Transfection of T-Box Transcription Factor BRACHYURY and SOX2 Synergistically Promote Self-Renewal and Invasive Phenotype in Oral Cancer Cells


Abstract
:1. Introduction
2. Results
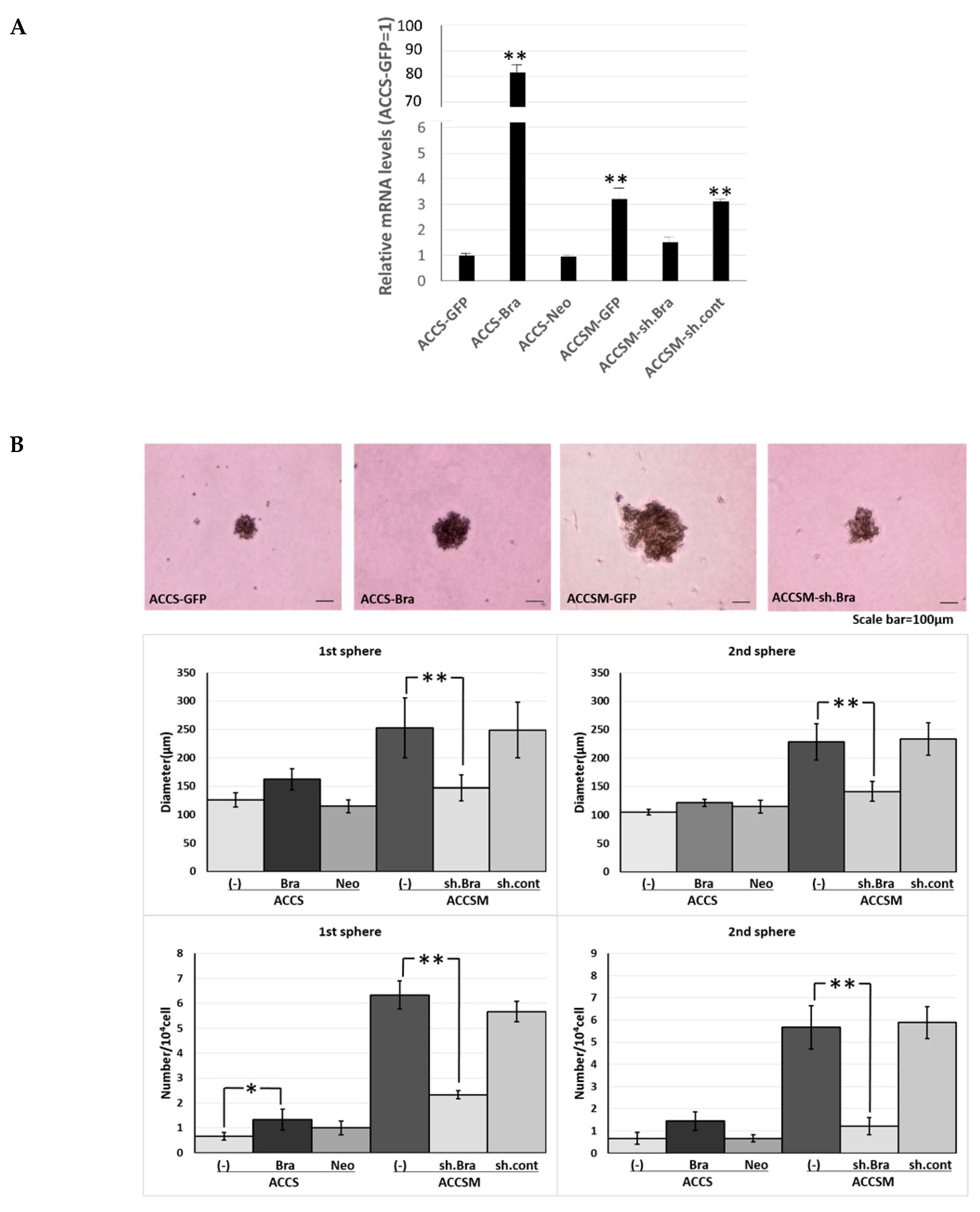
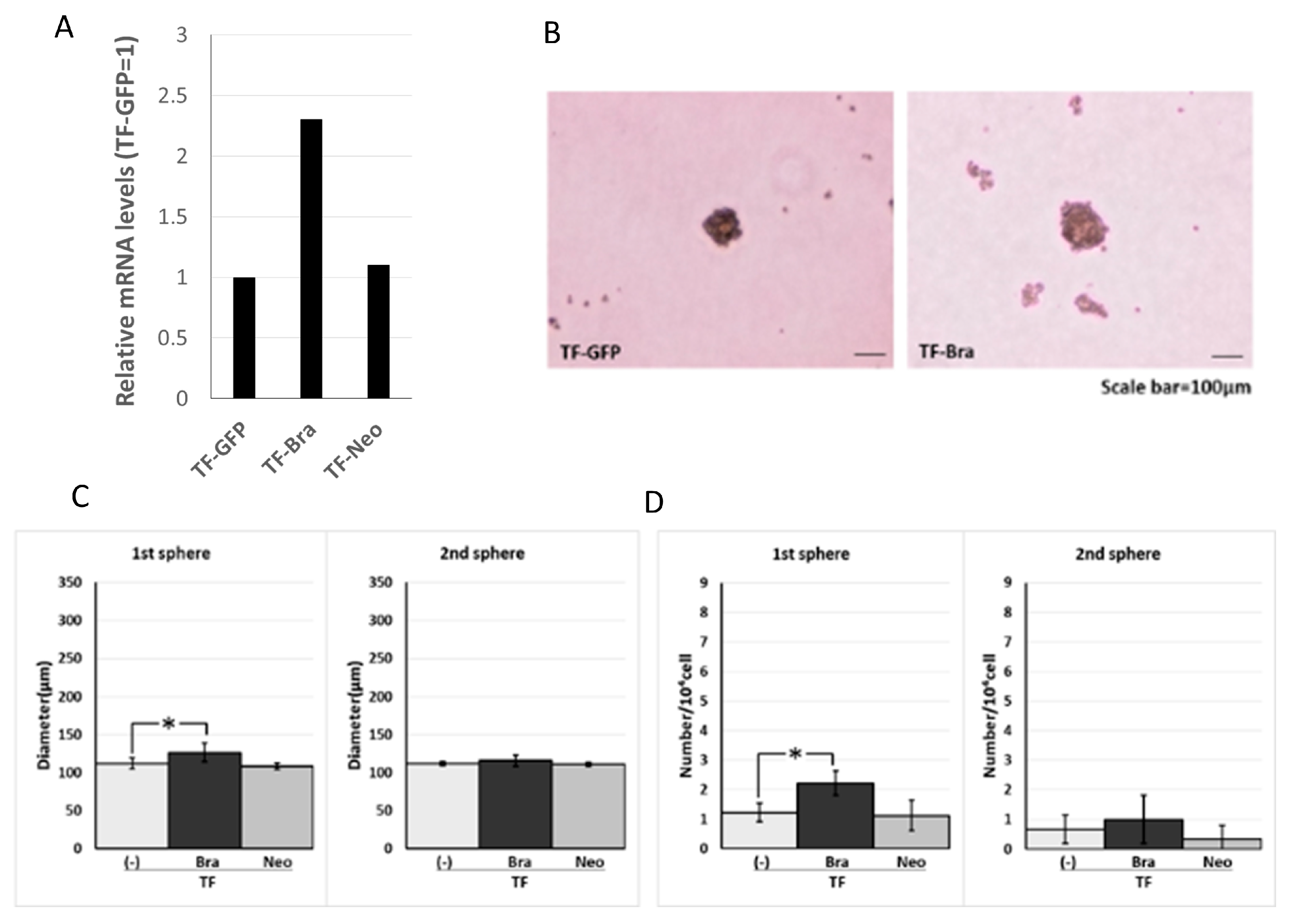
2.1. Forced Expression of BRACHYURY Does Not Promote Self-Renewal Capacity, But a BRACHYURYy Knockdown Suppresses the Self-Renewal Capacity in Oral Cancer Cell Lines
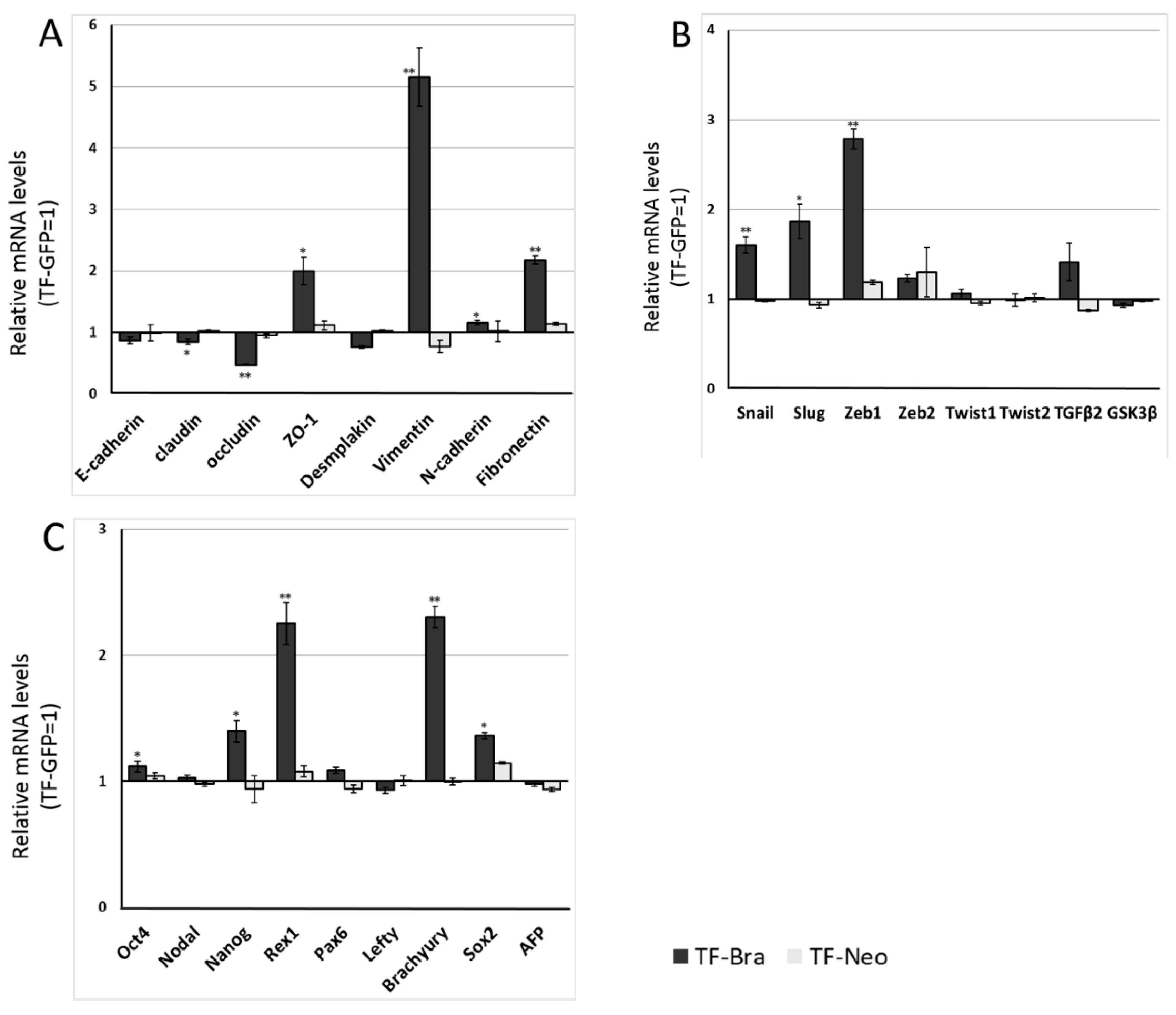
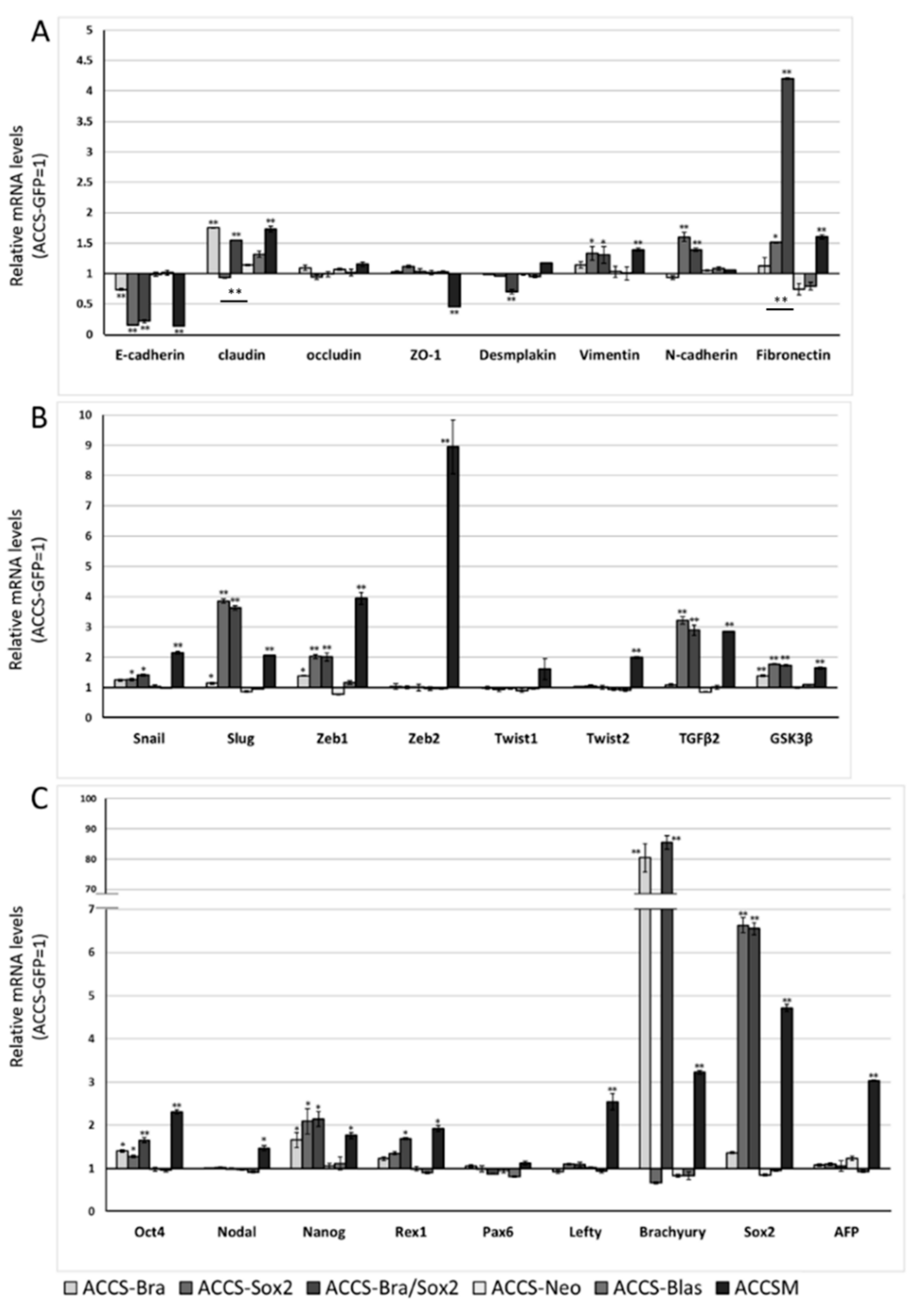
2.2. Forced Expression of BRACHYURY Weakly Induces EMT-Related Markers and Stem Cell Markers in Oral Cancer Cell Lines
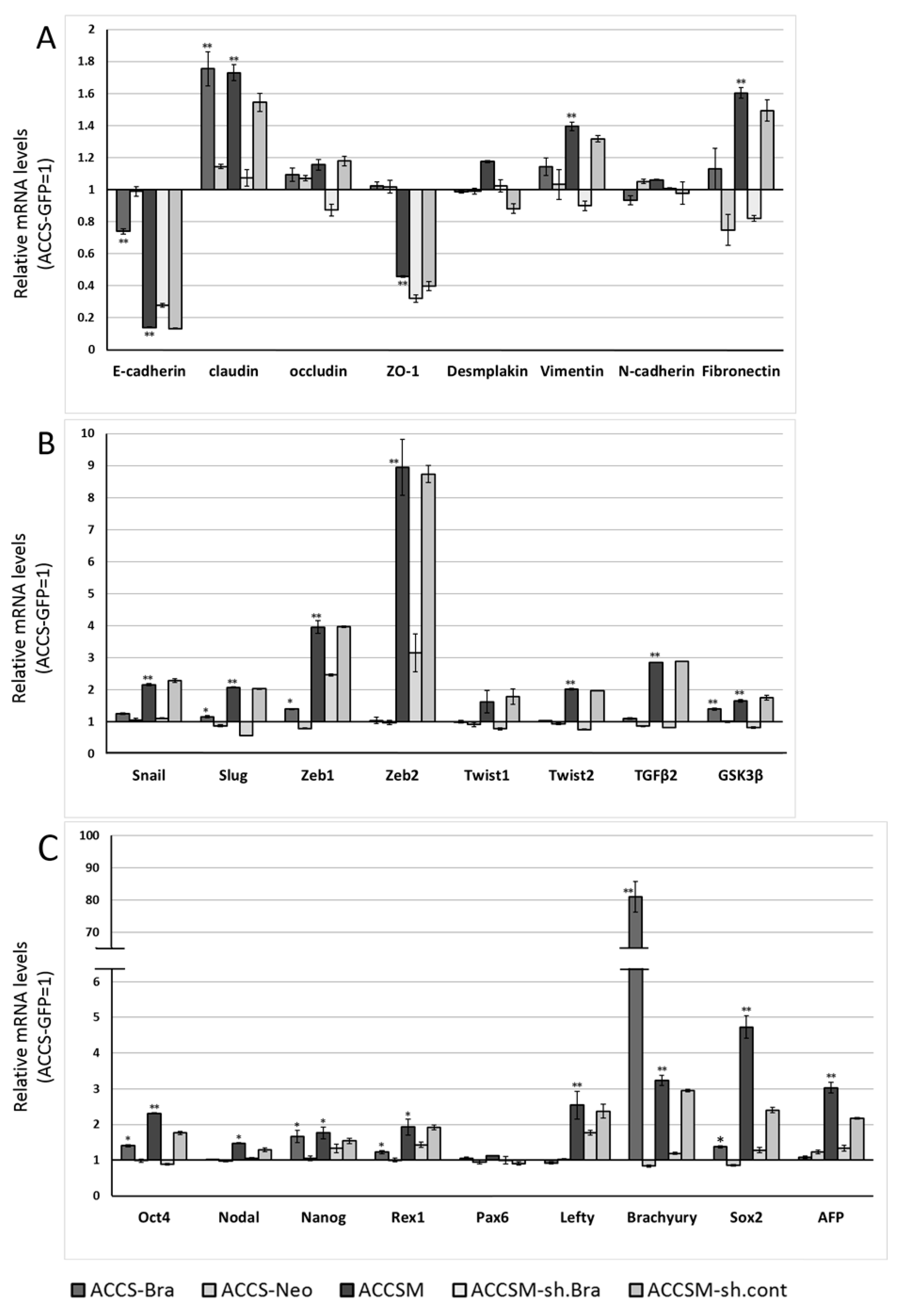
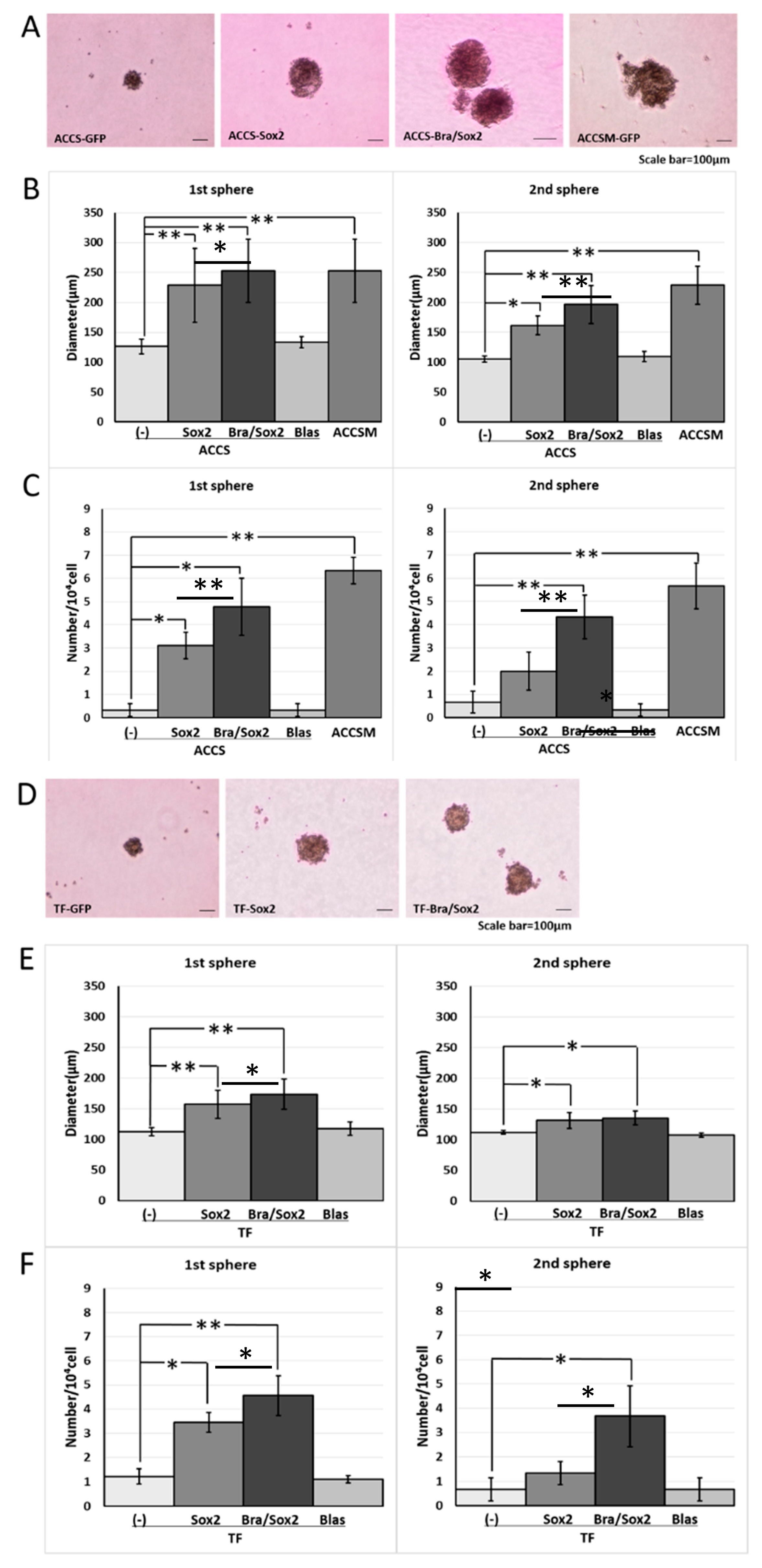
2.3. Forced Co-Expression of BRACHYURY and SOX2 Induced the Self-Renewal Phenotype in Oral Cancer Cell Lines
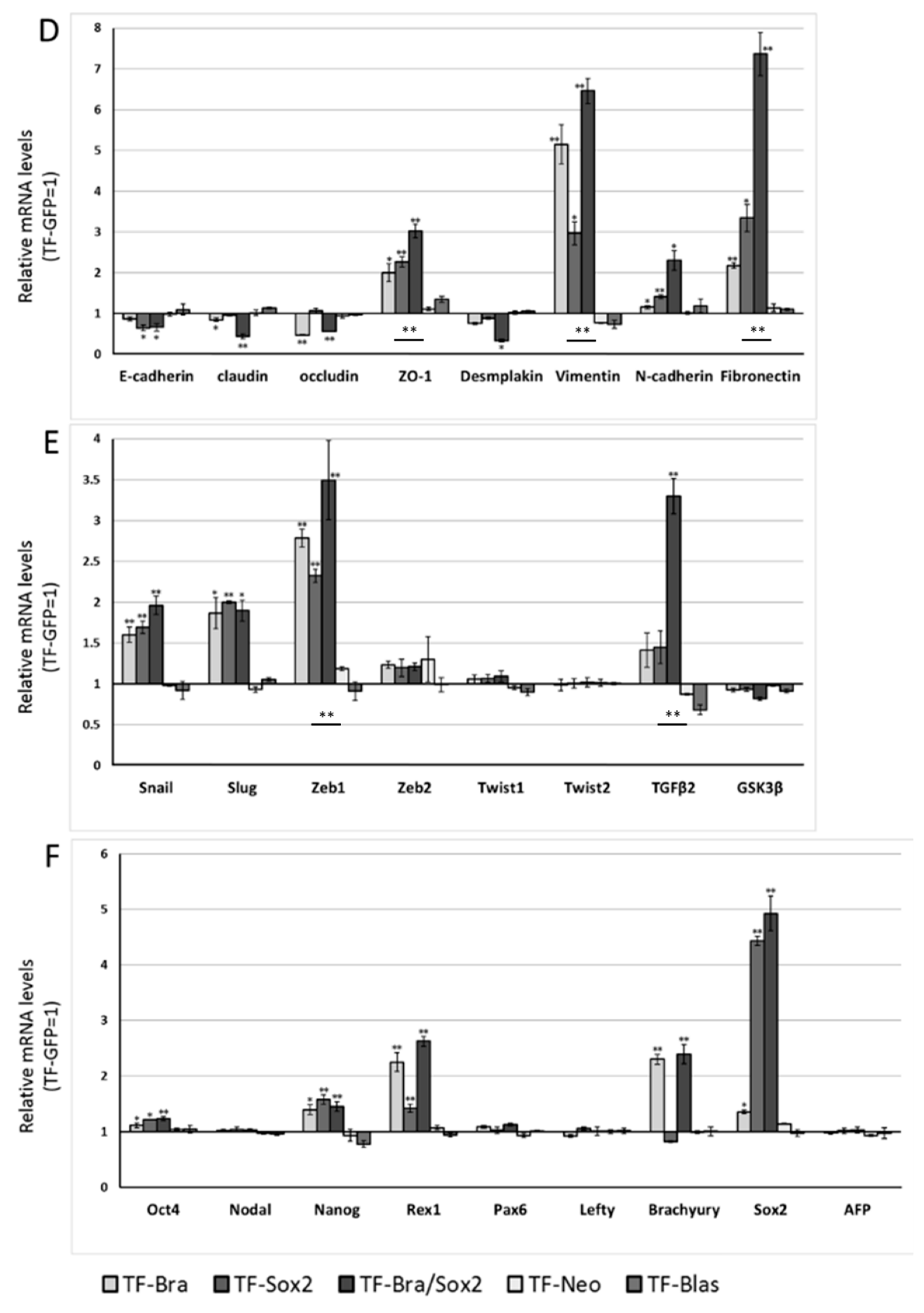
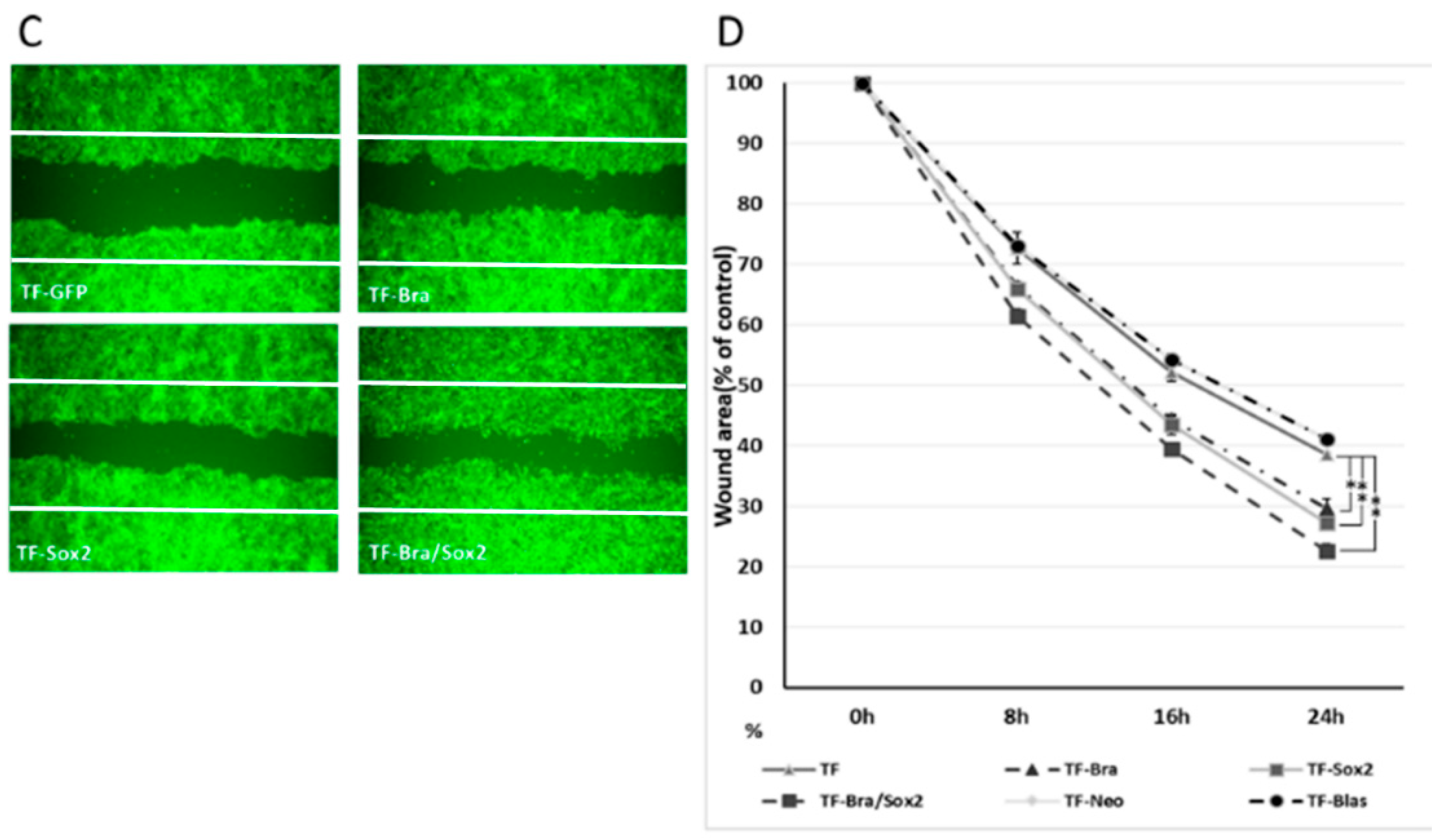
2.4. Forced Co-Expression of BRACHYURY and SOX2 Induced EMT-Related Markers and Stem Cell Markers in Oral Cancer Cell Lines
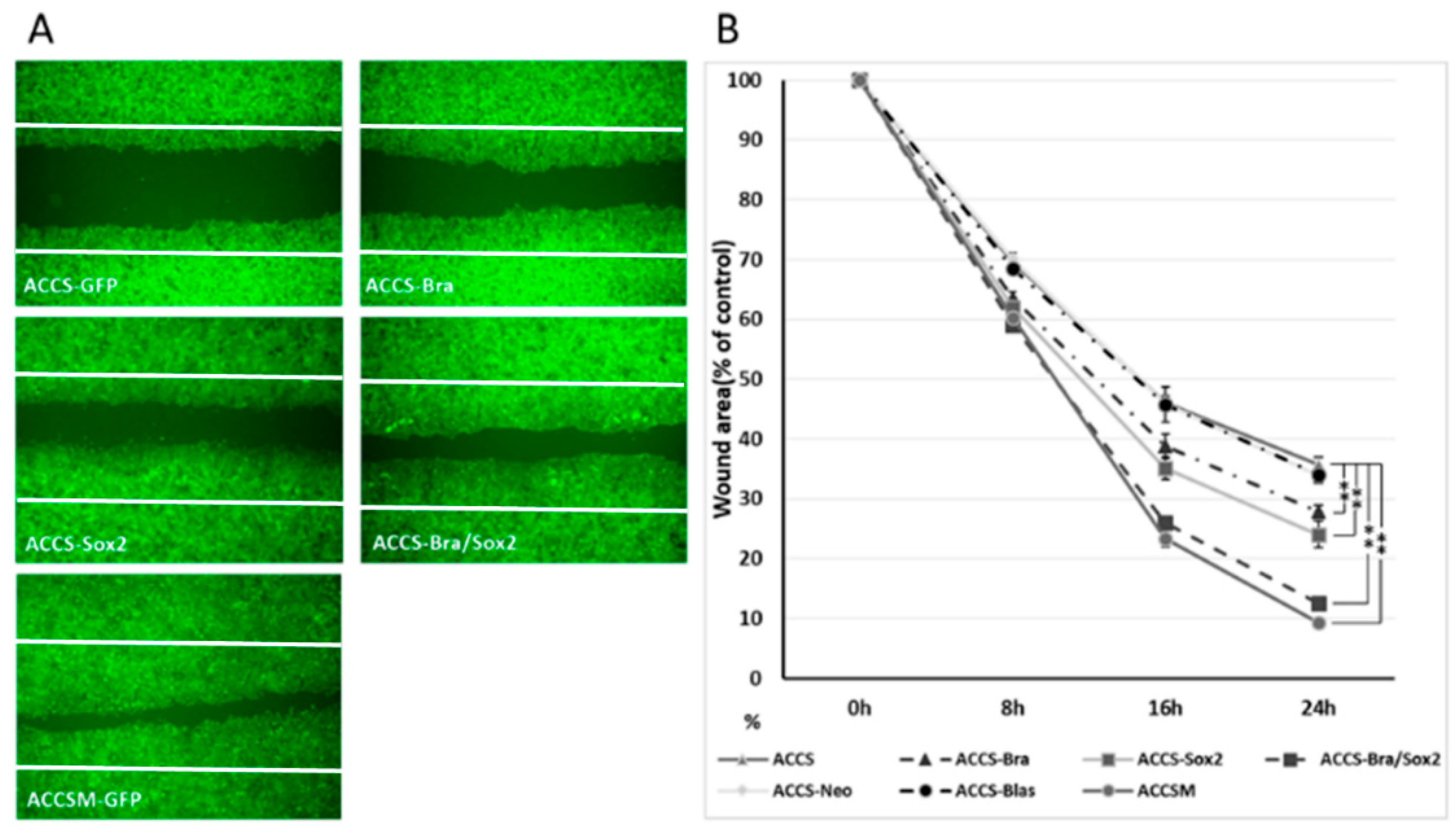
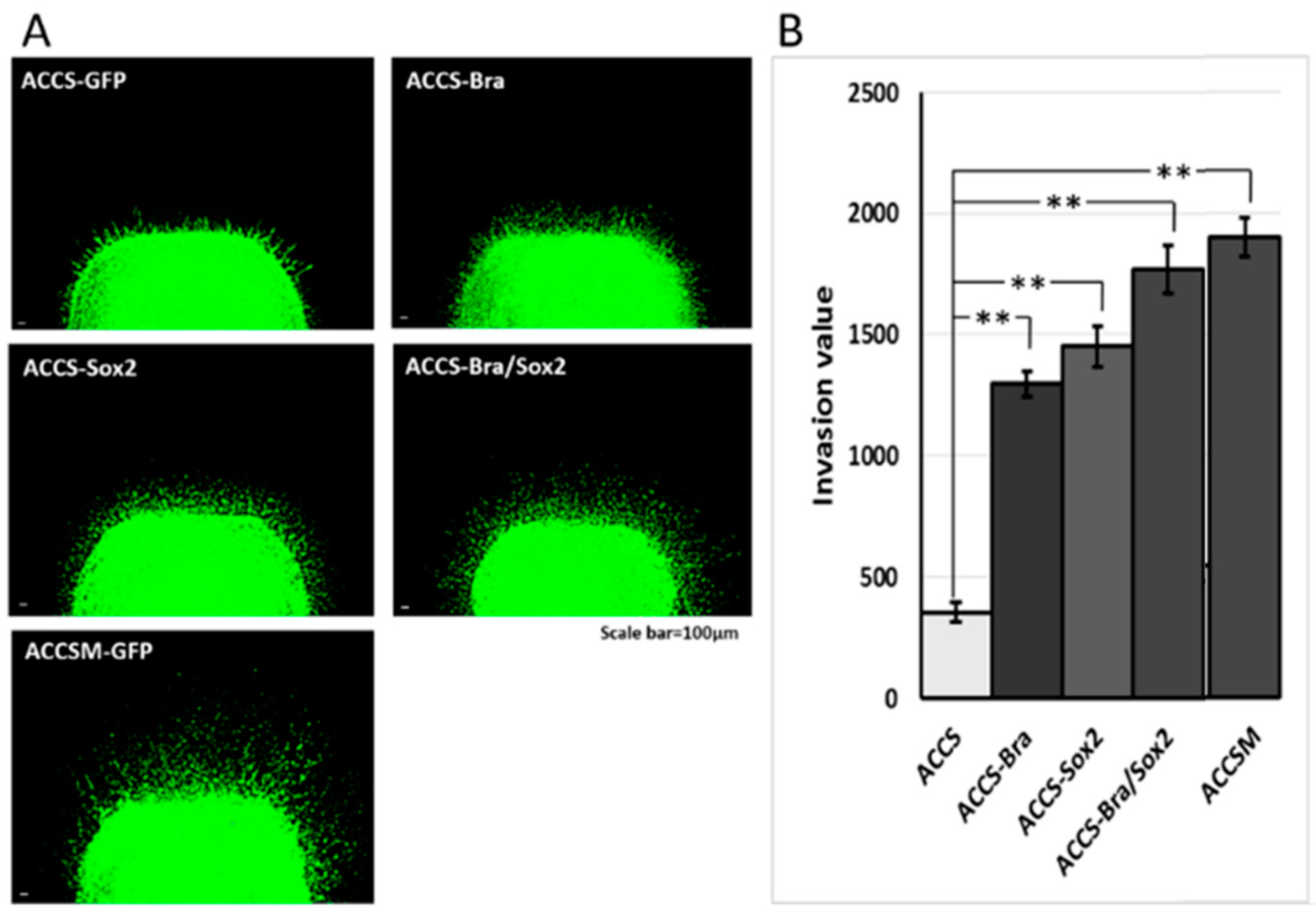
2.5. Artificial CSC-Like Cells Express an Invasive Phenotype In Vitro
3. Discussion
4. Materials and Methods
4.1. Cells and Culture
4.2. Transfection and Knockdown of BRACHYURY and SOX2
4.3. Real-Time RT-PCR
4.4. The Sphere-Formation Assay
4.5. The Wound Healing Assay
4.6. Evaluation of Tumour Dissemination from the Primary Cancer Nest
4.7. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| EMT | Epithelial–mesenchymal transition |
| CSCs | Cancer stem cells |
| shRNA | Small hairpin RNA |
| GFP | Green fluorescence protein |
| EDTA | Ethylendiaminetetraacetic acid |
References
- Chang, C.C.; Hsu, W.H.; Wang, C.C.; Chou, C.H.; Kuo, M.Y.; Lin, B.R.; Chen, S.T.; Tai, S.K.; Kuo, M.L.; Yang, M.H. Connective tissue growth factor activates pluripotency genes and mesenchymal-epithelial transition in head and neck cancer cells. Cancer Res. 2013, 73, 4147–4157. [Google Scholar] [CrossRef] [PubMed]
- Drasin, D.J.; Robin, T.P.; Ford, H.L. Breast cancer epithelial-to-mesenchymal transition: Examining the functional consequences of plasticity. Breast Cancer Res. 2011, 13, 226. [Google Scholar] [CrossRef] [PubMed]
- Pirozzi, G.; Tirino, V.; Camerlingo, R.; La Rocca, A.; Martucci, N.; Scognamiglio, G.; Franco, R.; Cantile, M.; Normanno, N.; Rocco, G. Prognostic value of cancer stem cells, epithelial-mesenchymal transition and circulating tumor cells in lung cancer. Oncol. Rep. 2013, 29, 1763–1768. [Google Scholar] [CrossRef] [PubMed]
- Zubeldia, I.G.; Bleau, A.M.; Redrado, M.; Serrano, D.; Agliano, A.; Gil-Puig, C.; Vidal-Vanaclocha, F.; Lecanda, J.; Calvo, A. Epithelial to mesenchymal transition and cancer stem cell phenotypes leading to liver metastasis are abrogated by the novel TGFbeta1-targeting peptides P17 and P144. Exp. Cell Res. 2013, 319, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Imajyo, I.; Sugiura, T.; Kobayashi, Y.; Shimoda, M.; Ishii, K.; Akimoto, N.; Yoshihama, N.; Kobayashi, I.; Mori, Y. T-box transcription factor Brachyury expression is correlated with epithelial-mesenchymal transition and lymph node metastasis in oral squamous cell carcinoma. Int. J. Oncol. 2012, 41, 1985–1995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mani, S.A.; Guo, W.; Liao, M.J.; Eaton, E.N.; Ayyanan, A.; Zhou, A.Y.; Brooks, M.; Reinhard, F.; Zhang, C.C.; Shipitsin, M.; et al. The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell 2008, 133, 704–715. [Google Scholar] [CrossRef] [PubMed]
- Hindriksen, S.; Bijlsma, M.F. Cancer stem cells, EMT, and developmental pathway activation in pancreatic tumors. Cancers 2012, 4, 989–1035. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.S.; Han, Z.P.; Jing, Y.Y.; Tao, S.F.; Li, T.J.; Wang, H.; Wang, Y.; Li, R.; Yang, Y.; Zhao, X.; et al. CD133(+)CXCR4(+) colon cancer cells exhibit metastatic potential and predict poor prognosis of patients. BMC Med. 2012, 10, 85. [Google Scholar] [CrossRef] [PubMed]
- Mallini, P.; Lennard, T.; Kirby, J.; Meeson, A. Epithelial-to-mesenchymal transition: What is the impact on breast cancer stem cells and drug resistance. Cancer Treat. Rev. 2014, 40, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.L.; Pernemalm, M.; Crosbie, P.A.; Whetton, A.D. The role of the tumor-microenvironment in lung cancer-metastasis and its relationship to potential therapeutic targets. Cancer Treat. Rev. 2014, 40, 558–566. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, M.; Sugiura, T.; Imajyo, I.; Ishii, K.; Chigita, S.; Seki, K.; Kobayashi, Y.; Shirasuna, K. The T-box transcription factor Brachyury regulates epithelial-mesenchymal transition in association with cancer stem-like cells in adenoid cystic carcinoma cells. BMC Cancer 2012, 12, 377. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.B.; Chaffer, C.L.; Weinberg, R.A. Cancer stem cells: Mirage or reality? Nat. Med. 2009, 15, 1010–1012. [Google Scholar] [CrossRef] [PubMed]
- Brabletz, S.; Brabletz, T. The ZEB/miR-200 feedback loop—A motor of cellular plasticity in development and cancer? EMBO Rep. 2010, 11, 670–677. [Google Scholar] [CrossRef] [PubMed]
- Fuxe, J.; Vincent, T.; Garcia de Herreros, A. Transcriptional crosstalk between TGFβ and stem cell pathways in tumor cell invasion: Role of EMT promoting Smad complexes. Cell Cycle 2010, 9, 2363–2374. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Kang, Y. Multilayer control of the EMT master regulators. Oncogene 2014, 33, 1755. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, Y.; Ahmad, A.; Banerjee, S.; Azmi, A.S.; Kong, D.; Sarkar, F.H. Pancreatic cancer: Understanding and overcoming chemoresistance. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Settleman, J. EMT, cancer stem cells and drug resistance: An emerging axis of evil in the war on cancer. Oncogene 2010, 29, 4741–4751. [Google Scholar] [CrossRef] [PubMed]
- Shekhani, M.T.; Jayanthy, A.S.; Maddodi, N.; Setaluri, V. Cancer stem cells and tumor transdifferentiation: Implications for novel therapeutic strategies. Am. J. Stem Cells 2013, 2, 52–61. [Google Scholar] [PubMed]
- Smalley, M.; Piggott, L.; Clarkson, R. Breast cancer stem cells: Obstacles to therapy. Cancer Lett. 2013, 338, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Palena, C.; Polev, D.E.; Tsang, K.Y.; Fernando, R.I.; Litzinger, M.; Krukovskaya, L.L.; Baranova, A.V.; Kozlov, A.P.; Schlom, J. The human T-box mesodermal transcription factor Brachyury is a candidate target for T-cell-mediated cancer immunotherapy. Clin. Cancer Res. 2007, 13, 2471–2478. [Google Scholar] [CrossRef] [PubMed]
- Fernando, R.I.; Litzinger, M.; Trono, P.; Hamilton, D.H.; Schlom, J.; Palena, C. The T-box transcription factor Brachyury promotes epithelial-mesenchymal transition in human tumor cells. J. Clin. Investig. 2010, 120, 533–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishii, K.; Shimoda, M.; Sugiura, T.; Seki, K.; Takahashi, M.; Abe, M.; Matsuki, R.; Inoue, Y.; Shirasuna, K. Involvement of epithelial-mesenchymal transition in adenoid cystic carcinoma metastasis. Int. J. Oncol. 2011, 38, 921–931. [Google Scholar] [CrossRef] [PubMed]
- Abe, M.; Sugiura, T.; Takahashi, M.; Ishii, K.; Shimoda, M.; Shirasuna, K. A novel function of CD82/KAI-1 on E-cadherin-mediated homophilic cellular adhesion of cancer cells. Cancer Lett. 2008, 266, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Biddle, A.; Mackenzie, I.C. Cancer stem cells and EMT in carcinoma. Cancer Metast. Rev. 2012, 31, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Rhim, A.D.; Mirek, E.T.; Aiello, N.M.; Maitra, A.; Bailey, J.M.; McAllister, F.; Reichert, M.; Beatty, G.L.; Rustgi, A.K.; Vonderheide, R.H.; et al. EMT and dissemination precede pancreatic tumor formation. Cell 2012, 148, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Wellner, U.; Schubert, J.; Burk, U.C.; Schmalhofer, O.; Zhu, F.; Sonntag, A.; Waldvogel, B.; Vannier, C.; Darling, D.; zur Hausen, A.; et al. The EMT-activator ZEB1 promotes tumorigenicity by repressing stemness-inhibiting microRNAs. Nat. Cell Biol. 2009, 11, 1487–1495. [Google Scholar] [CrossRef] [PubMed]
- Nicolini, A.; Ferrari, P.; Fini, M.; Borsari, V.; Fallahi, P.; Antonelli, A.; Berti, P.; Carpi, A.; Miccoli, P. Stem cells: Their role in breast cancer development and resistance to treatment. Curr. Pharm. Biotechnol. 2011, 12, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Pestell, T.G.; Lisanti, M.P.; Pestell, R.G. Cancer stem cells. Int. J. Biochem. Cell Biol. 2012, 44, 2144–2151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, C.D.; Parvani, J.G.; Schiemann, W.P. The relevance of the TGF-beta Paradox to EMT-MET programs. Cancer Lett. 2013, 341, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Brabletz, T. EMT and MET in metastasis: Where are the cancer stem cells? Cancer Cell 2012, 22, 699–701. [Google Scholar] [CrossRef] [PubMed]
- Kong, D.; Banerjee, S.; Ahmad, A.; Li, Y.; Wang, Z.; Sethi, S.; Sarkar, F.H. Epithelial to mesenchymal transition is mechanistically linked with stem cell signatures in prostate cancer cells. PLoS ONE 2010, 5, e12445. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, D.; Shields, B.; Davies, M.L.; Muller, J.; Wakeman, J.A. BRACHYURY confers cancer stem cell characteristics on colorectal cancer cells. Int. J. Cancer 2012, 130, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Chambers, I.; Silva, J.; Colby, D.; Nichols, J.; Nijmeijer, B.; Robertson, M.; Vrana, J.; Jones, K.; Grotewold, L.; Smith, A. Nanog safeguards pluripotency and mediates germline development. Nature 2007, 450, 1230–1234. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R. EMT: When epithelial cells decide to become mesenchymal-like cells. J. Clin. Investig. 2009, 119, 1417–1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeisberg, M.; Neilson, E.G. Biomarkers for epithelial-mesenchymal transitions. J. Clin. Investig. 2009, 119, 1429–1437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heldin, C.H.; Vanlandewijck, M.; Moustakas, A. Regulation of EMT by TGFβ in cancer. FEBS Lett. 2012, 586, 1959–1970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herreros-Villanueva, M.; Zhang, J.S.; Koenig, A.; Abel, E.V.; Smyrk, T.C.; Bamlet, W.R.; de Narvajas, A.A.; Gomez, T.S.; Simeone, D.M.; Bujanda, L.; et al. SOX2 promotes dedifferentiation and imparts stem cell-like features to pancreatic cancer cells. Oncogenesis 2013, 2, e61. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Fang, X.; Lou, X.; Hua, D.; Ding, W.; Foltz, G.; Hood, L.; Yuan, Y.; Lin, B. Silencing SOX2 induced mesenchymal-epithelial transition and its expression predicts liver and lymph node metastasis of CRC patients. PLoS ONE 2012, 7, e41335. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xu, Y.; Chen, Y.; Chen, S.; Jia, X.; Sun, T.; Liu, Y.; Li, X.; Xiang, R.; Li, N. SOX2 promotes tumor metastasis by stimulating epithelial-to-mesenchymal transition via regulation of WNT/β-catenin signal network. Cancer Lett. 2013, 336, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.; Zhu, X. The Wnt/β-catenin pathway regulates self-renewal of cancer stem-like cells in human gastric cancer. Mol. Med. Rep. 2012, 5, 1191–1196. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Rao, S.; Chu, J.; Shen, X.; Levasseur, D.N.; Theunissen, T.W.; Orkin, S.H. A protein interaction network for pluripotency of embryonic stem cells. Nature 2006, 444, 364–368. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Xu, Y.; Chen, Y.; Li, X.; Mou, W.; Wang, L.; Liu, Y.; Reisfeld, R.A.; Xiang, R.; Lv, D.; et al. SOX2 gene regulates the transcriptional network of oncogenes and affects tumorigenesis of human lung cancer cells. PLoS ONE 2012, 7, e36326. [Google Scholar] [CrossRef] [PubMed]
- Arnold, S.J.; Stappert, J.; Bauer, A.; Kispert, A.; Herrmann, B.G.; Kemler, R. Brachyury is a target gene of the Wnt/β-catenin signaling pathway. Mech. Dev. 2000, 91, 249–258. [Google Scholar] [CrossRef]
- Larocca, C.; Cohen, J.R.; Fernando, R.I.; Huang, B.; Hamilton, D.H.; Palena, C. An autocrine loop between TGF-β1 and the transcription factor brachyury controls the transition of human carcinoma cells into a mesenchymal phenotype. Mol. Cancer Ther. 2013, 12, 1805–1815. [Google Scholar] [CrossRef] [PubMed]
- Hotta, K.; Takahashi, H.; Satoh, N.; Gojobori, T. Brachyury-downstream gene sets in a chordate, Ciona intestinalis: Integrating notochord specification, morphogenesis and chordate evolution. Evol. Dev. 2008, 10, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Ikushima, H.; Todo, T.; Ino, Y.; Takahashi, M.; Miyazawa, K.; Miyazono, K. Autocrine TGF-β signaling maintains tumorigenicity of glioma-initiating cells through Sry-related HMG-box factors. Cell Stem Cell 2009, 5, 504–514. [Google Scholar] [CrossRef] [PubMed]
- Lou, X.; Han, X.; Jin, C.; Tian, W.; Yu, W.; Ding, D.; Cheng, L.; Huang, B.; Jiang, H.; Lin, B. SOX2 targets fibronectin 1 to promote cell migration and invasion in ovarian cancer: New molecular leads for therapeutic intervention. Omics J. Integr. Biol. 2013, 17, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Hunt, G.C.; Singh, P.; Schwarzbauer, J.E. Endogenous production of fibronectin is required for self-renewal of cultured mouse embryonic stem cells. Exp. Cell Res. 2012, 318, 1820–1831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gavert, N.; Vivanti, A.; Hazin, J.; Brabletz, T.; Ben-Ze’ev, A. L1-mediated colon cancer cell metastasis does not require changes in EMT and cancer stem cell markers. Mol. Cancer Res. 2011, 9, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Lou, Y.; Preobrazhenska, O.; auf dem Keller, U.; Sutcliffe, M.; Barclay, L.; McDonald, P.C.; Roskelley, C.; Overall, C.M.; Dedhar, S. Epithelial-mesenchymal transition (EMT) is not sufficient for spontaneous murine breast cancer metastasis. Dev. Dyn. 2008, 237, 2755–2768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chikaishi, Y.; Uramoto, H.; Tanaka, F. The EMT status in the primary tumor does not predict postoperative recurrence or disease-free survival in lung adenocarcinoma. Anticancer Res. 2011, 31, 4451–4456. [Google Scholar] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene (Human) | Primer Sequence |
|---|---|
| E-CADHERIN | (F) 5′ CAA CTG GAC CAT TCA GTA CAA C 3′ (R) 5′ TCC ATG AGC TTG AGA TTG AT 3′ |
| CLAUDIN | (F) 5′ GAC AAC ATT CAC TGCC TCA GG 3′ (R) 5′ TTC ACA TTT GGT GAT TCT CG 3′ |
| OCCULUDIN | (F) 5′ CTC GAA GAA AGA TGG ACA GGT 3′ (R) 5′ GCC ATG GGA CTG TCA ACT C 3′ |
| ZO-1 | (F) 5′ CGA AGG AGT TGA GCA GGA AAT CT 3′ (R) 5′ TCC ACA GGC TTC AGG AAC TTG 3′ |
| DESMOPLAKIN | (F) 5′ ACC GCT GGC AAA GGA TAG AT 3′ (R) 5′ CCA CTT GCA GAA AGC CTG AT 3′ |
| VIMENTIN | (F) 5′ ATT CAC TCC CTC TGG TTG ATA C 3′ (R) 5′ CGT GAT GCT GAG AAG TTT CG 3′ |
| N-CADHERIN | (F) 5′ GAC AAC ATT CAC TGC TCA GG 3′ (R) 5′ TTC ACA TTT GGT GAT TCT CG 3′ |
| FIBRONECTIN 1 | (F) 5′ CAA TGC CAG GAT TCA GAG AC 3′ (R) 5′ CTT CGA CAG GAC CAC TTG AG 3′ |
| SNAIL | (F) 5′ TCC ACA AGC ACCAAG AGT C 3′ (R) 5′ ATG GCA GTG AGA AGG ATG TG 3′ |
| SLUG | (F) 5′ ACT GCT CCA AAA CCT TCT CC 3′ (R) 5′ TGG TCA GCA CAG GAG AAA ATG 3′ |
| TWIST 1 | (F) 5′ CTC AGC TAC GCC TTC TCG 3′ (R) 5′ ACT GTC CAT TTT CTC ATT CTC TG 3′ |
| TWIST 2 | (F) 5′ AGG AGC TCG AGA GGC AG 3′ (R) 5′ CGT TGA GCG ACT GGC TG 3′ |
| ZEB1 | (F) 5′ CTC ACA CTC TGG GTC TTA TTC TC 3′ (R) 5′ GTC TTC ATC CTC TTC CCT TGT C 3′ |
| ZEB2 | (F) 5′ AAA GGA GAA AGT ACC AGC GG 3′ (R) 5′ AGG AGT CGG AGT CTG TCA TAT C 3′ |
| TGFB2 | (F) 5′ TTA ACA TCT CCA ACC CAG CG 3′ (R) 5′ TCC TGT CTTTAT GGT GAAGCC 3′ |
| GSK3B | (F) 5′ GGT CTA TCT TAA TCT GGT GCT GG 3′ (R) 5′ AGG TTC TGC GGT TTA ATA TCC C 3′ |
| NODAL | (F) 5′ ACC CAG CTG TGT GTA CTC AA 3′ (R) 5′ TGG TAA CGT TTC AGC AGA C 3′ |
| OCT4 | (F) 5′ TAT CGA GAA CCG AGT GAG AG 3′ (R) 5′ TCG TTG TGC ATA GTC GCT 3′ |
| PAX6 | (F) 5′ GGC GGA GTT ATG TAT ACC TAC 3′ (R) 5′ CTT GGC CAG TAT TGA GAC AT 3′ |
| REX1 | (F) 5′ AAA CGG GCA AAG ACA AGA 3′ (R) 5′ GCT CAT AGC ACA CAT AGC CAT 3′ |
| LEFTY | (F) 5′ TGT ATC CAT TGA GCC CTC T 3′ (R) 5′ CAG GAA ATG GAA GGA CAC A 3′ |
| NANOG | (F) 5′ ACC CAG CTG TGT GTA CTC AA 3′ (R) 5′ GCG TCA CCA TTG CTA TT 3′ |
| BRACHYURY | (F) 5′ TGC TGC AAT CCC ATG ACA 3′ (R) 5′ CGT TGC TCA CAG ACC ACA 3′ |
| SOX2 | (F) 5′ TGG GTT CGG TGG TCA AGT 3′ (R) 5′ CTC TGG TAG TGCTGG GAC A3′ |
| AFP | (F) 5′ CTG CAA ACT GAC CAC GCT 3′ (R) 5′ TGA GAC AGC AAG CTG AGG AT 3′ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akimoto, N.; Nakamura, K.; Hijioka, H.; Kume, K.; Matsumura, Y.; Sugiura, T. Transfection of T-Box Transcription Factor BRACHYURY and SOX2 Synergistically Promote Self-Renewal and Invasive Phenotype in Oral Cancer Cells. Int. J. Mol. Sci. 2018, 19, 3620. https://doi.org/10.3390/ijms19113620
Akimoto N, Nakamura K, Hijioka H, Kume K, Matsumura Y, Sugiura T. Transfection of T-Box Transcription Factor BRACHYURY and SOX2 Synergistically Promote Self-Renewal and Invasive Phenotype in Oral Cancer Cells. International Journal of Molecular Sciences. 2018; 19(11):3620. https://doi.org/10.3390/ijms19113620
Chicago/Turabian StyleAkimoto, Naonari, Kodai Nakamura, Hiroshi Hijioka, Kenichi Kume, Yoshiaki Matsumura, and Tsuyoshi Sugiura. 2018. "Transfection of T-Box Transcription Factor BRACHYURY and SOX2 Synergistically Promote Self-Renewal and Invasive Phenotype in Oral Cancer Cells" International Journal of Molecular Sciences 19, no. 11: 3620. https://doi.org/10.3390/ijms19113620