Evaluation of an Internally Controlled Multiplex Tth Endonuclease Cleavage Loop-Mediated Isothermal Amplification (TEC-LAMP) Assay for the Detection of Bacterial Meningitis Pathogens

 ,
,
Abstract
:1. Introduction
2. Results
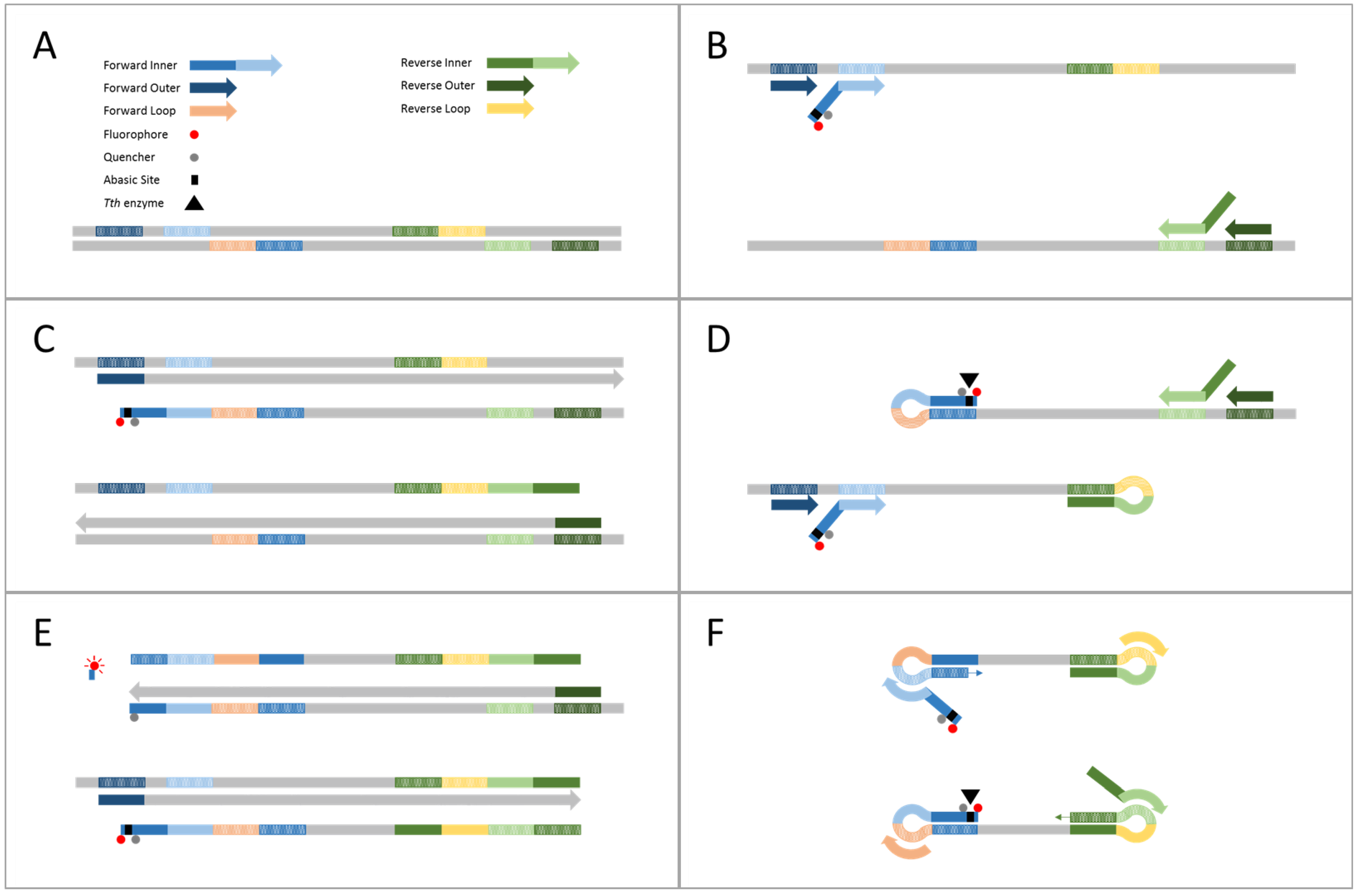
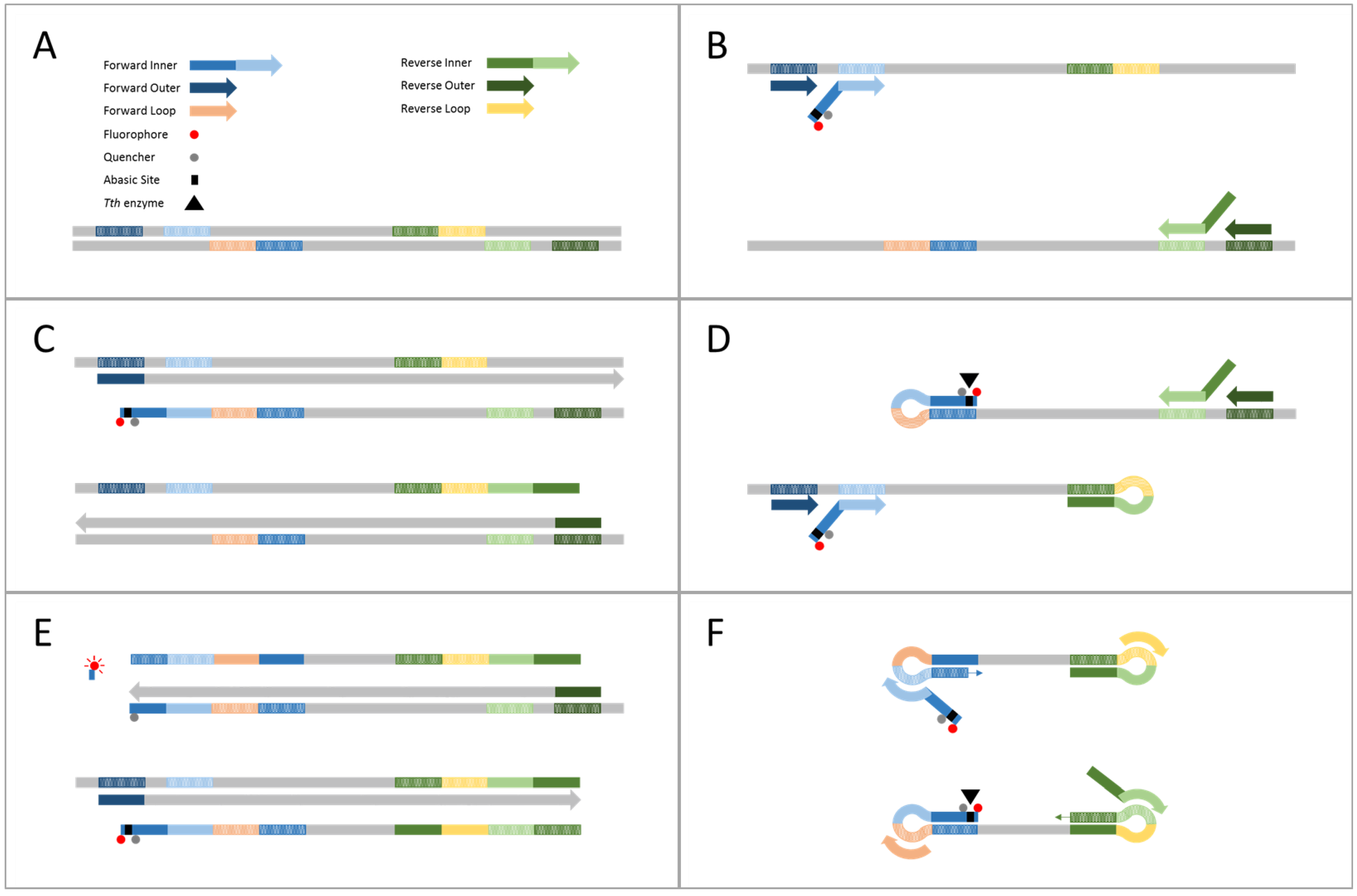
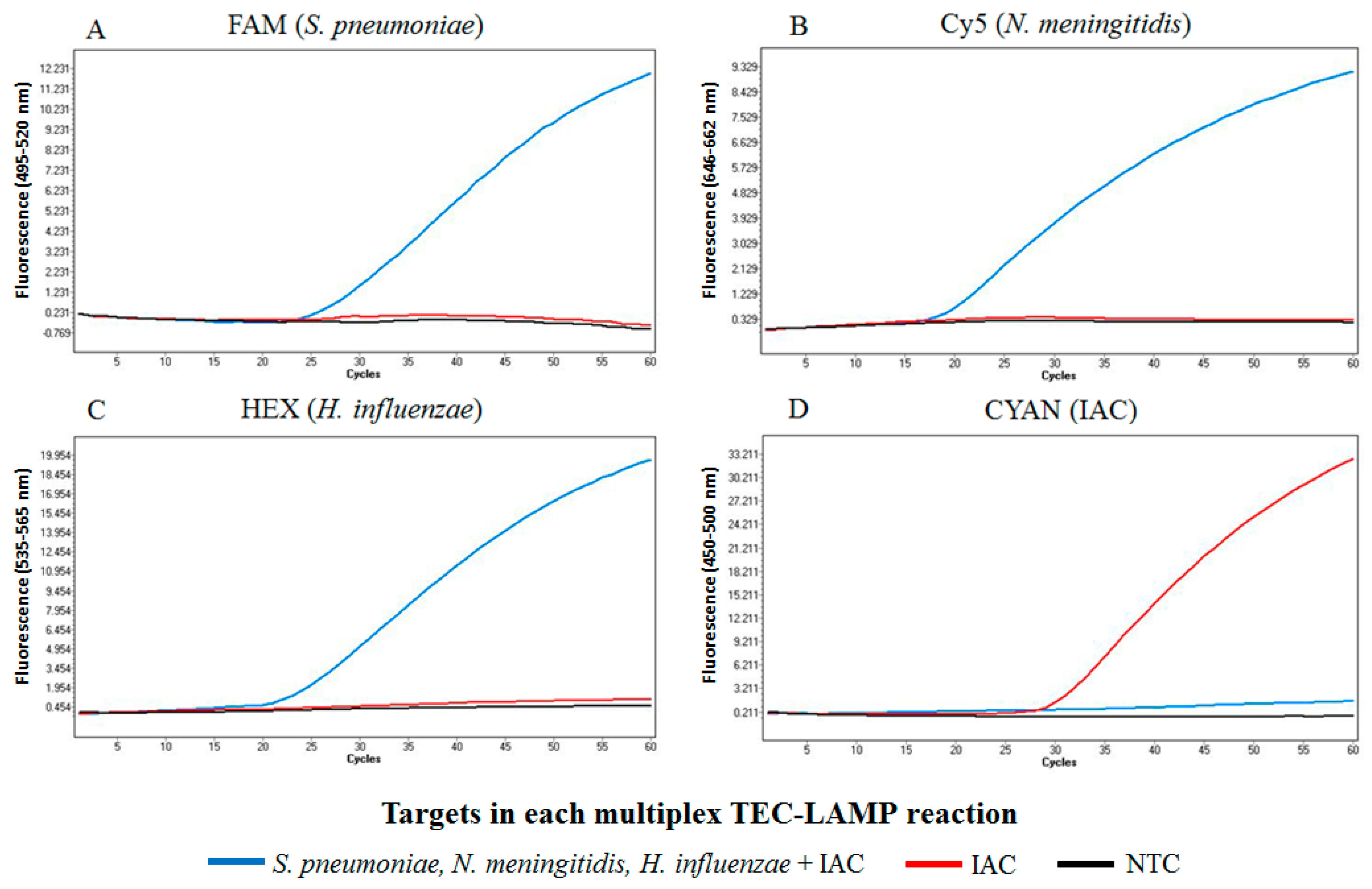
2.1. Demonstration of Internally Controlled Multiplex TEC-LAMP Detection
2.2. Analytical Specificity and Limit of Detection
2.3. Clinical Evaluation
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, DNA Isolation and Quantification
4.2. Diagnostic Targets and TEC-LAMP Oligonucleotides
4.3. Internally Controlled Multiplex TEC-LAMP Assay
4.4. Demonstration of Internally Controlled Multiplex TEC-LAMP Detection
4.5. Analytical Specificity and Limit of Detection
4.6. Clinical Evaluation
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Brouwer, M.C.; Tunkel, A.R.; van de Beek, D. Epidemiology, diagnosis, and antimicrobial treatment of acute bacterial meningitis. Clin. Microbiol. Rev. 2010, 23, 467–492. [Google Scholar] [CrossRef] [PubMed]
- McGill, F.; Heyderman, R.S.; Panagiotou, S.; Tunkel, A.R.; Solomon, T. Acute bacterial meningitis in adults. Lancet 2016, 388, 3036–3047. [Google Scholar] [CrossRef]
- Kim, K.S. Acute bacterial meningitis in infants and children. Lancet Infect. Dis. 2010, 10, 32–42. [Google Scholar] [CrossRef]
- Van de Beek, D. Progress and challenges in bacterial meningitis. Lancet 2012, 380, 1623–1624. [Google Scholar] [CrossRef]
- Paireau, J.; Chen, A.; Broutin, H.; Grenfell, B.; Basta, N.E. Seasonal dynamics of bacterial meningitis: A time-series analysis. Lancet Glob. Health 2016, 4, e370–e377. [Google Scholar] [CrossRef]
- Farahani, H.; Ghaznavi-Rad, E.; Mondanizadeh, M.; MirabSamiee, S.; Khansarinejad, B. Specific detection of common pathogens of acute bacterial meningitis using an internally controlled tetraplex-PCR assay. Mol. Cell. Probes 2016, 30, 261–265. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, P.B.; O’Brien, K.L.; Greenwood, B.; van de Beek, D. Effect of vaccines on bacterial meningitis worldwide. Lancet 2012, 380, 1703–1711. [Google Scholar] [CrossRef]
- Watt, J.P.; Wolfson, L.J.; O’Brien, K.L.; Henkle, E.; Deloria-Knoll, M.; McCall, N.; Lee, E.; Levine, O.S.; Hajjeh, R.; Mulholland, K. Burden of disease caused by Haemophilus influenzae type b in children younger than 5 years: Global estimates. Lancet 2009, 374, 903–911. [Google Scholar] [CrossRef]
- Proulx, N.; Frechette, D.; Toye, B.; Chan, J.; Kravcik, S. Delays in the administration of antibiotics are associated with mortality from adult acute bacterial meningitis. QJM 2005, 98, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Leekha, S.; Terrell, C.L.; Edson, R.S. General Principles of Antimicrobial Therapy. Elsevier 2011, 86, 156–167. [Google Scholar] [CrossRef] [PubMed]
- Corless, C.E.; Guiver, M.; Borrow, R.; Edwards-Jones, V.; Fox, A.J.; Kaczmarski, E.B. Simultaneous detection of Neisseria meningitidis, Haemophilus influenzae, and Streptococcus pneumoniae in suspected cases of meningitis and septicemia using real-time PCR. J. Clin. Microbiol. 2001, 39, 1553–1558. [Google Scholar] [CrossRef] [PubMed]
- Brouwer, M.C.; Thwaites, G.E.; Tunkel, A.R.; van de Beek, D. Dilemmas in the diagnosis of acute community-acquired bacterial meningitis. Lancet 2012, 380, 1684–1692. [Google Scholar] [CrossRef]
- Espy, M.; Uhl, J.; Sloan, L.; Buckwalter, S.; Jones, M.; Vetter, E.; Yao, J.; Wengenack, N.; Rosenblatt, J.; Cockerill, F. Real-time PCR in clinical microbiology: Applications for routine laboratory testing. Clin. Microbiol. Rev. 2006, 19, 165–256. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.M.; Cordeiro, S.M.; Harcourt, B.H.; Carvalho, M.; Azevedo, J.; Oliveira, T.Q.; Leite, M.C.; Salgado, K.; Reis, M.G.; Plikaytis, B.D. Accuracy of real-time PCR, gram stain and culture for Streptococcus pneumoniae, Neisseria meningitidis and Haemophilus influenzae meningitis diagnosis. BMC Infect. Dis. 2013, 13, 26. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Theodore, M.J.; Mair, R.; Trujillo-Lopez, E.; du Plessis, M.; Wolter, N.; Baughman, A.L.; Hatcher, C.; Vuong, J.; Lott, L.; et al. Clinical validation of multiplex real-time PCR assays for detection of bacterial meningitis pathogens. J. Clin. Microbiol. 2012, 50, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucl. Acids Res. 2000, 28, e63. [Google Scholar] [CrossRef] [PubMed]
- Nagamine, K.; Hase, T.; Notomi, T. Accelerated reaction by loop-mediated isothermal amplification using loop primers. Mol. Cell. Probes 2002, 16, 223–229. [Google Scholar] [CrossRef] [PubMed]
- McKenna, J.P.; Fairley, D.J.; Shields, M.D.; Cosby, S.L.; Wyatt, D.E.; McCaughey, C.; Coyle, P.V. Development and clinical validation of a loop-mediated isothermal amplification method for the rapid detection of Neisseria meningitidis. Diagn. Microbiol. Infect. Dis. 2011, 69, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Kim, E.J.; Kilgore, P.E.; Kim, S.A.; Takahashi, H.; Ohnishi, M.; Anh, D.D.; Dong, B.Q.; Kim, J.S.; Tomono, J.; et al. Clinical evaluation of a loop-mediated isothermal amplification (LAMP) assay for rapid detection of Neisseria meningitidis in cerebrospinal fluid. PLoS ONE 2015, 10, e0122922. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, H.; Kawana, T.; Fukushima, E.; Suzutani, T. Tolerance of loop-mediated isothermal amplification to a culture medium and biological substances. J. Biochem. Biophys. Methods 2007, 70, 499–501. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, D.M.; Kubota, R.; Dong, J.; Li, Y.; Higashiguchi, D. Handheld device for real-time, quantitative, lamp-based detection of Salmonella enterica using assimilating probes. Biosens. Bioelectron. 2011, 30, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Kubota, R.; Labarre, P.; Weigl, B.H.; Li, Y.; Haydock, P.; Jenkins, D.M. Molecular diagnostics in a teacup: Non-instrumented nucleic acid amplification (NINA) for rapid, low cost detection of Salmonella enterica. Chin. Sci. Bull. 2013, 58, 1162–1168. [Google Scholar] [CrossRef] [PubMed]
- Dou, M.; Sanjay, S.T.; Dominguez, D.C.; Liu, P.; Xu, F.; Li, X. Multiplexed instrument-free meningitis diagnosis on a polymer/paper hybrid microfluidic biochip. Biosens. Bioelectron. 2017, 87, 865–873. [Google Scholar] [CrossRef] [PubMed]
- Ball, C.S.; Light, Y.K.; Koh, C.Y.; Wheeler, S.S.; Coffey, L.L.; Meagher, R.J. Quenching of unincorporated amplification signal reporters in reverse-transcription loop-mediated isothermal amplification enabling bright, single-step, closed-tube, and multiplexed detection of RNA viruses. Anal. Chem. 2016, 88, 3562–3568. [Google Scholar] [CrossRef] [PubMed]
- Priye, A.; Bird, S.W.; Light, Y.K.; Ball, C.S.; Negrete, O.A.; Meagher, R.J. A smartphone-based diagnostic platform for rapid detection of zika, chikungunya, and dengue viruses. Sci. Rep. 2017, 7, 44778. [Google Scholar] [CrossRef] [PubMed]
- Aliotta, J.M.; Pelletier, J.J.; Ware, J.L.; Moran, L.S.; Benner, J.S.; Kong, H. Thermostable bst DNA polymerase i lacks a 3′→5′ proofreading exonuclease activity. Genet. Anal. Biomol. Eng. 1996, 12, 185–195. [Google Scholar] [CrossRef]
- Hoorfar, J.; Malorny, B.; Abdulmawjood, A.; Cook, N.; Wagner, M.; Fach, P. Practical considerations in design of internal amplification controls for diagnostic PCR assays. J. Clin. Microbiol. 2004, 42, 1863–1868. [Google Scholar] [CrossRef] [PubMed]
- De Filippis, I.; de Andrade, C.F.; Caldeira, N.; de Azevedo, A.C.; de Almeida, A.E. Comparison of PCR-based methods for the simultaneous detection of Neisseria meningitidis, Haemophilus influenzae, and Streptococcus pneumoniae in clinical samples. Braz. J. Infect. Dis. 2016, 20, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Abdeldaim, G.M.; Strålin, K.; Korsgaard, J.; Blomberg, J.; Welinder-Olsson, C.; Herrmann, B. Multiplex quantitative PCR for detection of lower respiratory tract infection and meningitis caused by Streptococcus pneumoniae, Haemophilus influenzae and Neisseria meningitidis. BMC Microbiol. 2010, 10, 310. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Kim, E.J.; Kilgore, P.E.; Takahashi, H.; Ohnishi, M.; Tomono, J.; Miyamoto, S.; Omagari, D.; Kim, D.W.; Seki, M. A novel loop-mediated isothermal amplification assay for serogroup identification of Neisseria meningitidis in cerebrospinal fluid. Front. Microbiol. 2015, 6, 1548. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.W.; Kilgore, P.E.; Kim, E.J.; Kim, S.A.; Anh, D.D.; Dong, B.Q.; Kim, J.S.; Seki, M. The enhanced pneumococcal lamp assay: A clinical tool for the diagnosis of meningitis due to Streptococcus pneumoniae. PLoS ONE 2012, 7, e42954. [Google Scholar] [CrossRef] [PubMed]
- Coughlan, H.; Reddington, K.; Tuite, N.; Boo, T.W.; Cormican, M.; Barrett, L.; Smith, T.J.; Clancy, E.; Barry, T. Comparative genome analysis identifies novel nucleic acid diagnostic targets for use in the specific detection of Haemophilus influenzae. Diagn. Microbiol. Infect. Dis. 2015, 83, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.W.; Kilgore, P.E.; Kim, E.J.; Kim, S.A.; Anh, D.D.; Seki, M. Loop-mediated isothermal amplification assay for detection of Haemophilus influenzae type b in cerebrospinal fluid. J. Clin. Microbiol. 2011, 49, 3621–3626. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Zhu, S.; Jin, C.; Chen, F. Development of multiplex loop-mediated isothermal amplification-RFLP (mLAMP-RFLP) to detect Salmonella spp. and Shigella spp. in milk. Int. J. Food Microbiol. 2011, 148, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Nurul Najian, A.B.; Engku Nur Syafirah, E.A.; Ismail, N.; Mohamed, M.; Yean, C.Y. Development of multiplex loop mediated isothermal amplification (m-LAMP) label-based gold nanoparticles lateral flow dipstick biosensor for detection of pathogenic leptospira. Anal. Chim. Acta 2016, 903, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Chu, Y.; Cheng, S.; Wu, H.; Kajiyama, T.; Kambara, H.; Zhou, G. Multiplex loop-mediated isothermal amplification detection by sequence-based barcodes coupled with nicking endonuclease-mediated pyrosequencing. Anal. Chem. 2012, 84, 3758–3763. [Google Scholar] [CrossRef] [PubMed]
- Nixon, G.J.; Svenstrup, H.F.; Donald, C.E.; Carder, C.; Stephenson, J.M.; Morris-Jones, S.; Huggett, J.F.; Foy, C.A. A novel approach for evaluating the performance of real time quantitative loop-mediated isothermal amplification-based methods. Biomol. Detect. Quantif. 2014, 2, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.H.; Oh, S.J.; Kim, Y.T.; Kim, S.Y.; Kim, W.J.; Jung, J.; Seo, T.S. Combination of multiplex reverse-transcription loop-mediated isothermal amplification with an immunochromatographic strip for subtyping influenza a virus. Anal. Chim. Acta 2015, 853, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Kasahara, K.; Ishikawa, H.; Sato, S.; Shimakawa, Y.; Watanabe, K. Development of multiplex loop-mediated isothermal amplification assays to detect medically important yeasts in dairy products. FEMS Microbiol. Lett. 2014, 357, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Tanner, N.A.; Zhang, Y.; Evans, T.C., Jr. Simultaneous multiple target detection in real-time loop-mediated isothermal amplification. Biotechniques 2012, 53, 81–89. [Google Scholar] [PubMed]
- Kubota, R.; Jenkins, D.M. Real-time duplex applications of loop-mediated amplification (LAMP) by assimilating probes. Int. J. Mol. Sci. 2015, 16, 4786–4799. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Lan, R.; Xu, H.; Ma, A.; Li, D.; Dai, H.; Yuan, X.; Xu, J.; Ye, C. Multiple endonuclease restriction real-time loop-mediated isothermal amplification: A novel analytically rapid, sensitive, multiplex loop-mediated isothermal amplification detection technique. J. Mol. Diagn. 2015, 17, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Dou, M.; Sanjay, S.T.; Dominguez, D.C.; Zhan, S.; Li, X. A paper/polymer hybrid cd-like microfluidic spinchip integrated with DNA-functionalized graphene oxide nanosensors for multiplex qlamp detection. Chem. Commun. 2017, 53, 10886–10889. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Primer Type | Sequence (5′-3′) |
|---|---|
| S. pneumoniae | |
| TEC primer/probe | (FAM)TGGA(dSpacer)AA(BHQ1-dT)GCTCTGGCTTTTGAAGTGA-CCTACACCAATATCCTCGCT |
| Forward Inner | TGGAAAATGCTCTGGCTTTTGAAGTGA-CCTACACCAATATCCTCGCT |
| Reverse Inner | TCTGTCTGGTAGACAGAATGACGGA-TCTTTGAGAATCAGATGCTGGA |
| Forward Outer | TCCGTCAACGAGGCACAA |
| Reverse Outer | AGCAAACTCACCAAGCGC |
| Forward Loop | TGATGAAACAGACAAGCTGATTCT |
| Reverse Loop | GCGCAATGATGGTATAATCCAG |
| N. meningitidis | |
| TEC primer/probe | (Cy5)TGTC(dSpacer)G(BHQ2-dT)GGCTTTGTTGGTGGTGTCGC-GTGCAAACAGATACGTCCG |
| Forward Inner | TGTCGGTGGCTTTGTTGGTGGTGTCGC-GTGCAAACAGATACGTCCG |
| Reverse Inner | CCGATGTACCAGCACCTTGTCC-GTTTGCGCTGATTACGCCTC |
| Forward Outer | CCCAATTCCACATCAATACGTG |
| Reverse Outer | GTGGTGTCGGTGGTGTTG |
| Forward Loop | GAGATTGTGTTGGGCGGTTTG |
| Reverse Loop | CACCACTTGGAAAAACAGAGGC |
| H. influenzae | |
| TEC primer/probe | (HEX)TGCC(dSpacer)C(BHQ1-dT)GCTTCACGTAAATTATTTGG-TGCTTATTCCTATCGTGGTACG |
| Forward Inner | TGCCGCTGCTTCACGTAAATTATTTGG-TGCTTATTCCTATCGTGGTACG |
| Reverse Inner | CTTGGTTGCTCTCAATGGCAAG-GCACGCCAGTTAAAATCCCT |
| Forward Outer | GGCTGGAGCATTCGCATT |
| Reverse Outer | TTCTCCTGAAATTCGGGCAA |
| Forward Loop | AACATATTGTCCGTAGTGCG |
| Reverse Loop | TGATGATTTGTTATCGAGCAGC |
| Internal Amplification Control | |
| TEC primer/probe | (CYAN)TGTT(dSpacer)A(BHQ1-dT)ATCCGCGATCCTTGCGTTGT-TCCCCGCTATGGAAGGTC |
| Forward Inner | TGTTTATATCCGCGATCCTTGCGTTGT-TCCCCGCTATGGAAGGTC |
| Reverse Inner | CACCTGTTCGTGTCGTATCGGT-ATGCATTACCAGAGTGCTCC |
| Forward Outer | TACAGCGAAAAGCCCAGC |
| Reverse Outer | AAGCGACGAATGTCCTGTG |
| Forward Loop | TCTTAATTGCTTGCCGGAGC |
| Reverse Loop | GAGCCATGTGCCATACTCGTC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Higgins, O.; Clancy, E.; Cormican, M.; Boo, T.W.; Cunney, R.; Smith, T.J. Evaluation of an Internally Controlled Multiplex Tth Endonuclease Cleavage Loop-Mediated Isothermal Amplification (TEC-LAMP) Assay for the Detection of Bacterial Meningitis Pathogens. Int. J. Mol. Sci. 2018, 19, 524. https://doi.org/10.3390/ijms19020524
Higgins O, Clancy E, Cormican M, Boo TW, Cunney R, Smith TJ. Evaluation of an Internally Controlled Multiplex Tth Endonuclease Cleavage Loop-Mediated Isothermal Amplification (TEC-LAMP) Assay for the Detection of Bacterial Meningitis Pathogens. International Journal of Molecular Sciences. 2018; 19(2):524. https://doi.org/10.3390/ijms19020524
Chicago/Turabian StyleHiggins, Owen, Eoin Clancy, Martin Cormican, Teck Wee Boo, Robert Cunney, and Terry J. Smith. 2018. "Evaluation of an Internally Controlled Multiplex Tth Endonuclease Cleavage Loop-Mediated Isothermal Amplification (TEC-LAMP) Assay for the Detection of Bacterial Meningitis Pathogens" International Journal of Molecular Sciences 19, no. 2: 524. https://doi.org/10.3390/ijms19020524
APA StyleHiggins, O., Clancy, E., Cormican, M., Boo, T. W., Cunney, R., & Smith, T. J. (2018). Evaluation of an Internally Controlled Multiplex Tth Endonuclease Cleavage Loop-Mediated Isothermal Amplification (TEC-LAMP) Assay for the Detection of Bacterial Meningitis Pathogens. International Journal of Molecular Sciences, 19(2), 524. https://doi.org/10.3390/ijms19020524