 2.27 | 1.66 × 10−6 1.04 × 10−5 6.38 × 10−3 0.026 0.026 0.026 0.037 0.041 0.041 0.041 0.042 0.042 0.047 0.047 0.049 0.049 | hsa-miR-7109-3p hsa-miR-654-5p | −4.62 −2.27 | 0.044 0.049 |
| miRNA | pGRE | Start | End | Strand | Chrom | Expression |
|---|---|---|---|---|---|---|
| hsa-miR-363 | GTGATAATGTGTGCTT | 133303695 | 133303710 | − | chrX | Up |
| hsa-miR-96 | AGGACAAAGAGTCCTC | 129416083 | 129416098 | − | chr7 | Up |
| hsa-miR-142 | CTCACCTTCAGTTCTG | 58331606 | 58331621 | + | Chr17 | Up |
| hsa-miR-142 | CTGTCAGTCTGTCCTC | 58332656 | 58332671 | − | Chr17 | Up |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Iudicibus, S.; Lucafò, M.; Vitulo, N.; Martelossi, S.; Zimbello, R.; De Pascale, F.; Forcato, C.; Naviglio, S.; Di Silvestre, A.; Gerdol, M.; et al. High-Throughput Sequencing of microRNAs in Glucocorticoid Sensitive Paediatric Inflammatory Bowel Disease Patients. Int. J. Mol. Sci. 2018, 19, 1399. https://doi.org/10.3390/ijms19051399
De Iudicibus S, Lucafò M, Vitulo N, Martelossi S, Zimbello R, De Pascale F, Forcato C, Naviglio S, Di Silvestre A, Gerdol M, et al. High-Throughput Sequencing of microRNAs in Glucocorticoid Sensitive Paediatric Inflammatory Bowel Disease Patients. International Journal of Molecular Sciences. 2018; 19(5):1399. https://doi.org/10.3390/ijms19051399
Chicago/Turabian StyleDe Iudicibus, Sara, Marianna Lucafò, Nicola Vitulo, Stefano Martelossi, Rosanna Zimbello, Fabio De Pascale, Claudio Forcato, Samuele Naviglio, Alessia Di Silvestre, Marco Gerdol, and et al. 2018. "High-Throughput Sequencing of microRNAs in Glucocorticoid Sensitive Paediatric Inflammatory Bowel Disease Patients" International Journal of Molecular Sciences 19, no. 5: 1399. https://doi.org/10.3390/ijms19051399
APA StyleDe Iudicibus, S., Lucafò, M., Vitulo, N., Martelossi, S., Zimbello, R., De Pascale, F., Forcato, C., Naviglio, S., Di Silvestre, A., Gerdol, M., Stocco, G., Valle, G., Ventura, A., Bramuzzo, M., & Decorti, G. (2018). High-Throughput Sequencing of microRNAs in Glucocorticoid Sensitive Paediatric Inflammatory Bowel Disease Patients. International Journal of Molecular Sciences, 19(5), 1399. https://doi.org/10.3390/ijms19051399