Comparison of the Abundance and Community Structure of N-Cycling Bacteria in Paddy Rhizosphere Soil under Different Rice Cultivation Patterns


Abstract
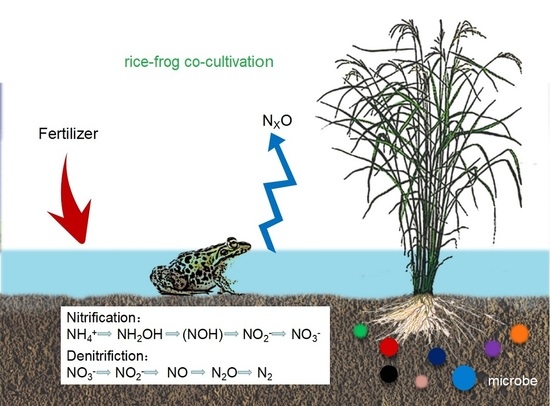
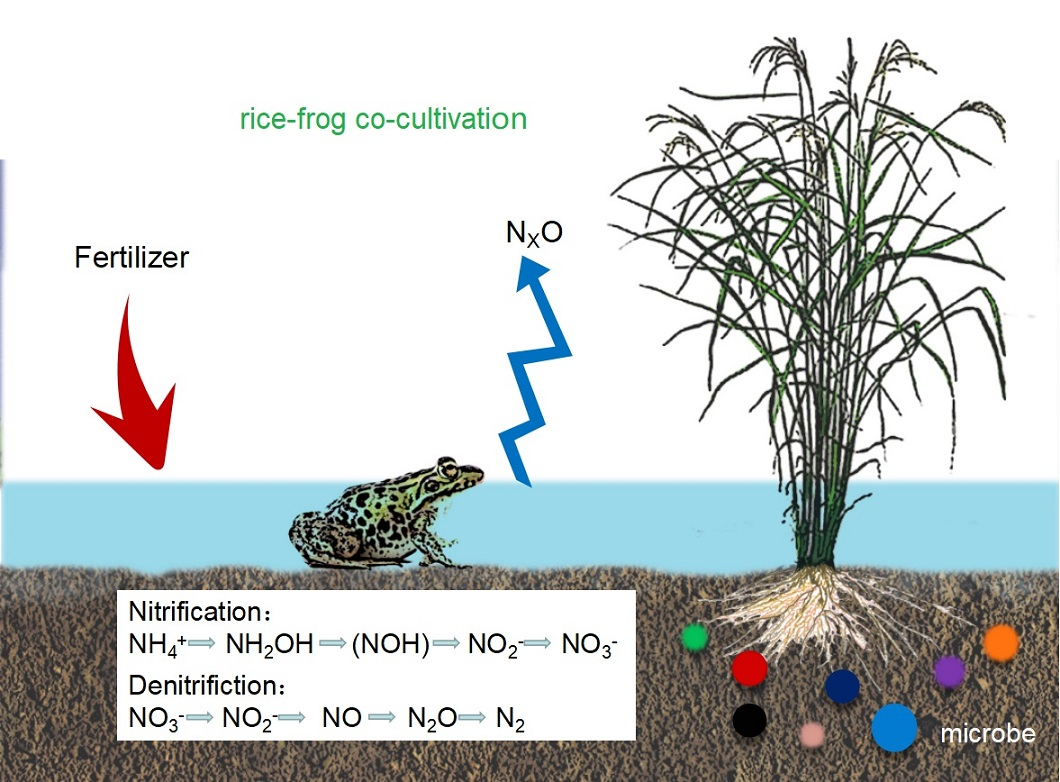
:
1. Introduction
2. Results
2.1. Soil Enzyme Activities
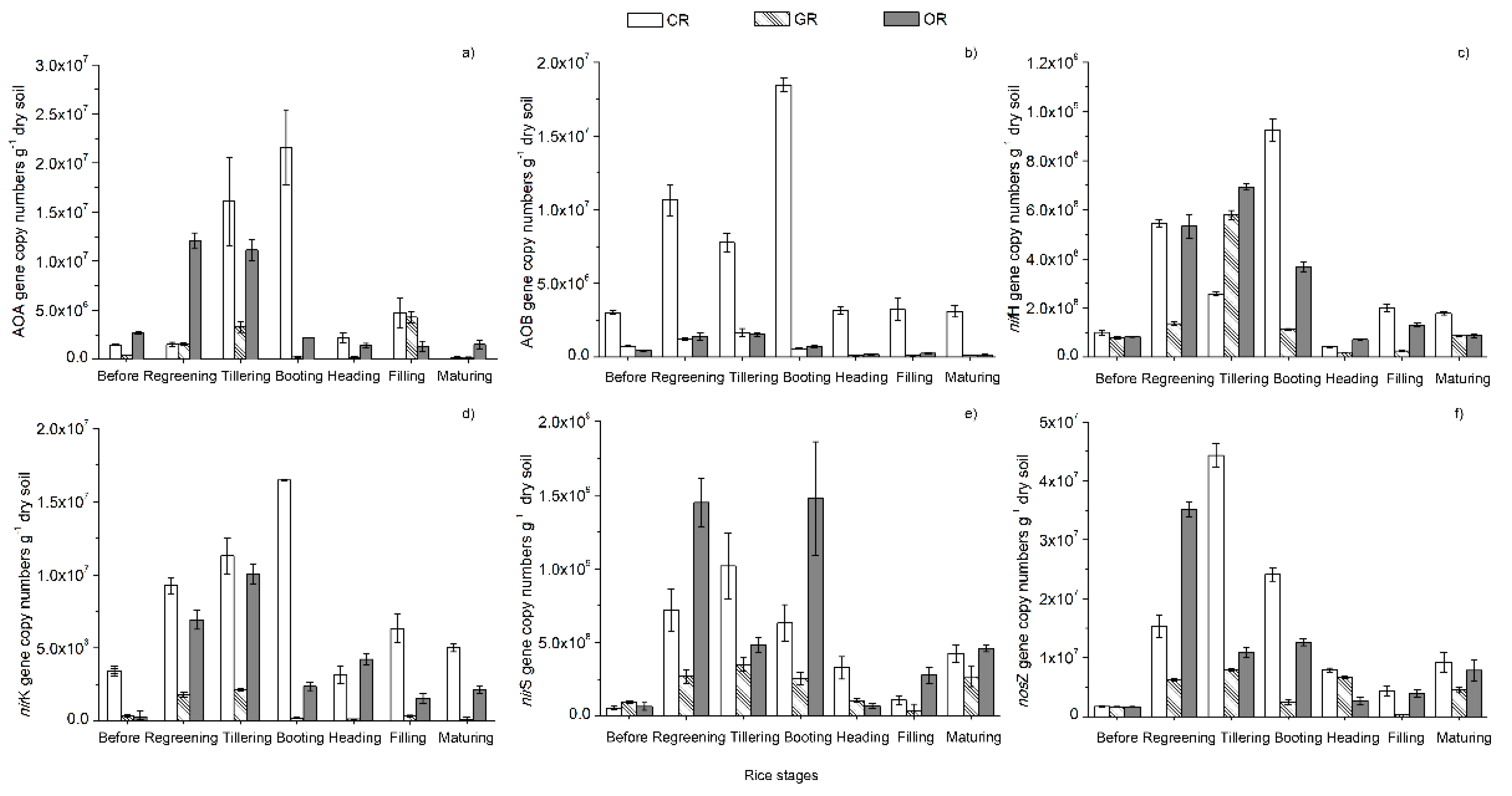
2.2. Quantification of N-cycling Genes
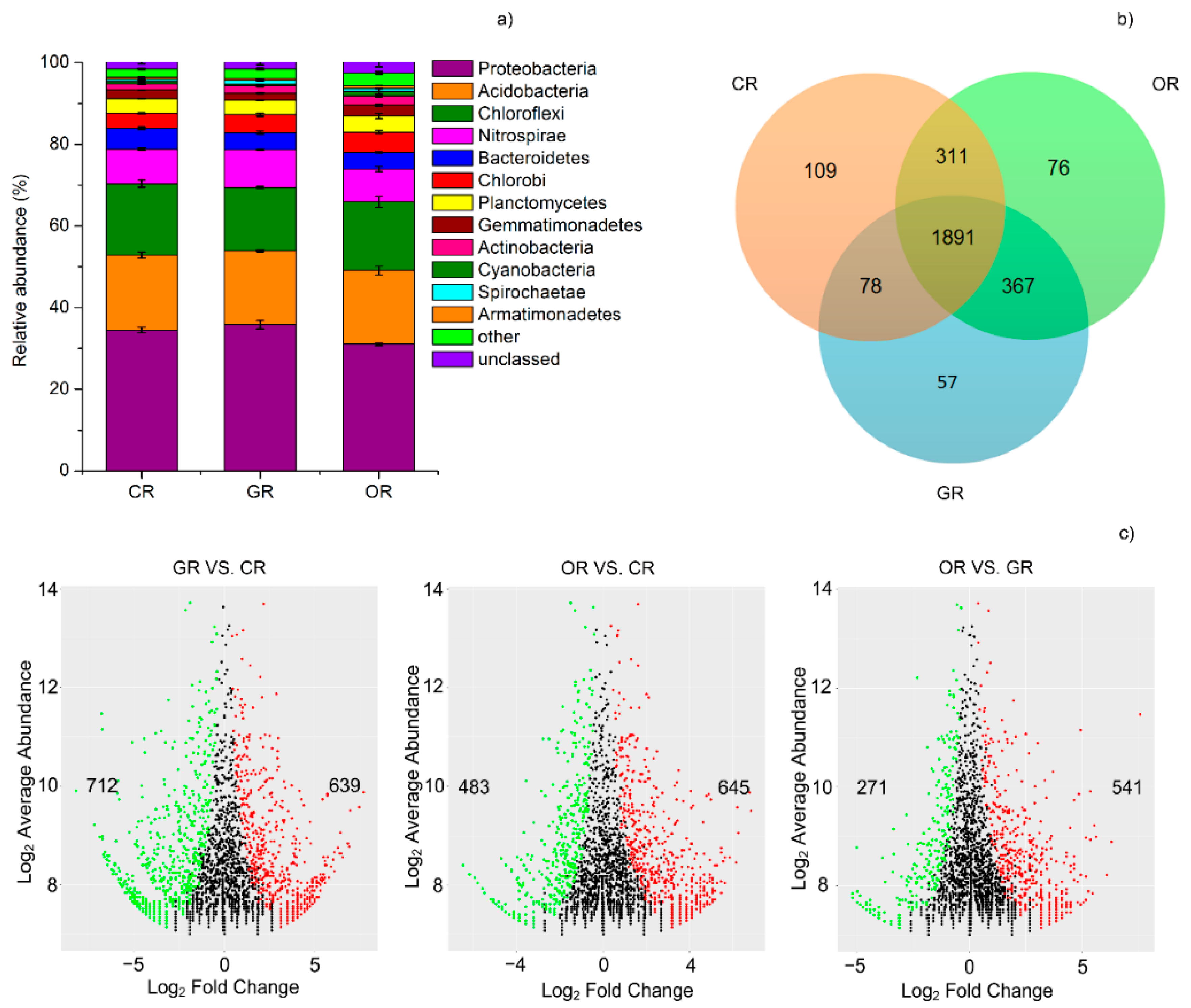
2.3. Bacterial Community Structure in Paddy Rhizosphere Soil
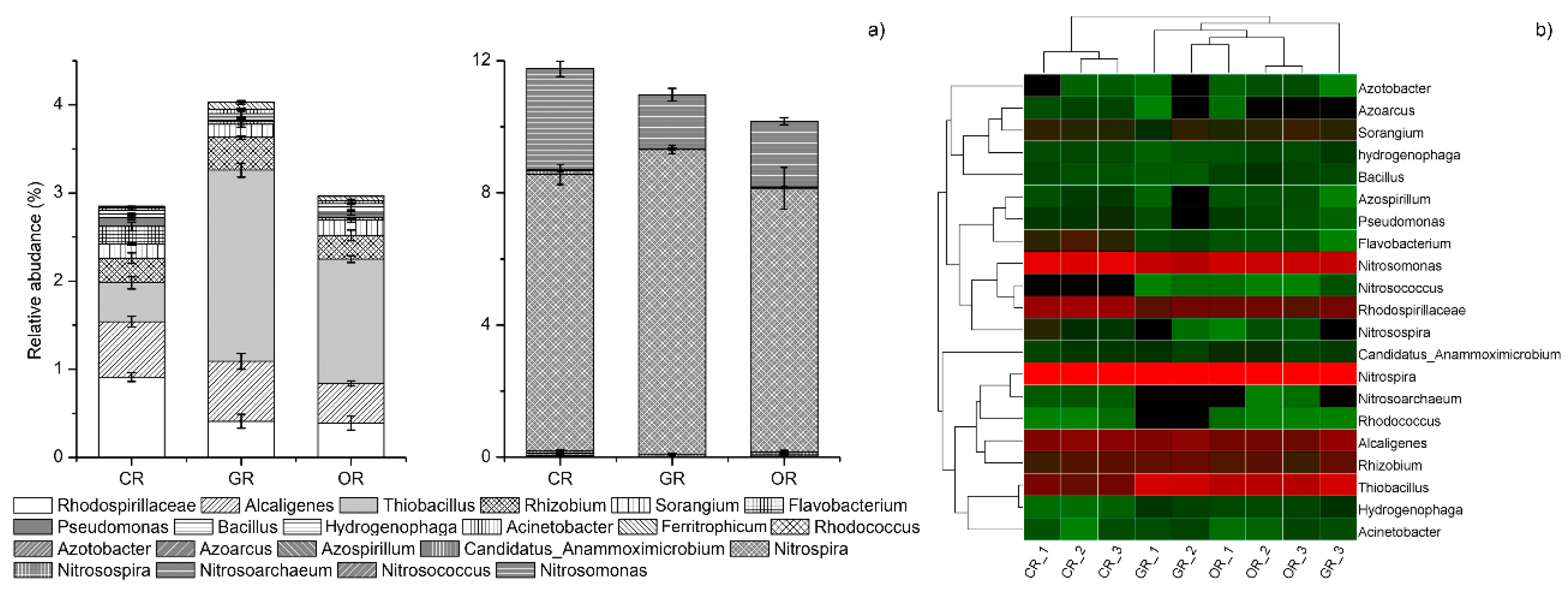
2.4. N-cycling Bacterial Community
3. Materials and Methods
3.1. Experimental Site
3.2. Experimental Design
3.3. Sampling and Measurements
3.3.1. Soil
3.3.2. Plant Harvesting and Grain Analysis
3.4. DNA Extraction
3.5. Quantitative PCR
3.5.1. PCR Amplification of Target Genes
3.5.2. Standard Plasmid and Standard Curve
3.6. MiSeq Sequencing of 16S rRNA and Bioinformatics Analysis
3.7. Statistical Analysis
4. Discussion
4.1. Different Rice Cultivation Patterns Drive Soil Enzymes Change
4.2. Soil N-Cycling Genes of Different Rice Cultivation Patterns
4.3. Microbial Community Structure Change among Different Rice Cultivation Patterns
4.4. N-Cycling Bacterial Community Abundance Was Effected by Rice Cultivation Pattern
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Ma, B.; Uphoff, N. A review of the system of rice intensification in china. Plant. Soil. 2015, 393, 361–381. [Google Scholar] [CrossRef]
- FAO, Food and Agriculture Organization of the United Nations Statistics Division (FAOSTAT). 2018. Available online: http://www.fao.org/faostat/zh/#data (accessed on 15 March 2018).
- Hu, X.K.; Su, F.; Ju, X.T.; Gao, B.; Oenema, O.; Christie, P.; Huang, B.X.; Jiang, R.F.; Zhang, F.S. Greenhouse gas emissions from a wheat-maize double cropping system with different nitrogen fertilization regimes. Environ. Pollut. 2013, 176, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Gu, B.; Schlesinger, W.H. Significant accumulation of nitrate in Chinese semi-humid croplands. Sci. Rep. 2016, 6, 25088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, Y.; Stewart, B.A.; Zhang, F.S. Long-term experiments for sustainable nutrient management in China. A review. Agron. Sustain. Dev. 2010, 31, 397–414. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.H.; Liu, X.J.; Zhang, Y.; Shen, J.L.; Han, W.X.; Zhang, W.F.; Christie, P.; Goulding, K.W.; Vitousek, P.M.; Zhang, F.S. Significant acidification in major chinese croplands. Science 2010, 327, 1008–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mae, T. Nitrogen acquisition and its relation to growth and yield in recent high-yielding cultivars of rice (Oryza sativa L.) in Japan. J. Soil. Sci. Plant. Nut. 2011, 57, 625–635. [Google Scholar] [CrossRef]
- Li, E.H.; Li, W.; Liu, G.H.; Yuan, L.Y. The effect of different submerged macrophyte species and biomass on sediment resuspension in a shallow freshwater lake. Aquatic Botany. 2008, 88, 121–126. [Google Scholar] [CrossRef]
- Xie, J.; Hu, L.L.; Tang, J.J.; Wu, X.; Li, N.N.; Yuan, Y.G.; Yang, H.S.; Zhang, J.E.; Luo, S.M.; Chen, X. Ecological mechanisms underlying the sustainability of the agricultural heritage rice-fish coculture system. Proc. Natl. Acad. Sci. USA 2011, 108, E1381–1387. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.L.; Zhang, J.; Ren, W.Z.; Guo, L.; Cheng, Y.X.; Li, J.Y.; Li, K.X.; Zhu, Z.W.; Zhang, J.E.; Luo, S.M.; et al. Can the co-cultivation of rice and fish help sustain rice production? Sci. Rep. 2016, 6, 28728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granstedt, A.; Schneider, T.; Seuri, P.; Thomsson, O. Ecological Recycling Agriculture to Reduce Nutrient Pollution to Baltic Sea. Biol. Agric. Hortic. 2008, 26, 279–307. [Google Scholar] [CrossRef]
- Stein-Bachinger, K.; Reckling, M.; Bachinger, J.; Hufnagel, J.; Koker, W.; Granstedt, A. Ecological Recycling Agriculture to Enhance Agro-Ecosystem Services in the Baltic Sea Region: Guidelines for Implementation. Land. 2015, 4, 737–753. [Google Scholar] [CrossRef] [Green Version]
- Li, C.F.; Cao, C.G.; Wang, J.P.; Ming, Z.; Yuan, W.L.; Ahmad, S. Nitrogen losses from integrated rice-duck and rice-fish ecosystems in southern China. Plant. Soil. 2008, 307, 207–217. [Google Scholar]
- Zheng, H.B.; Huang, H.; Li, D.Q.; Li, X.M.; Fu, Z.Q.; Chen, C. Assessment of nomadic rice-duck complex ecosystem on energy and economy. Ecol. Proc. 2014, 3, 20. [Google Scholar] [CrossRef]
- Ren, W.Z.; Hu, L.L.; Guo, L.; Zhang, J.; Tang, L.; Zhang, E.T.; Zhang, J.E.; Luo, S.M.; Tang, J.J.; Chen, X. Preservation of the genetic diversity of a local common carp in the agricultural heritage rice-fish system. Proc. Natl. Acad. Sci. USA 2018, 115, E546–E554. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.S.; Yu, D.G.; Zhou, J.J.; Zhai, S.L.; Bian, X.M.; Weih, M. Rice-duck co-culture for reducing negative impacts of biogas slurry application in rice production systems. J. Environ. Manage. 2018, 213, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Teng, Q.; Hu, X.F.; Cheng, C.; Luo, Z.Q.; Luo, F.; Xue, Y.; Jiang, Y.J.; Mu, Z.; Liu, L.L.; Yang, M.Y. Ecological effects of rice-duck integrated farming on soil fertility and weed and pest control. J. Soil. Sediment. 2016, 16, 2395–2407. [Google Scholar] [CrossRef]
- Zhou, H.B.; Chen, J.L.; Cheng, D.F.; Francis, F.; Liu, Y.; Sun, J.R. Effects of ecological regulation of biodiversity on insects in agroecosystems. Plant. Prot. 2012, 38, 6–47. [Google Scholar]
- Tsuji, M.; Ushimaru, A.; Osawa, T.; Mitsuhashi, H. Paddy-associated frog declines via urbanization: A test of the dispersal-dependent-decline hypothesis. Landscape. Urban. Plan. 2011, 103, 318–325. [Google Scholar] [CrossRef]
- Ben, T.F.; Fdhila, K.; Elabed, H.; Bouguerra, A.; Kouidhi, B.; Bakhrouf, A.; Chaieb, K. Molecular identification of potential denitrifying bacteria and use of d-optimal mixture experimental design for the optimization of denitrification process. Microb. Pathog. 2016, 93, 158–165. [Google Scholar]
- Hamer, A.J.; Makings, J.A.; Lane, S.J.; Mahony, M.J. Amphibian decline and fertilizers used on agricultural land in south-eastern Australia. Agr. Ecosyst. Environ. 2004, 102, 299–305. [Google Scholar] [CrossRef]
- Sha, Z.M.; Chu, Q.N.; Zhao, Z.; Yue, Y.B.; Lu, L.F.; Yuan, J.; Cao, L.K. Variations in nutrient and trace element composition of rice in an organic rice-frog coculture system. Sci. Rep. 2017, 7, 15706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tringe, S.G.; Mering, C.V.; Kobayashi, A.; Asaf, A.; Chen, K.; Chang, H.W.; Podar, M.; Short, J.M.; Mathur, E.J.; Detter, J.C.; Bork, P.; Hugenholtz, P.; Rubin, E.M. Comparative metagenomics of microbial communities. Science 2005, 308, 554–557. [Google Scholar] [CrossRef] [PubMed]
- Edwards, J.; Johnson, C.; Santos-Medellin, C.; Lurie, E.; Podishetty, N.K.; Bhatnagar, S.; Eisen, J.A.; Sundaresan, V. Structure, variation, and assembly of the root-associated microbiomes of rice. Proc. Natl. Acad. Sci. USA 2015, 112, E911–E920. [Google Scholar] [CrossRef] [PubMed]
- Sims, A.; Zhang, Y.; Gajaraj, S.; Brown, P.B.; Hu, Z. Toward the development of microbial indicators for wetland assessment. Water Res. 2013, 47, 1711–1725. [Google Scholar] [CrossRef] [PubMed]
- Urakawa, H.; Bernhard, A.E. Wetland management using microbial indicators. Ecol. Eng. 2017, 108, 456–476. [Google Scholar] [CrossRef]
- Nannipieri, P.; Eldor, P. The chemical and functional characterization of soil n and its biotic components. Soil Biol. Biochem. 2010, 42, 863. [Google Scholar] [CrossRef]
- Petersen, D.G.; Blazewicz, S.J.; Firestone, M.; Herman, D.J.; Turetsky, M.; Waldrop, M. Abundance of microbial genes associated with nitrogen cycling as indices of biogeochemical process rates across a vegetation gradient in Alaska. Environ. Microbiol. 2012, 14, 993–1008. [Google Scholar] [CrossRef] [PubMed]
- Wolińska, A.; Kuzńiar, A.; Zielenkiewicz, U.; Banach, A.; Izak, D.; Stepniewska, Z.; Błaszczyk, M. Metagenomic analysis of some potential nitrogen-fixing bacteria in arable soils at different formation processes. Microb. Ecol. 2017, 73, 162–176. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.H.; Nasir, M.; Lv, J.; Dai, Y.C.; Gao, J.K. Understanding the variation of microbial community in heavy metals contaminated soil using high throughput sequencing. Ecotoxicol. Environ. Saf. 2017, 144, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Yue, Y.B.; Sha, Z.M.; Li, C.S.; Deng, J.; Zhang, H.L.; Gao, M.F.; Cao, L.K. Assessing impacts of alternative fertilizer management practices on both nitrogen loading and greenhouse gas emissions in rice cultivation. Atmos. Environ. 2015, 119, 393–401. [Google Scholar] [CrossRef]
- Yuan, J.; Sha, Z.M.; Hassani, D.; Zhao, Z.; Cao, L.K. Assessing environmental impacts of organic and inorganic fertilizer on daily and seasonal Greenhouse Gases effluxes in rice field. Atmos. Environ. 2017, 155, 119–128. [Google Scholar] [CrossRef]
- Yuan, J.; Yuan, Y.K.; Zhu, Y.H.; Cao, L.K. Effects of different fertilizers on methane emissions and methanogenic community structures in paddy rhizosphere soil. Sci. Total. Environ. 2018, 627, 770–781. [Google Scholar] [CrossRef] [PubMed]
- Shen, R.F.; Cai, H.; Gong, W.H. Transgenic Bt cotton has no apparent effect on enzymatic activities or functional diversity of microbial communities in rhizosphere soil. Plant Soil. 2006, 285, 149–159. [Google Scholar] [CrossRef]
- Xiong, L.; Xu, Z.F.; Wu, F.Z.; Yang, W.Q.; Yin, R.; Li, Z.P.; Gou, X.L.; Tang, S.S. Effects of snow pack on soil nitrogen transformation enzyme activities in a subalpine abies faxioniana forest of western sichuan, china. CHN. J. Appl. Ecol. 2014, 25, 1293–1299. [Google Scholar]
- Francis, C.A.; Roberts, K.J.; Beman, J.M.; Santoro, A.E.; Oakley, B.B. Ubiquity and diversity of ammonia-oxidizing archaea in water columns and sediments of the ocean. Proc. Natl. Acad. Sci. USA 2005, 102, 14683–14688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotthauwe, J.H.; Witzel, K.P.; Liesack, W. The ammonia monooxygenase structural gene amoA as a functional marker: Molecular fine-scale analysis of natural ammonia-oxidizing populations. Appl. Environ. Microb. 1997, 63, 4704–4712. [Google Scholar]
- Rosch, C.; Mergel, A.; Bothe, H. Biodiversity of Denitrifying and Dinitrogen-Fixing Bacteria in an Acid Forest Soil. Appl. Environ. Microb. 2002, 68, 3818–3829. [Google Scholar] [CrossRef] [Green Version]
- Throback, I.N.; Enwall, K.; Jarvis, A.; Hallin, S. Reassessing PCR primers targeting nirS, nirK and nosZ genes for community surveys of denitrifying bacteria with DGGE. FEMS. Microbiol. Ecol. 2004, 49, 401–417. [Google Scholar] [CrossRef] [PubMed]
- Henry, S.; Bru, D.; Stres, B.; Hallet, S.; Philippot, L. Quantitative detection of the nosZ gene, encoding nitrous oxide reductase, and comparison of the abundances of 16s rRNA, narG, nirK, and nosZ genes in soils. Appl. Environ. Microbiol. 2006, 72, 5181–5189. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.X.; Yi, M.; Yi, H.L.; Guo, E.H.; Zhang, E.Y. Manure and mineral fertilization change enzyme activity and bacterial community in millet rhizosphere soils. World J. Microbiol. Biotechnol. 2017, 34, 8. [Google Scholar] [CrossRef] [PubMed]
- Magoc, T.; Salzberg, S.L. FLASH: fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microb. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I. QIIME allows analysis of high-throughput community sequencing data. Nat. Method. 2010, 7, 335. [Google Scholar] [CrossRef] [PubMed]
- Burns, R.G.; Deforest, J.L.; Marxsen, J.; Sinsabaugh, R.L.; Stromberger, M.E.; Wallenstein, M.D.; Weintraub, M.N.; Zoppinih, A. Soil enzymes in a changing environment: Current knowledge and future directions. Soil Biol. Biochem. 2013, 58, 216–234. [Google Scholar] [CrossRef]
- Kotroczó, Z.; Veres, Z.; Fekete, I.; Krakomperger, Z.; Tóth, J.A.; Lajtha, K.; Lajtha, K.; Tóthmérész, B. Soil enzyme activity in response to long-term organic matter manipulation. Soil Biol. Biochem. 2014, 70, 237–243. [Google Scholar] [CrossRef]
- Jian, S.Y.; Li, J.W.; Chen, J.; Wang, G.S.; Mayes, M.A.; Dzantor, K.E.; Hui, D.F.; Luo, Y.Q. Soil extracellular enzyme activities, soil carbon and nitrogen storage under nitrogen fertilization: A meta-analysis. Soil Biol. Biochem. 2016, 101, 32–43. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Li, D.J.; Zhao, J.; Xiao, K.C.; Wang, K.L. Effects of nitrogen addition on activities of soil nitrogen acquisition enzymes: A meta-analysis. Agr. Ecosyst. Environ. 2018, 252, 126–131. [Google Scholar] [CrossRef]
- López-Aizpún, M.; Arango-Mora, C.; Santamaría, C.; Lasheras, E.; Santamaría, J.M.; Ciganda, V.S.; Cárdenas, L.M.; Elustondo, D. Atmospheric ammonia concentration modulates soil enzyme and microbial activity in an oak forest affecting soil microbial biomass. Soil Biol. Biochem. 2018, 116, 378–387. [Google Scholar] [CrossRef]
- Moro, E.; Crusciol, C.A.C.; Nascente, A.S.; Cantarella, H.; Broetto, F.; Moro, A.L. Nitrate Reductase, Micronutrients and Upland Rice Development as Influenced by Soil pH and Nitrogen Sources. Commun. Soil. Sci. Plan. 2017, 48, 2642–2651. [Google Scholar]
- Chen, Z.; Luo, X.Q.; Hu, R.G.; Wu, M.N.; Wu, J.S.; Wei, W.X. Impact of Long-Term Fertilization on the Composition of Denitrifier Communities Based on Nitrite Reductase Analyses in a Paddy Soil. Microb. Ecol. 2010, 60, 850–861. [Google Scholar] [CrossRef] [PubMed]
- Khalil, K.; Renault, P.; Guérin, N.; Mary, B. Modelling denitrification including the dynamics of denitrifiers and their progressive ability to reduce nitrous oxide: comparison with batch experiments. Eur. J. Soi. Sci. 2005, 56, 491–504. [Google Scholar] [CrossRef]
- Hoffman, B.M.; Lukoyanov, D.; Dean, D.R.; Seefeldt, L.C. Nitrogenase: A Draft Mechanism. Accounts Chem. Res. 2013, 46, 587–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duval, S.; Danyal, K.; Shaw, S.; Lytle, A.K.; Dean, D.R.; Hoffman, B.M.; Antony, E.; Seefeldt, L.C. Electron transfer precedes ATP hydrolysis during nitrogenase catalysis. Proc. Natl. Acad. Sci. USA 2013, 110, 16414–16419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, X.Y.; Hurek, T.; Reinhold-Hurek, B. Effect of N-fertilization, plant genotype and environmental conditions on nifH gene pools in roots of rice. Environ. Microbiol. 2003, 5, 1009–1015. [Google Scholar] [CrossRef] [PubMed]
- Coelho, M.R.R.; Marriel, I.E.; Jenkins, S.N.; Lanyon, C.V.; Seldin, L.; O’Donnell, A.G. Molecular detection and quantification of nifH gene sequences in the rhizosphere of sorghum (Sorghum bicolor) sown with two levels of nitrogen fertilizer. Appl. Soil. Ecol. 2009, 42, 48–53. [Google Scholar] [CrossRef]
- Fierer, N.; Lauber, C.L.; Ramirez, K.S.; Zaneveld, J.; Bradford, M.A.; Knight, R. Comparative metagenomic, phylogenetic and physiological analyses of soil microbial communities across nitrogen gradients. ISME. J. 2012, 6, 1007–1017. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.Y.; Park, S.J.; Min, D.; Kim, J.S.; Rijpstra, W.I.; Sinninghe Damste, J.S.; Kim, G.J.; Madsen, E.L.; Rhee, S.K. Enrichment and characterization of an autotrophic ammonia-oxidizing archaeon of mesophilic crenarchaeal group I.1a from an agricultural soil. Appl. Environ. Microbiol. 2011, 77, 8635–8647. [Google Scholar] [CrossRef] [PubMed]
- Kerou, M.; Offre, P.; Valledor, L.; Abby, S.S.; Melcher, M.; Nagler, M.; Weckwerth, W.; Schleper, C. Proteomics and comparative genomics of Nitrososphaera viennensis reveal the core genome and adaptations of archaeal ammonia oxidizers. Proc Natl Acad Sci USA 2016, 113, E7937–E7946. [Google Scholar] [CrossRef] [PubMed]
- Di, H.J.; Cameron, K.C.; Shen, J.P.; Winefield, C.S.; He, J.Z. Nitrification driven by bacteria and not archaea in nitrogen-rich grassland soils. Nat. Geosci. 2009, 2, 621–624. [Google Scholar] [CrossRef]
- Levy-Booth, D.J.; Prescott, C.E.; Grayston, S.J. Microbial functional genes involved in nitrogen fixation, nitrification and denitrification in forest ecosystems. Soil Biol. Biochem. 2014, 75, 11–25. [Google Scholar] [CrossRef]
- Harter, J.; Krause, H.M.; Schuettler, S.; Ruser, R.; Fromme, M.; Scholten, T.; Kappler, A.; Behrens, S. Linking N2O emissions from biochar-amended soil to the structure and function of the N-cycling microbial community. ISME. J. 2014, 8, 660–674. [Google Scholar] [CrossRef] [PubMed]
- Teng, Q.; Hu, X.F.; Luo, F.; Cheng, C.; Ge, X.Y.; Yang, M.Y.; Liu, L.M. Influences of introducing frogs in the paddy fields on soil properties and rice growth. J. Soil. Sediment. 2015, 16, 51–61. [Google Scholar] [CrossRef]
- Sánchez, O. Constructed Wetlands Revisited: Microbial Diversity in the -omics Era. Microb. Ecol. 2017, 73, 722–733. [Google Scholar] [CrossRef] [PubMed]
- Demoling, F.; Figueroa, D.; Baath, E. Comparison of factors limiting bacterial growth in different soils. Soil Biol. Biochem. 2007, 39, 2485–2495. [Google Scholar] [CrossRef]
- Xia, L.L.; Lam, S.K.; Yan, X.Y.; Chen, D.L. How Does Recycling of Livestock Manure in Agroecosystems Affect Crop Productivity, Reactive Nitrogen Losses, and Soil Carbon Balance? Environ. Sci. Techno. 2017, 51, 7450–7457. [Google Scholar] [CrossRef] [PubMed]
- Sachdeva, V.; Hooda, V. Immobilization of nitrate reductase onto epoxy affixed silver nanoparticles for determination of soil nitrates. Int. J. Biol. Macromol. 2015, 79, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Banning, N.C.; Maccarone, L.D.; Fisk, L.M.; Murphy, D.V. Ammonia-oxidising bacteria not archaea dominate nitrification activity in semi-arid agricultural soil. Sci. Rep. 2015, 5, 11146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirk, G.J.; Kronzucker, H.J. The potential for nitrification and nitrate uptake in the rhizosphere of wetland plants: a modelling study. Ann. Bot. 2005, 96, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.J.; Zhang, J.B.; Cai, Z.C. Nitrification activities and N mineralization in paddy soils are insensitive to oxygen concentration. Acta. Agr. Scand. B-S. P. 2015, 66, 272–281. [Google Scholar] [CrossRef]
- Hayatsu, M.; Tago, K.; Uchiyama, I.; Toyoda, A.; Wang, Y.; Shimomura, Y.; Okubo, T.; Kurisu, F.; Hirono, Y.; Nonaka, K.; et al. An acid-tolerant ammonia-oxidizing gamma-proteobacterium from soil. ISME. J. 2017, 11, 1130–1141. [Google Scholar] [CrossRef] [PubMed]
- Tao, R.; Wakelin, S.A.; Liang, Y.; Hu, B.; Chu, G. Nitrous oxide emission and denitrifier communities in drip-irrigated calcareous soil as affected by chemical and organic fertilizers. Sci. Total. Environ. 2018, 612, 739–749. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pattern | Grain Protein Content (%) | Plant N Content (%) | Dry Matter Biomass (g plant−1) | Rice Yield (kg ha−1) |
|---|---|---|---|---|
| CR | 7.44 ± 0.06 b | 1.05 ± 0.01 b | 84.56 ± 3.23 a | 8827.56 a |
| GR | 9.72 ± 0.42 a | 1.35 ± 0.02 a | 86.95 ± 4.12 a | 8650.38 a |
| OR | 7.94 ± 0.10 b | 0.97 ± 0.02 b | 74.14 ± 5.02 b | 7350.69 b |
| UEA | PTA | AOA | AOB | nirK | nifH | nirS | Plant N | |
|---|---|---|---|---|---|---|---|---|
| URA | - | - | - | - | - | - | - | - |
| PRO | - | - | - | - | - | - | - | - |
| NRA | - | - | - | - | - | - | - | - |
| NiRA | - | 0.52 * | - | - | - | - | - | - |
| AOA | 0.61 ** | - | - | - | - | - | - | - |
| AOB | 0.66 ** | - | 0.68 ** | - | - | - | - | - |
| nirK | 0.81 ** | - | 0.85 ** | 0.85 ** | - | - | - | - |
| nifH | 0.61 ** | - | 0.72 ** | 0.67 ** | 0.78 ** | - | - | - |
| nirS | 0.62 ** | - | 0.47 * | - | 0.45 * | 0.53 * | - | - |
| nosZ | 0.68 ** | 0.44 * | 0.77 ** | 0.53 * | 0.70 ** | 0.53 * | 0.78 ** | - |
| Plant N | 0.84 ** | - | - | - | - | - | - | - |
| Grain Pro | −0.73 * | - | - | - | - | - | - | 0.94 ** |
| Rice yield | - | −0.67 * | - | 0.83 ** | - | - | - | - |
| Index | Chao 1 | ACE | Shannon | InvSimpson |
|---|---|---|---|---|
| CR | 2391.55 ± 17.52 b | 2396.35 ± 25.90 b | 6.58 ± 0.03 b | 293.07 ± 5.38 c |
| GR | 2398.68 ± 60.50 b | 2397.42 ± 77.19 b | 6.61 ± 0.02 b | 326.91 ± 2.52 b |
| OR | 2654.24 ± 27.03 a | 2646.10 ± 30.40 a | 6.81 ± 0.03 a | 406.75 ± 18.23 a |
| NO3−-N | NH4+-N | PH | EC | TC | TN | C:N | AK | AP | AN | N1 | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| NO3−-N | |||||||||||
| NH4+-N | −0.88 ** | ||||||||||
| PH | −0.84 ** | ||||||||||
| TC | 0.96 ** | 0.90 ** | |||||||||
| TN | 0.70 * | 0.67 * | |||||||||
| C: N | 0.84 ** | 0.91 ** | |||||||||
| AK | 0.95 ** | −0.92 ** | −0.88 ** | ||||||||
| AP | 0.73 * | −0.93 ** | −0.91 ** | 0.77 * | |||||||
| AHN | −0.91 ** | 0.98 ** | 0.93 ** | −0.92 ** | −0.94 ** | ||||||
| N1 | −0.76 * | 0.92** | 0.94 ** | 0.86 ** | −0.70 * | ||||||
| N2 | 0.72 * | −0.80 * | −0.81 ** | 0.75 * | 0.74 * | −0.75 * | |||||
| N3 | 0.89 ** | 0.95 ** | 0.74 * | 0.80 ** | 0.87 ** |
| Patterns | Pre-transplanting | Jointing Stage | Heading Stage | |||
|---|---|---|---|---|---|---|
| Chinese milk vetch | rapeseed cake | bulk blending fertilizer | bio-organic fertilizer | bulk blending fertilizer | urea | |
| CR | - | - | 150 | - | 75 | 75 |
| GR | 22.5 | 127.5 | - | 75 | 75 | |
| OR | 22.5 | 127.5 | - | 150 | - | - |
| Rice Stage | Day | Dates | Major Farming Management Practices |
|---|---|---|---|
| Pre-transplanting | 31 | 14 May–14 Jun. | Ploughed Chinese milk vetch in GR and OR (May 14), applied 100% rapeseed cake in GR and OR, 67% bulk blending fertilizer in CR (Jun. 14). |
| Regreening | 15 | 15 Jun.–30 Jun. | |
| Tillering | 21 | 1 Jul.–22 Jul. | Put tiger frogs into fields (6000 frogs ha−1, Jul. 1). Applied 33% bulk blending fertilizer in CR and 100% in GR, 100% bio-organic fertilizer in OR (Jul. 15). |
| Jointing | 20 | 23 Jul.–12 Aug | |
| Booting | 18 | 13 Aug.–31 Aug. | Applied 100% urea in CR and GR (Aug. 15). |
| Heading | 17 | 1 Sep.–18 Sep. | |
| Filling | 20 | 19 Sep.–9 Oct. | |
| Maturing | 19 | 10 Oct.–29 Oct. | Sowed Chinese milk vetch in GR and OR (Oct. 20) |
| Harvesting | 16 | 30 Oct.–14 Nov. | Harvested on Nov. 14. |
| Target Gene | Primer Set | Primer Sequence (5′-3′) | Reference |
|---|---|---|---|
| AOA a (amoA) | Arch-amoAF Arch-amoAR | STAATGGTCTGGCTTAGACG GCGGCCATCCATCTGTATGT | [37] |
| AOB b (amoA) | amoA1F amoA2R | GGGGTTTCTACTGGTGGT CCCCTCKGSAAAGCCTTCTTC | [38] |
| nifH | nifH-F nifH-R | AAAGGYGGWATCGGYAARTCCACCAC TTGTTSGCSGCRTACATSGCCATCAT | [39] |
| Cd-nitrite reductase (nirS) | Cd3aF R3cd | GTSAACGTSAAGGARACSGG GASTTCGGRTGSGTCTTGA | [40] |
| Cu-nitrite reductase (nirK) | FlaCu R3Cu | ATCATGGTSCTGCCGCG TTGGTGTTRGACTAGCTCCG | [40] |
| Nitrous oxide reductase (nosZ) | nosZ2F nosZ2R | CGCRACGGCAASAAGGTSMSSG CAKRTGCAKSGCRTGGCAGAA | [41] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yi, X.; Yuan, J.; Zhu, Y.; Yi, X.; Zhao, Q.; Fang, K.; Cao, L. Comparison of the Abundance and Community Structure of N-Cycling Bacteria in Paddy Rhizosphere Soil under Different Rice Cultivation Patterns. Int. J. Mol. Sci. 2018, 19, 3772. https://doi.org/10.3390/ijms19123772
Yi X, Yuan J, Zhu Y, Yi X, Zhao Q, Fang K, Cao L. Comparison of the Abundance and Community Structure of N-Cycling Bacteria in Paddy Rhizosphere Soil under Different Rice Cultivation Patterns. International Journal of Molecular Sciences. 2018; 19(12):3772. https://doi.org/10.3390/ijms19123772
Chicago/Turabian StyleYi, Xiaomei, Jing Yuan, Yuanhong Zhu, Xiaojian Yi, Qi Zhao, Kaikai Fang, and Linkui Cao. 2018. "Comparison of the Abundance and Community Structure of N-Cycling Bacteria in Paddy Rhizosphere Soil under Different Rice Cultivation Patterns" International Journal of Molecular Sciences 19, no. 12: 3772. https://doi.org/10.3390/ijms19123772