LncRNA HOTTIP-Mediated HOXA11 Expression Promotes Cell Growth, Migration and Inhibits Cell Apoptosis in Breast Cancer

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
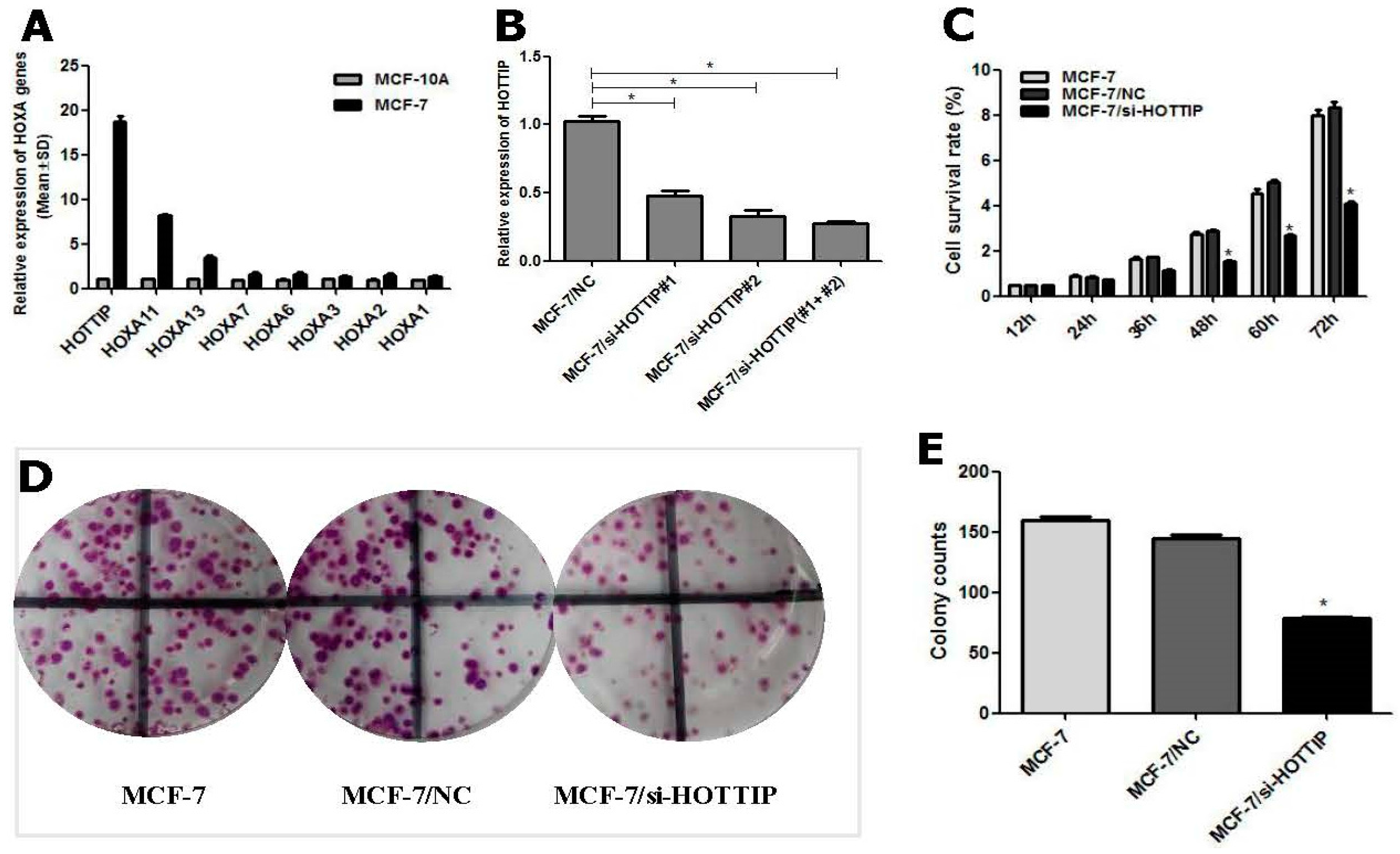
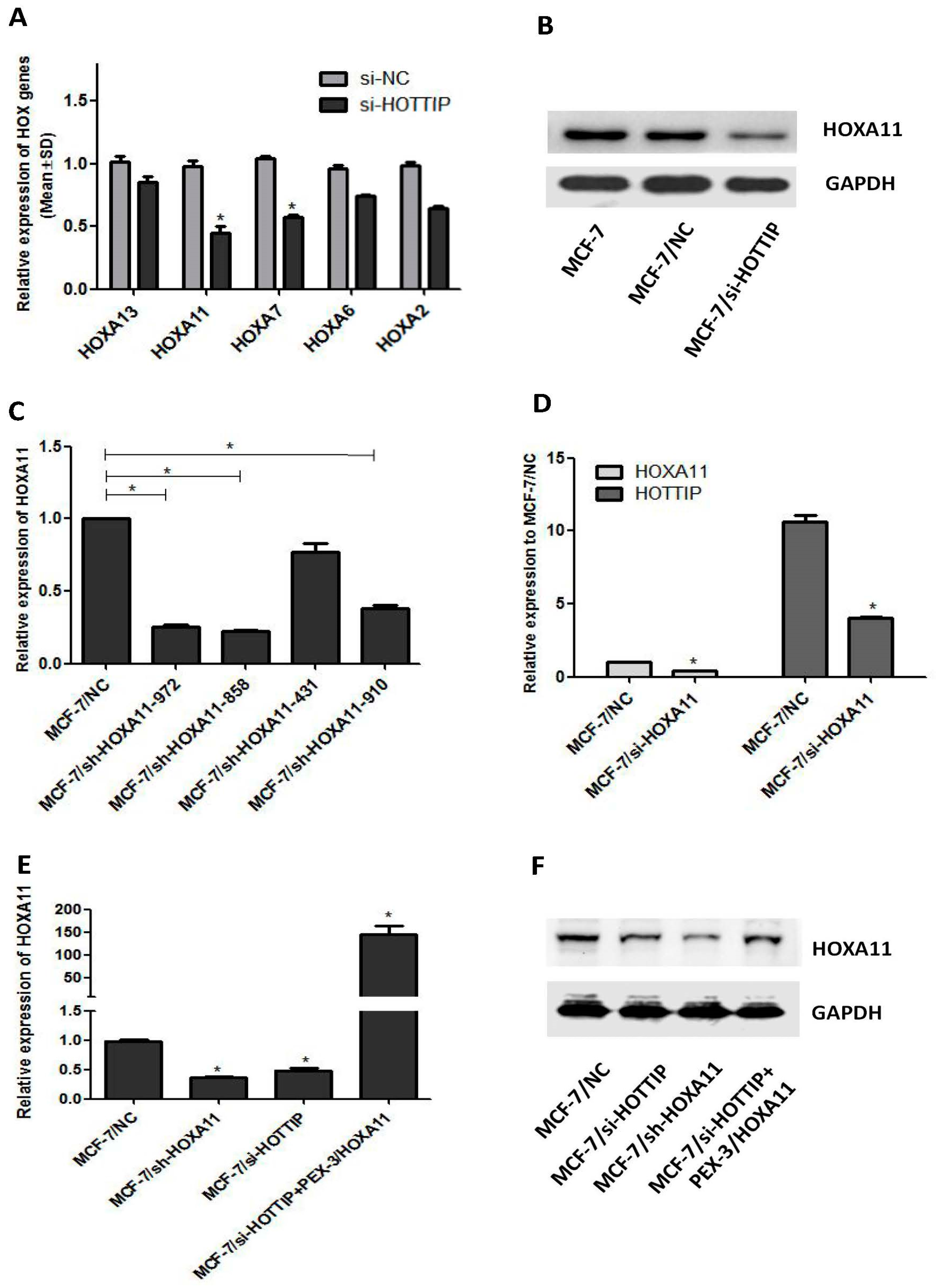
2.1. Determination of HOTTIP/HOXA11 Levels in Breast Cancer MCF-7 Cells
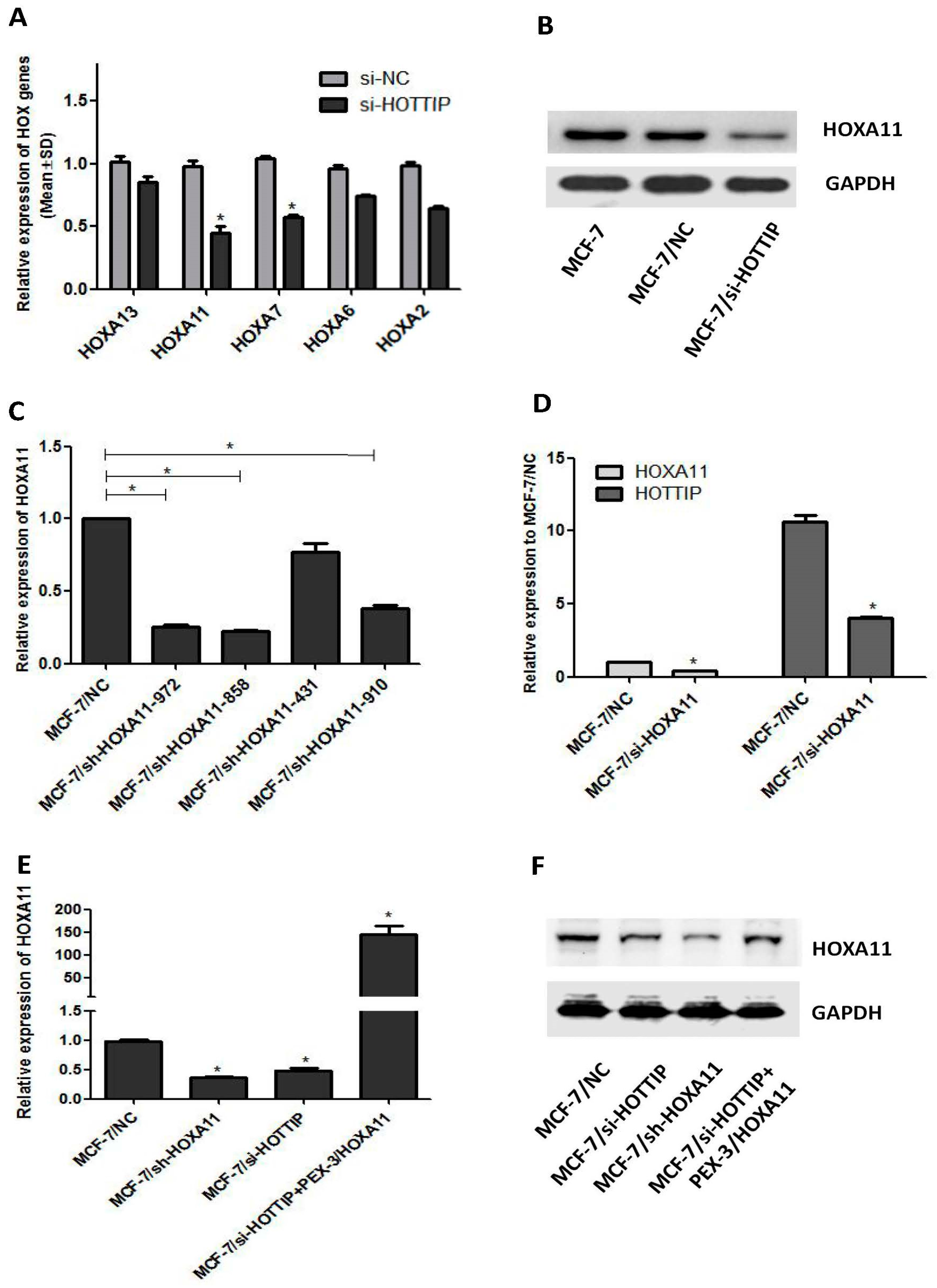
2.2. Screening of HOTTIP/HOXA11 Interference Sequences
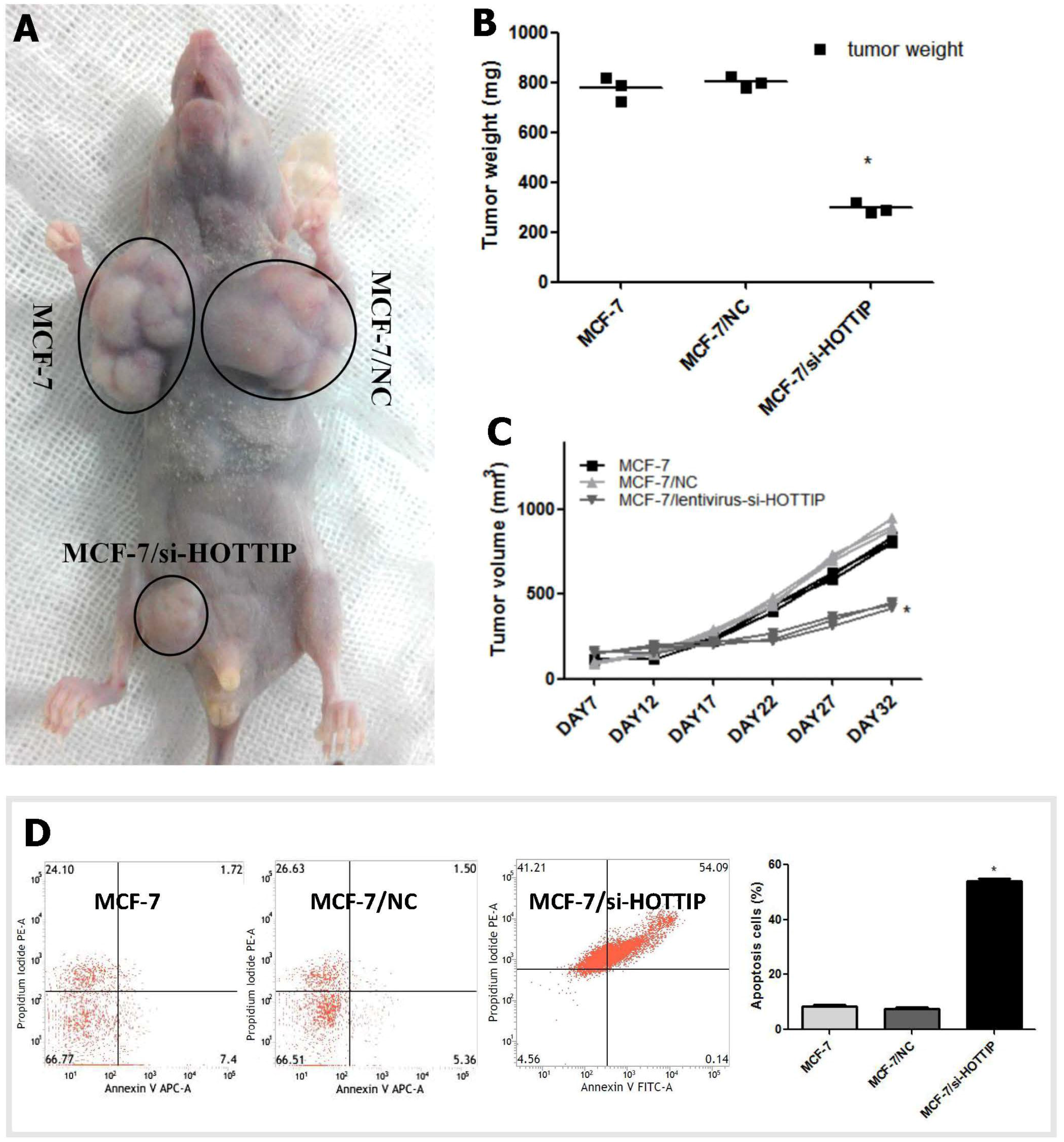
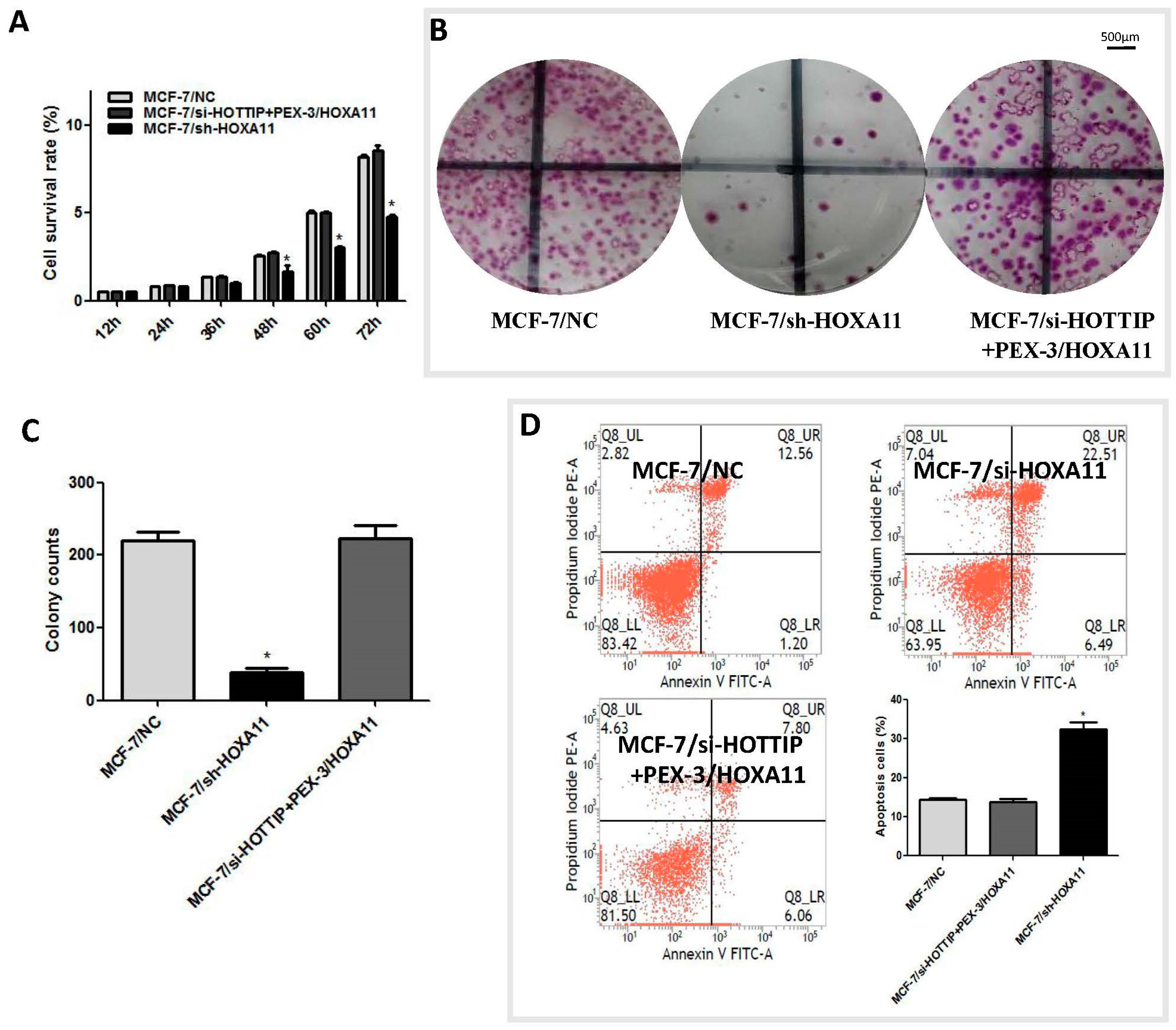
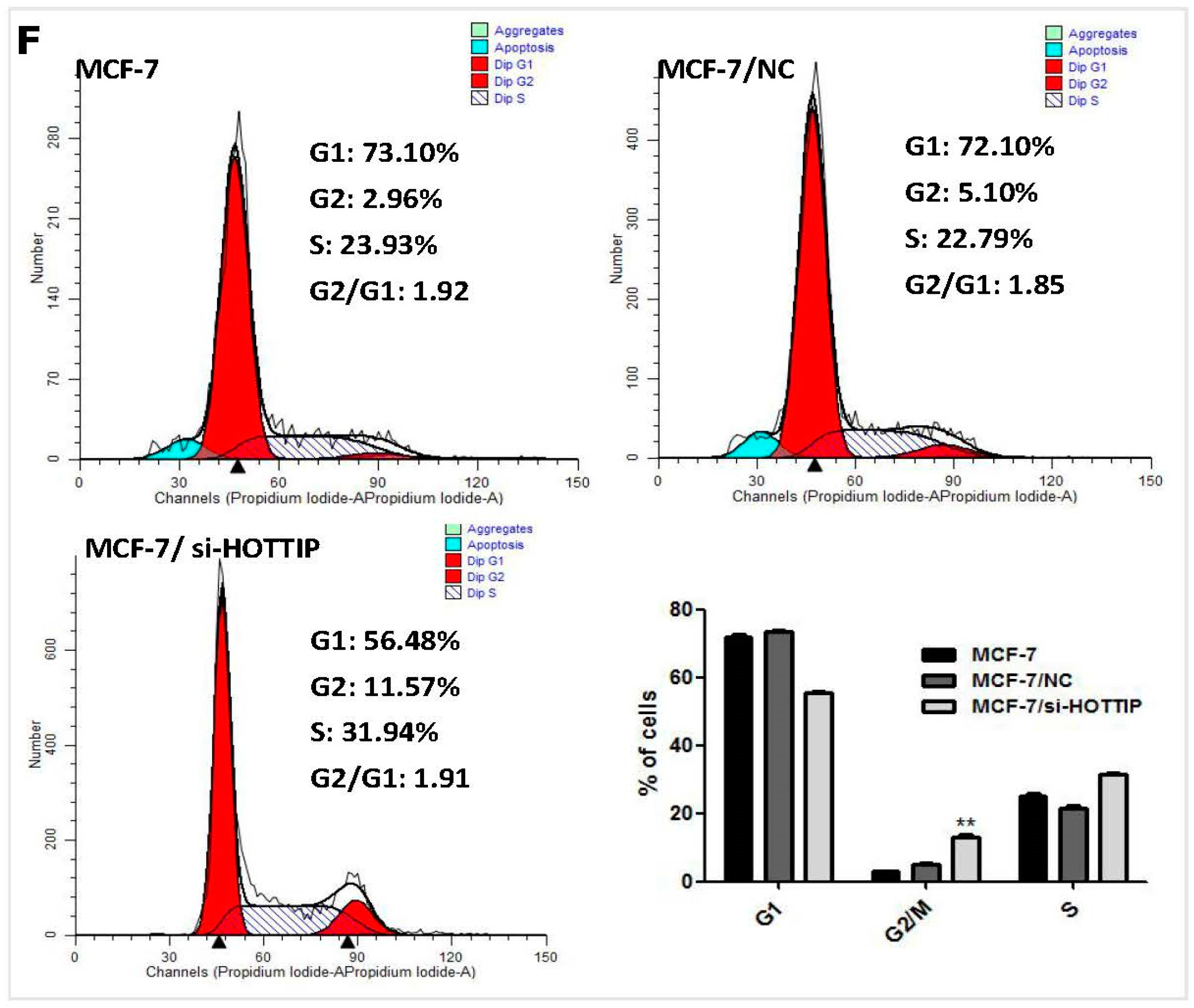
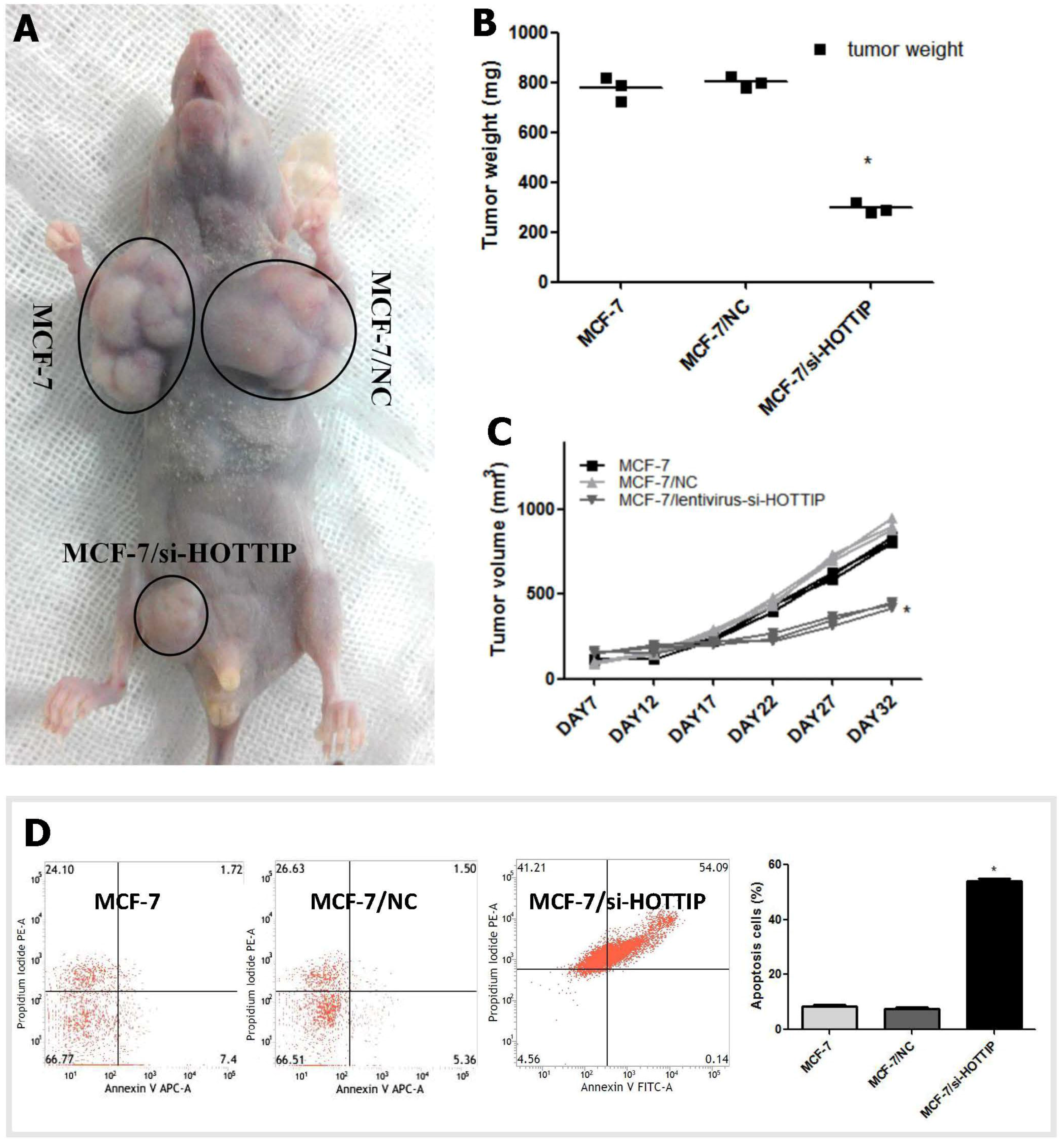
2.3. HOTTIP Regulates Breast Cancer Cell Growth In Vitro and In Vivo
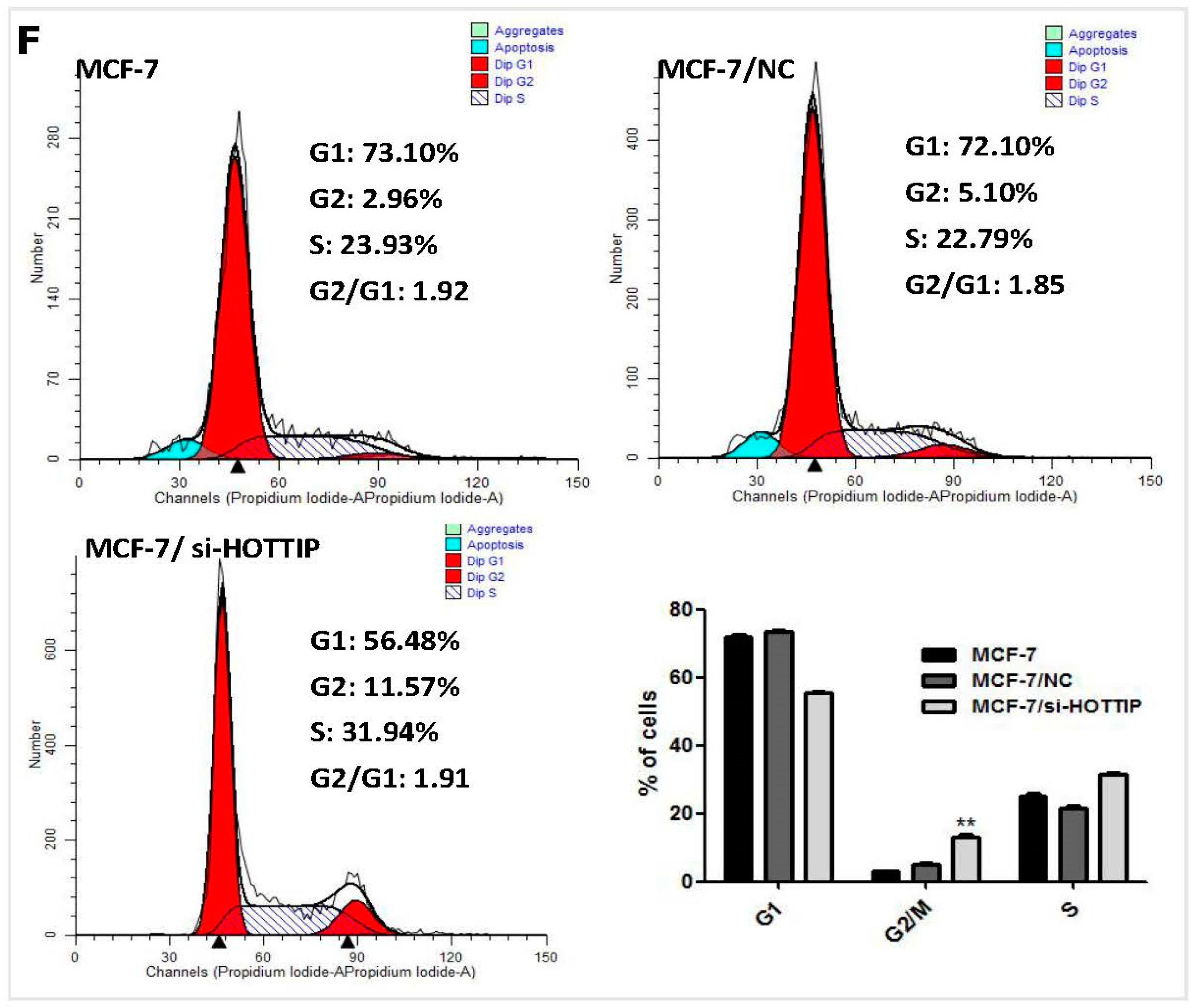
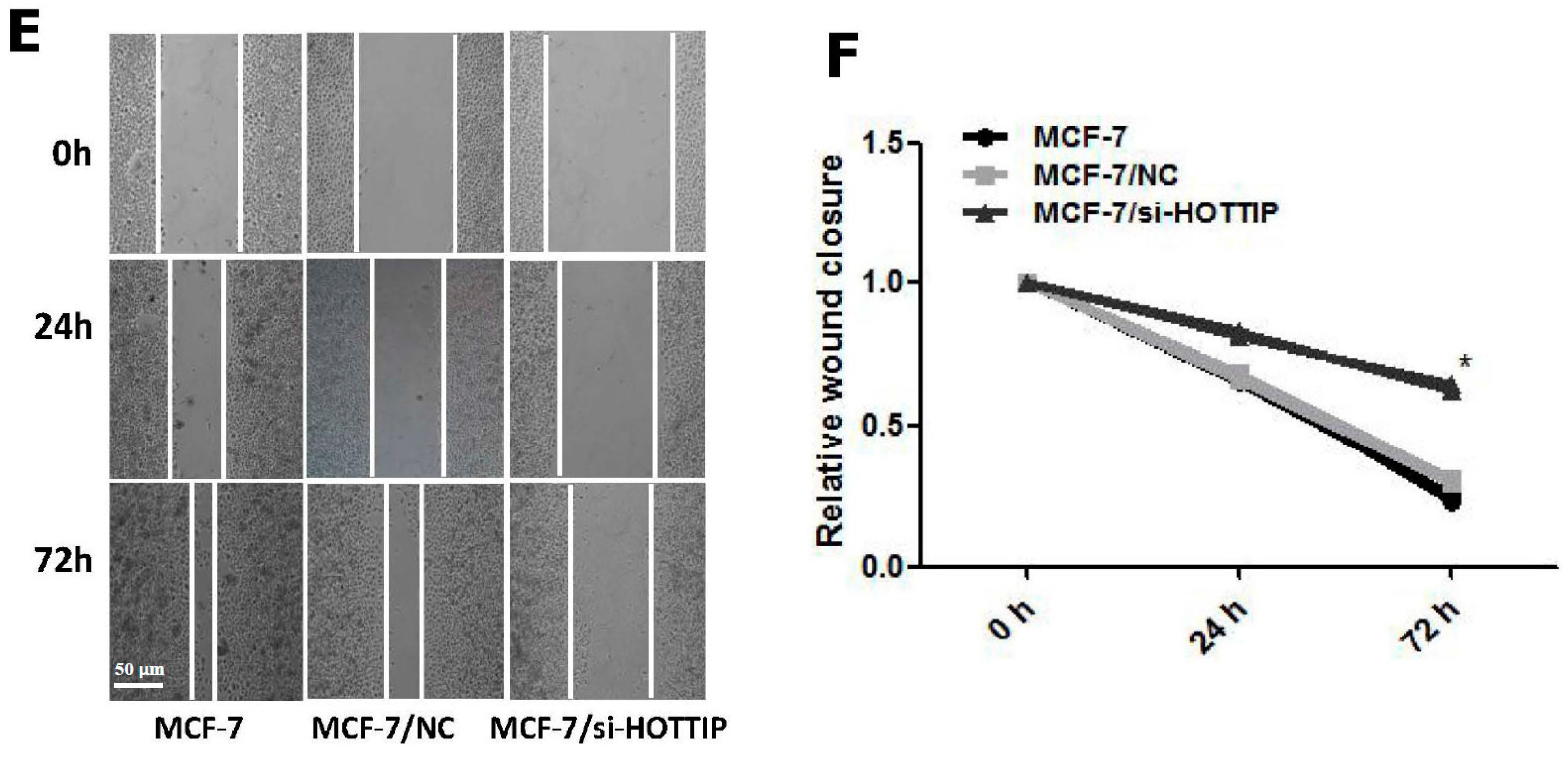
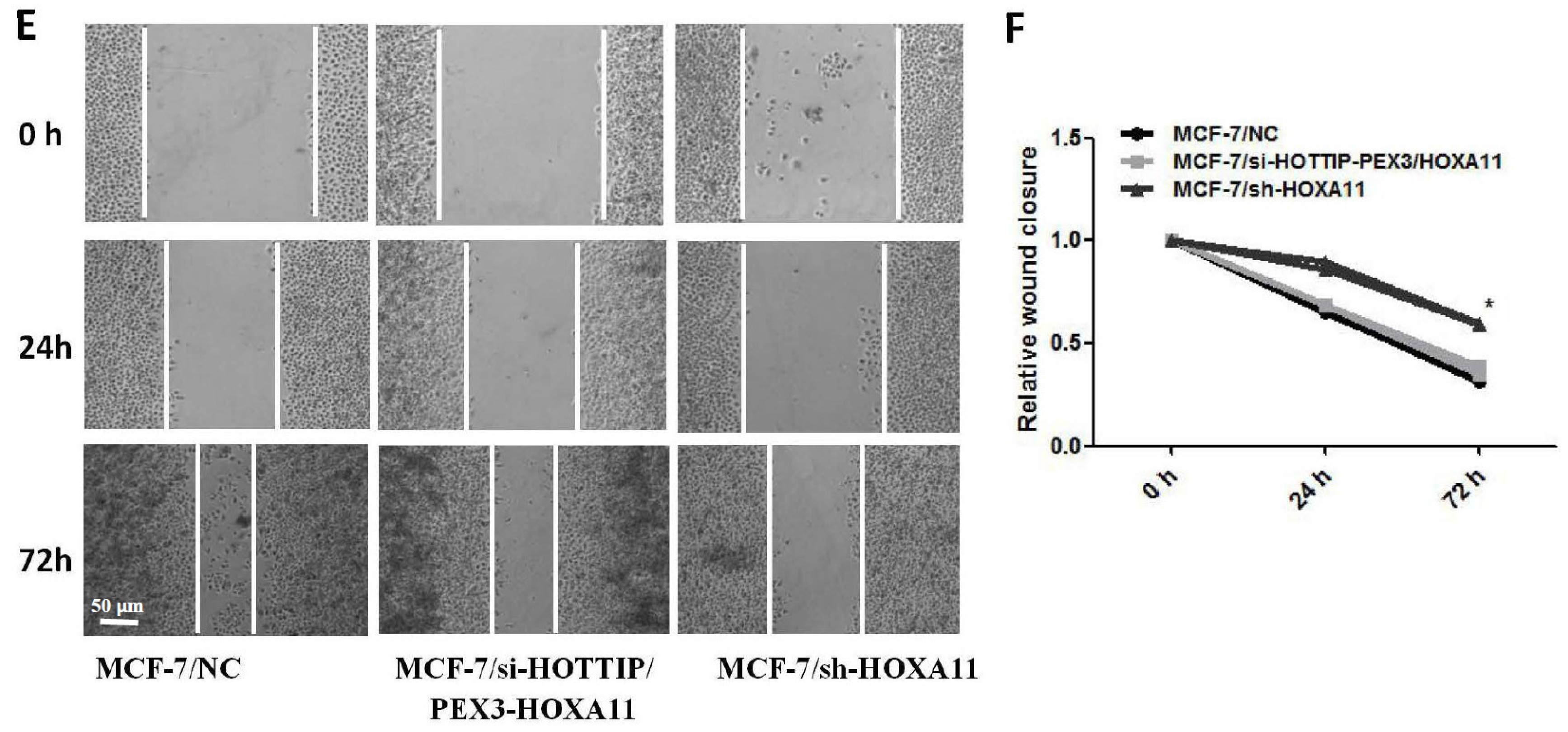
2.4. HOTTIP Suppresses Cell Apoptosis and Promotes Cell Migration In Vitro
2.5. A Potential Bidirectional Regulation between HOTTIP/HOXA11 in MCF-7 Cells
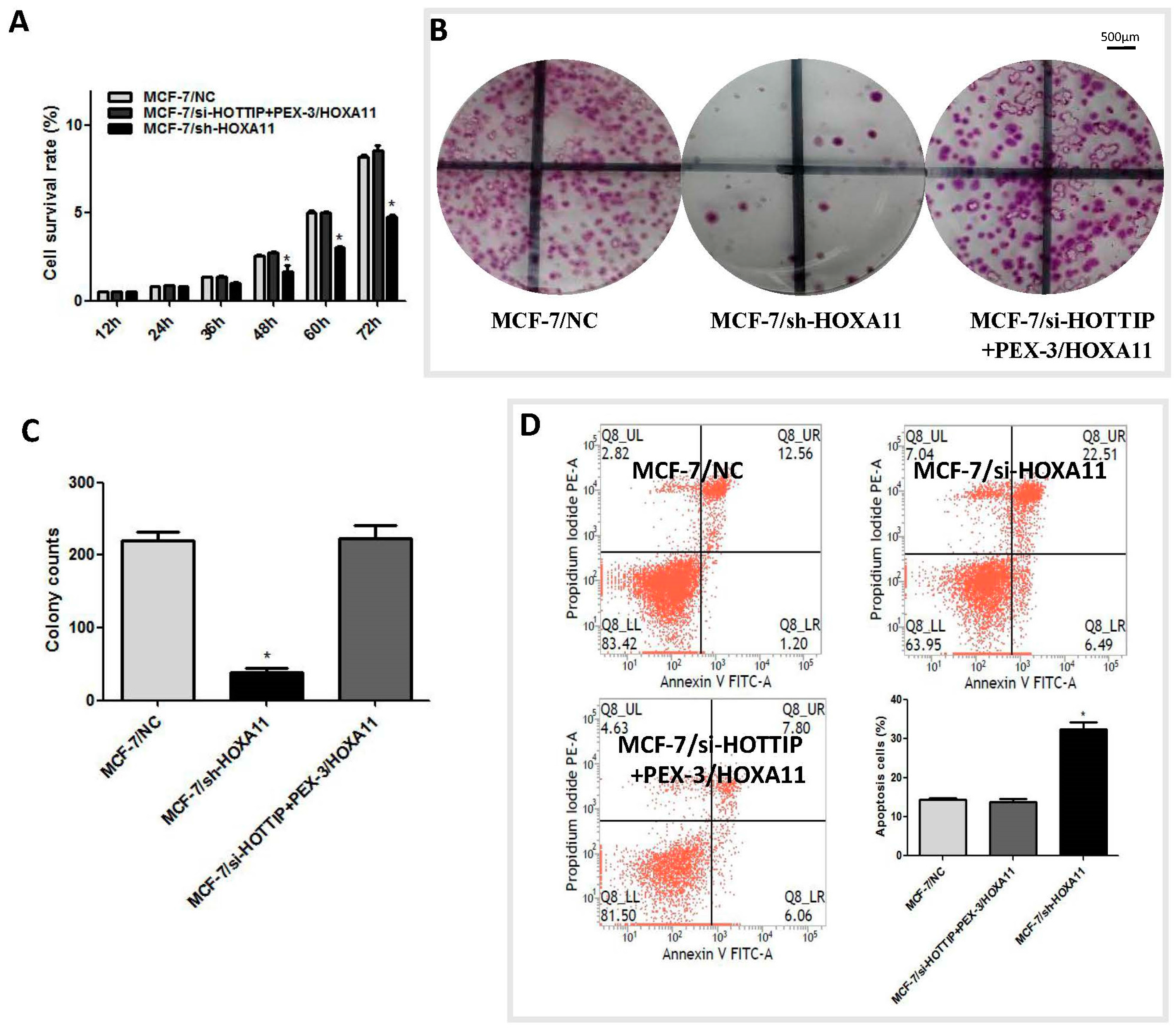
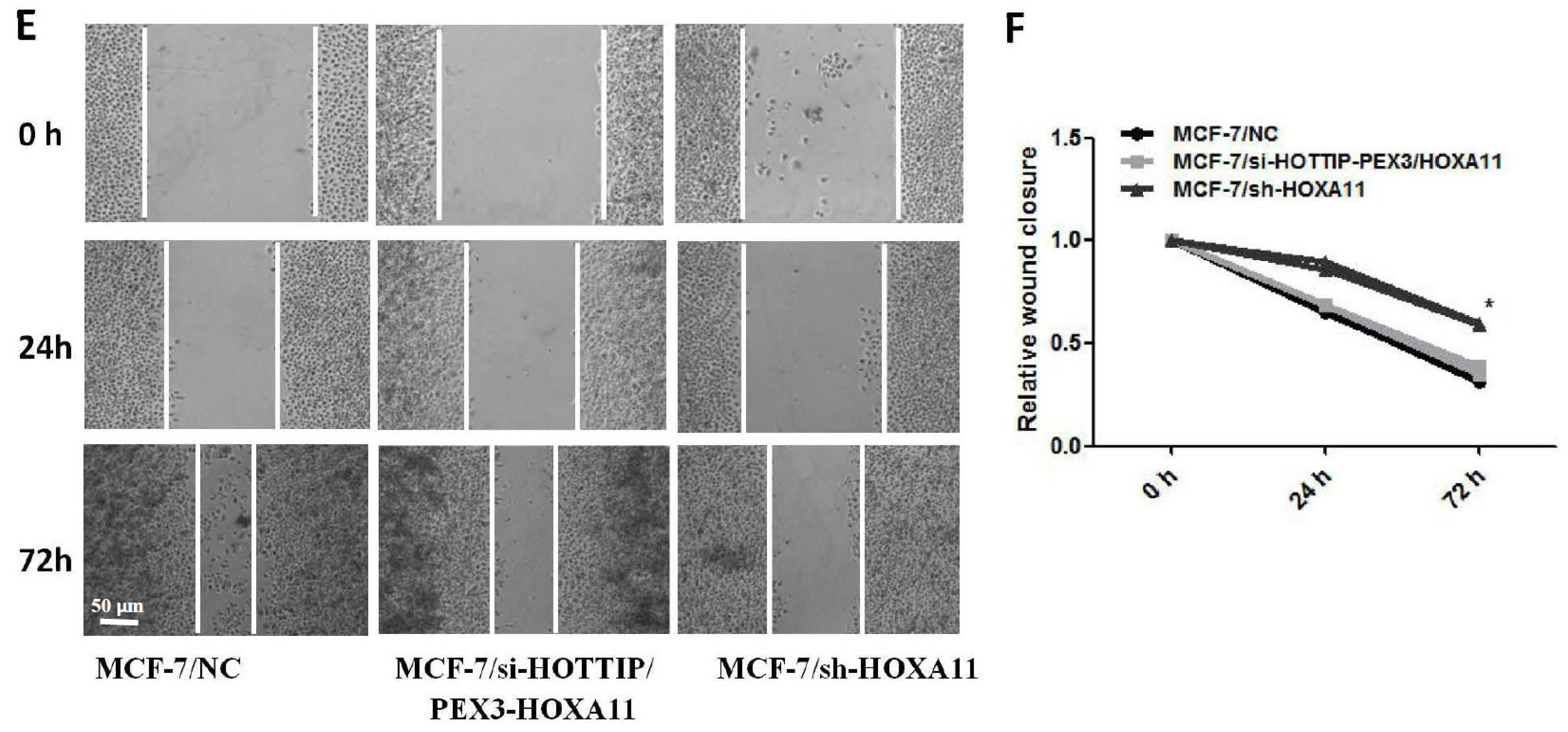
2.6. HOXA11 Partly Mediates the Effect of HOTTIP in Breast Cancer Pathogenesis
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Transient and Stable Transfection
4.3. Reverse Transcription Quantitative PCR (RT-qPCR)
4.4. Western Blot Analysis
4.5. Flow Cytometric Analysis for Cell Cycle and Apoptosis
4.6. In Vitro Proliferation Assay
4.7. In Vitro Wound Healing Experiment
4.8. In Vivo Xenograft Model in Nude Mice
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| lncRNAs | long non-coding RNAs |
| HOTTIP | HOXA transcript at the distal tip |
| RT-qPCR | Reverse transcription quantitative PCR |
| CCK-8 | cell counting kit 8 |
| WDR5 | WD repeat domain 5 |
| MLL | mixed lineage leukemia |
References
- Ribeiro Pereira, A.C.P.; Koifman, R.J.; Bergmann, A. Incidence and risk factors of lymphedema after breast cancer treatment: 10 years of follow-up. Breast 2017, 36, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Sidaway, P. Breast cancer: LAG3 expression indicates favourable outcomes. Nat. Rev. Clin. Oncol. 2017, 14, 712. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Lu, X.; Yuan, L. LncRNA: A link between RNA and cancer. Biochim. Biophys. Acta 2017, 1839, 1097–1109. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Koirala, P.; Mo, Y.Y. LncRNA-mediated regulation of cell signaling in cancer. Oncogene 2017, 36, 5661–5667. [Google Scholar] [CrossRef] [PubMed]
- Valadkhan, S.; Gunawardane, L.S. LncRNA-mediated regulation of the interferon response. Virus Res. 2016, 212, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.C.; Yang, Y.W.; Liu, B.; Sanyal, A.; Corces-Zimmerman, R.; Chen, Y.; Lajoie, B.R.; Protacio, A.; Flynn, R.A.; Gupta, R.A.; et al. A long noncoding RNA maintains active chromatin to coordinate homeotic gene expression. Nature 2011, 472, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Lian, Y.; Cai, Z.; Gong, H.; Xue, S.; Wu, D.; Wang, K. HOTTIP: A critical oncogenic long non-coding RNA in human cancers. Mol. Biosyst. 2016, 12, 3247–3253. [Google Scholar] [CrossRef] [PubMed]
- Quagliata, L.; Matter, M.S.; Piscuoglio, S.; Arabi, L.; Ruiz, C.; Procino, A.; Kovac, M.; Moretti, F.; Makowska, Z.; Boldanova, T.; et al. Long noncoding RNA HOTTIP/HOXA13 expression is associated with disease progression and predicts outcome in hepatocellular carcinoma patients. Hepatology 2014, 5, 911–923. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Qian, J.; Xiang, Y.; Chen, Y.; Qu, J. The prognostic value of long noncoding RNA HOTTIP on clinical outcomes in breast cancer. Oncotarget 2017, 8, 6833–6844. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.; Gao, H.; Tong, Y.; Yang, J.; Tang, R.; Niu, Y.; Guo, L. Long noncoding RNA-HOTAIR affects chemoresistance by regulating HOXA1 methylation in small cell lung cancer cells. Lab. Investig. 2016, 96, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Jia, X.; Wang, J.; Li, Y.; Xie, S. Knockdown of homeobox A5 by small hairpin RNA inhibits proliferation and enhances cytarabine chemosensitivity of acute myeloid leukemia cells. Mol. Med. Rep. 2015, 12, 6861–6866. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yang, S.; Su, N.; Wang, Y.; Yu, J.; Qiu, H.; He, X. Overexpression of long non-coding RNA HOTAIR leads to chemoresistance by activating the Wnt/β-catenin pathway in human ovarian cancer. Tumour Biol. 2016, 37, 2057–2065. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.; Li, Y.; Deng, H.; Zhang, C.; Pu, Y.; Qian, L.; Xiao, J.; Zhao, W.; Liu, Q.; Zhang, D.; et al. MiR-193a-3p promotes the multi-chemoresistance of bladder cancer by targeting the HOXC9 gene. Cancer Lett. 2015, 357, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Jutooru, I.; Chadalapaka, G.; Corton, J.C.; Safe, S. The long non-coding RNA HOTTIP enhances pancreatic cancer cell proliferation, survival and migration. Oncotarget 2015, 6, 10840–10852. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, N.; Kang, X.; Shi, K.; Chen, Q. Prognostic value of the long noncoding RNA HOTTIP in human cancers. Oncotarget 2017, 8, 59563–59569. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Lin, J.; Li, Y.; Zhang, Y.; Chen, X. Prognostic role of lncRNA TUG1 for cancer outcome: Evidence from 840 cancer patients. Oncotarget 2017, 8, 50051–50060. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Chen, C.; Zhou, Q.; Wang, Y.; Zhao, Y.; Zhao, X.; Li, W.; Zheng, S.; Ye, H.; Wang, L.; et al. LncRNA HOTTIP modulates cancer stem cell properties in human pancreatic cancer by regulating HOXA9. Cancer Lett. 2017, 410, 68–81. [Google Scholar] [CrossRef] [PubMed]
- Malek, R.; Gajula, R.P.; Williams, R.D.; Nghiem, B.; Simons, B.W.; Nugent, K.; Wang, H.; Taparra, K.; Lemtiri-Chlieh, G.; Yoon, A.R.; et al. TWIST1-WDR5-Hottip Regulates HOXA9 Chromatin to Facilitate Prostate Cancer Metastasis. Cancer Res. 2017, 77, 3181–3193. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.; Liu, J.; Guo, S.; He, S.; Qiu, G.; Lu, J.; Wang, J.; Fan, L.; Zhao, W.; Che, X. HOTTIP and HOXA13 are oncogenes associated with gastric cancer progression. Oncol. Rep. 2016, 35, 3577–3585. [Google Scholar] [CrossRef] [PubMed]
- Richards, E.J.; Permuth-Wey, J.; Li, Y.; Chen, Y.A.; Coppola, D.; Reid, B.M.; Lin, H.Y.; Teer, J.K.; Berchuck, A.; Birrer, M.J.; et al. A functional variant in HOXA11-AS, a novel long non-coding RNA, inhibits the oncogenic phenotype of epithelial ovarian cancer. Oncotarget 2015, 6, 34745–34757. [Google Scholar] [CrossRef] [PubMed]
- Kallianos, A.; Rapti, A.; Zarogoulidis, P.; Tsakiridis, K.; Mpakas, A.; Katsikogiannis, N.; Kougioumtizi, I.; Li, Q.; Huang, H.; Zaric, B.; et al. Therapeutic procedure in small cell lung cancer. J. Thorac. Dis. 2013, 5, S420–S442. [Google Scholar] [PubMed]
- Levesley, J.; Steele, L.; Bruning-Richardson, A.; Davison, A.; Zhou, J.; Ding, C.; Lawler, S.; Short, S.C. Selective BCL-XL inhibition promotes apoptosis in combination with MLN8237 in medulloblastoma and pediatric glioblastoma cells. Neuro Oncol. 2017, 20, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhou, Y.; Bai, Y.; Wang, Q.; Bao, J.; Luo, Y.; Guo, Y.; Guo, L. A long non-coding RNA HOTTIP expression is associated with disease progression and predicts outcome in small cell lung cancer patients. Mol. Cancer 2017, 16, 162. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.S.; Li, X.M.; Yang, M.; Ren, X.L.; Hu, J.L.; Zhu, X.H.; Wang, F.F.; Zeng, Z.C.; Li, J.Y.; Cheng, Z.Q.; et al. FMNL2 destabilises COMMD10 to activate NF-κB pathway in invasion and metastasis of colorectal cancer. Br. J. Cancer 2017, 117, 1164–1175. [Google Scholar] [CrossRef] [PubMed]
- Su, F.; Li, H.; Yan, C.; Jia, B.; Zhang, Y.; Chen, X. Depleting MEKK1 expression inhibits the ability of invasion and migration of human pancreatic cancer cells. J. Cancer Res. Clin. Oncol. 2009, 135, 1655–1663. [Google Scholar] [CrossRef] [PubMed]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Zeng, C.; Gan, S.; Li, H.; Cheng, Y.; Chen, D.; Li, R.; Zhu, W. LncRNA HOTTIP-Mediated HOXA11 Expression Promotes Cell Growth, Migration and Inhibits Cell Apoptosis in Breast Cancer. Int. J. Mol. Sci. 2018, 19, 472. https://doi.org/10.3390/ijms19020472
Sun Y, Zeng C, Gan S, Li H, Cheng Y, Chen D, Li R, Zhu W. LncRNA HOTTIP-Mediated HOXA11 Expression Promotes Cell Growth, Migration and Inhibits Cell Apoptosis in Breast Cancer. International Journal of Molecular Sciences. 2018; 19(2):472. https://doi.org/10.3390/ijms19020472
Chicago/Turabian StyleSun, Yanqin, Chao Zeng, Siyuan Gan, Hongmei Li, Ying Cheng, Dongjie Chen, Rujia Li, and Wei Zhu. 2018. "LncRNA HOTTIP-Mediated HOXA11 Expression Promotes Cell Growth, Migration and Inhibits Cell Apoptosis in Breast Cancer" International Journal of Molecular Sciences 19, no. 2: 472. https://doi.org/10.3390/ijms19020472