Hepatocyte Growth Factor Activator: A Proteinase Linking Tissue Injury with Repair


1
Section of Oncopathology and Regenerative Biology, Faculty of Medicine, University of Miyazaki, 5200 Kihara, Kiyotake 889-1692, Miyazaki, Japan
2
Department of Surgery, Miyakonojyo Medical Association Hospital, 1364-1 Tarobocho, Miyakonojyo 885-0002, Miyazaki, Japan
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2018, 19(11), 3435; https://doi.org/10.3390/ijms19113435
Submission received: 27 September 2018
/
Revised: 22 October 2018
/
Accepted: 30 October 2018
/
Published: 1 November 2018
(This article belongs to the Special Issue Hepatocyte Growth Factor (HGF))
Abstract
:Hepatocyte growth factor (HGF) promotes pleiotropic signaling through its specific receptor tyrosine kinase, MET. As such, it has important roles in the regeneration of injured tissues. Since HGF is produced mainly by mesenchymal cells and MET is expressed in most epithelial, endothelial and somatic stem cells, HGF functions as a typical paracrine growth factor. HGF is secreted as an inactive precursor (proHGF) and requires proteolytic activation to initiate HGF-induced MET signaling. HGF activator (HGFAC) is a serum activator of proHGF and produces robust HGF activities in injured tissues. HGFAC is a coagulation factor XII-like serine endopeptidase that circulates in the plasma as a zymogen (proHGFAC). Thrombin, kallikrein-related peptidase (KLK)-4 or KLK-5 efficiently activates proHGFAC. The activated HGFAC cleaves proHGF at Arg494-Val495, resulting in the formation of the active disulfide-linked heterodimer HGF. Macrophage stimulating protein, a ligand of RON, is also activated by HGFAC in vivo. Although HGFAC functions primarily at the site of damaged tissue, a recent report has suggested that activated HGFAC relays a signal to stem cells in non-injured tissues via proHGF activation in the stem cell niche. This review focuses on current knowledge regarding HGFAC-mediated proHGF activation and its roles in tissue regeneration and repair.
1. Activation of Hepatocyte Growth Factor (HGF) Precursor
HGF is a pleiotropic peptide growth factor that was discovered as a potent mitogen for cultured hepatocytes [1]. This molecule is identical to scatter factor, an independently identified molecule that dissociated epithelial cells and increased their migratory activity [2]. Now, it is well established that HGF plays a major role in the regeneration and repair of the liver as well as other organs, including the gastrointestinal tract, kidney, lung and nervous system [3,4,5,6]. HGF is a typical paracrine factor that is produced by mesenchymal cells in a precursor form (proHGF) that acts on MET-expressing parenchymal cells after its proteolytic activation [7]. In this context, regulation of the activating proteinase is a very important step in the control of HGF-MET signaling [8]. Indeed, the HGF molecule detected in normal tissues is primarily in a form of single-chain inactive proHGF [9,10].
Several proteinases have been reported to activate proHGF. All are serine proteinases and they can be categorized into two groups: (1) circulating proteinases that emerge after activation of the coagulation cascade (serum proteinases) and, (2) tissue proteinases that function as proHGF activators in localized pericellular microenvironments (cellular proteinases) (Table 1) [11]. ProHGF protein has a structure similar to plasminogen. Consequently, plasminogen activators, particularly urokinase-type plasminogen activator (uPA), were initially reported to be proHGF converting enzymes [12]. However, the processing by uPA was a stoichiometric reaction [13], not an enzymatic reaction, and thus, its efficacy to produce mature HGF was very low [13,14]. It is now believed that the major proHGF-converting proteinases are serum HGF activator (HGFAC) and cellular type II transmembrane serine proteinases (TTSP), particularly matriptase [11,14,15,16].
2. Discovery, Molecular Structure and Physiological Substrates of HGFAC
In the process of purifying recombinant human HGF from culture supernatants of Chinese hamster ovary cells transfected with the expression vector harboring full-length HGF cDNA, Shimomura et al. [17] noticed that fetal bovine serum (FBS) in the growth medium had a potent activity, converting the single-chain inactive proHGF to mature two-chain active HGF. In 1992, they purified the corresponding proteinase from FBS and named it HGFAC [17]. Shortly afterwards, the human counterpart of this novel proteinase was purified from human serum and its cDNA was cloned [18]. The cloning study revealed that HGFAC is a coagulation factor XII-like serine endopeptidase of the trypsin-like S1 family. Moreover, it was found to be initially synthesized as a single-chain inactive zymogen (proHGFAC) [18]. Indeed, the phylogeny of the cDNA sequences of HGFAC and factor XII (F12) genes indicated that the gene for factor XII evolved from duplication of the HGFAC gene [19,20]. It is noteworthy that proHGF protein is homologous to plasminogen and has the same ancestral gene as plasminogen [19]. Therefore, the HGFAC-HGF system likely evolved along with that of the coagulation and fibrinolysis systems, suggesting its primary role is involved in the host’s response to tissue injuries. In accordance with this assumption, thrombin efficiently converts proHGFAC to the active two-chain form of HGFAC [21].
ProHGFAC is primarily synthesized by hepatocytes and circulates in plasma [8]. It is a relatively abundant plasma protein with a mean concentration around 40 nM in healthy individuals. On the other hand, low but distinct expression of HGFAC mRNA has been reported in extrahepatic organs, including the gastrointestinal tract, kidney, lung and central nervous system [8]. Regarding the substrates for HGFAC, only two proteins have been reported in vivo: proHGF and pro-macrophage stimulating protein (proMSP) [8,22]. MSP is an HGF-like protein synthesized by the liver and circulating in plasma (2–5 nM) as an inactive precursor form (proMSP) [22,23,24]. HGFAC cleaves proMSP at the Arg483-Val484 bond and generates a two-chain active MSP that exerts its biological activity through RON receptor tyrosine kinase, expressed by macrophages, epithelial cells and cancer cells [23]. However, considering its relatively high concentration in the plasma, it would not be surprising if HGFAC had additional unknown physiological substrates in vivo. Further studies for the substrates and biological functions of HGFAC in vivo will be required.
3. Robust Activation of proHGF by HGFAC in Response to Tissue Injury and Inflammation
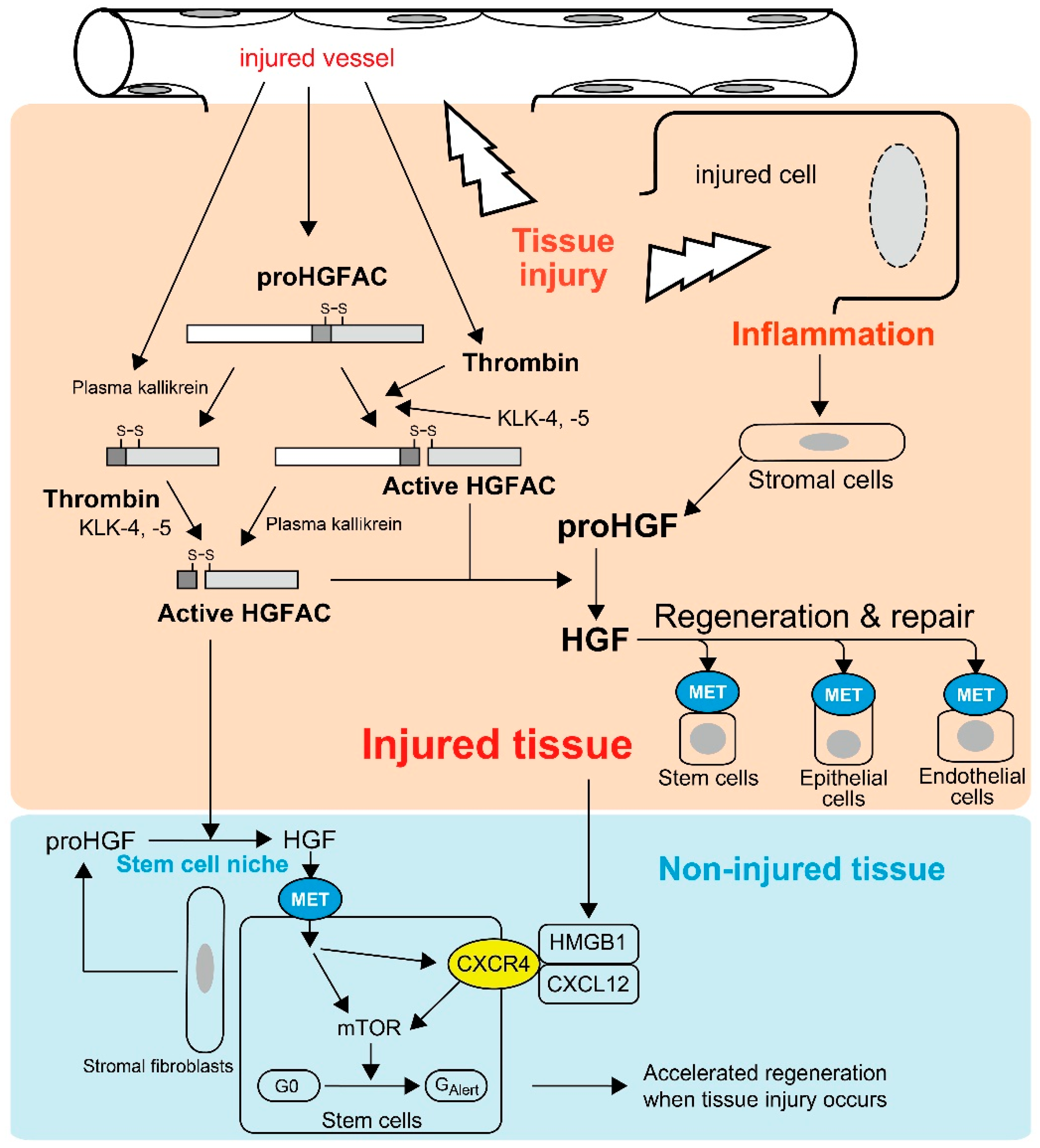
Most HGFAC proteins circulate in plasma as inactive zymogens [21]. HGFAC was initially discovered in bovine and human serum from which it was purified [8,17,18]. Supporting these observations, the serum activity of proHGF processing was severely attenuated in sera from Hgfac knockout mice, indicating that HGFAC is responsible for the proHGF-processing activity in serum [25]. The identification of a strong HGFAC activity in serum, but not plasma, implied that HGFAC-HGF-MET signaling is a system evoked by the activation of the coagulation cascade to promote the repair of injured tissue. This is also the case for HGFAC-MSP-RON signaling [8,11,21]. Indeed, proHGFAC is efficiently activated by thrombin, particularly in the presence of dextran sulfate, chondroitin sulfate and heparin [21] (Figure 1). These negatively-charged macromolecules are rich in the pericellular microenvironment as glycosaminoglycans. Of note, HGF is a heparin-binding growth factor and activated HGFAC acquires heparin affinity [18]. Thus, these proteins may co-localize with pericellular glycosaminoglycans, thereby efficiently forming localized machinery for the activation of HGF-MET signaling in the tissue microenvironment upon tissue injury (Figure 1).
The above hypothesis was confirmed by Miyazawa et al., who showed that robust activation of proHGF occurred exclusively at the site of tissue injury in their rat models of CCl4-induced liver injury and HgCl2-induced renal injury [9]. In these models, the processing of proHGF to active HGF was significantly suppressed by an anti-HGFAC neutralizing antibody [26]. Similarly, tissue injury induced robust activation of proHGF in acetic acid-induced and dextran sodium sulfate-induced mouse colitis models, and the activation was significantly attenuated in Hgfac-deleted mouse intestine accompanied with retarded repair and decreased survival of the mice [25]. HGFAC zymogen can also be activated efficiently by tissue proteases such as kallikrein-like peptidases (KLK), particularly KLK-4 or KLK-5 [27]. Furthermore, Hgf mRNA levels were increased in stromal cells by stimulation of inflammatory cytokines and growth factors, for example interleukin-1 (IL-1), tumor necrosis factor-α, platelet-derived growth factor and transforming growth factor-α [28]. Therefore, in addition to tissue injury-induced activation of HGFAC, post-injury inflammation would orchestrate the sustained activation of proHGF at the site of tissue injury (Figure 1).
Experimental evidence supports the significance of inflammation in sustaining HGF activities. In rat liver injury models, CCl4 treatment induced significant activation of proHGF in the liver tissue [9]. However, partial hepatectomy did not induce robust proHGF activation [29]. In accordance with those findings, we observed that liver regeneration after CCl4-induced damage was delayed in Hgfac knockout mice, whereas that after partial hepatectomy was not affected by the deletion of Hgfac (Figure 2). Taken altogether, we propose that HGFAC-dependent sustained HGF-MET signaling is a system evoked by tissue damage and orchestrated by the subsequent inflammation to accelerate the repair process. In contrast, HGFAC is not essential for normal development or tissue homeostasis, so that Hgfac knockout mice showed no developmental defects and matured normally and were fertile [25].
4. Emerging Alarmin Function of HGFA upon Tissue Injury
Recently, Rodgers et al. observed activated circulating HGFAC in plasma after experimental tissue injury in mice [30]. They revealed that circulating HGFAC relays a signal to quiescent muscle stem cells and fibro-adipogenic progenitors in non-injured tissues to stimulate their transition from the G0 state to a primed GAlert state, which is mediated by HGF-MET-mTORC1 signaling (Figure 1) [30,31]. Therefore, HGFAC likely stimulates the G0 → GAlert transition of stem cells through activation of pericellular proHGF in the stem cell niche. Other stem cells such as epidermal, mesenchymal, hepatic and hematopoietic stem cells also express MET [5,6,30,32], which implies these stem cells may also respond to HGFAC in a similar fashion. Therefore, the function of HGFAC in tissue repair may not be restricted to the site of injury and HGFAC may function as a kind of alarmin. Alarmins are endogenous molecules released upon tissue damage to activate host responses, particularly inflammatory responses [33,34]. Upon tissue injury, activated HGFAC is systematically released and likely functions as a new type of alarmin to prepare for the regeneration phase (Figure 1). Very recently, high mobility group box 1 (HMGB1) complexed with CXCL12 was identified as another alarmin to transition stem cells from G0 to GAlert through CXCR4 [35]. Interestingly, inhibition of MET activity reduced the expression of surface CXCR4 in murine muscle stem cells, suggesting that the HGFAC-HGF-MET pathway and the HMGB1-CXCL12-CXCR4 pathway may be complementary for accelerated tissue regeneration (Figure 1) [35].
5. Regulation of HGFAC Activity by Endogenous Proteinase Inhibitors
To date, two endogenous proteinase inhibitors have been shown to regulate HGFAC activity in a cell-based assay and in vivo. One is protein C inhibitor (PCI, also known as SERPINA5), a circulating serpin-type inhibitor produced by the liver and also locally expressed by reproductive organs [36]. The other one is a Kunitz-type type 1 transmembrane inhibitor, HGFAC inhibitor type 1 (HAI-1)/serine peptidase inhibitor, Kunitz type 1 (SPINT1). It is widely expressed by epithelial cells and regulates HGFAC activity in the pericellular microenvironment [16,37,38]. HAI-1/SPINT1 has two extracellular Kunitz-type serine proteinase inhibitor domains: an N-terminal Kunitz domain (KD1) and a C-terminal Kunitz domain (KD2) (Figure 3). HAI-2/SPINT2 is another type 1 transmembrane Kunitz-type inhibitor with Kunitz domains homologous to those of HAI-1/SPINT1. Although HAI-2/SPINT2 was also identified as a protein efficiently inhibiting HGFAC [39], the physiological role of HAI-2/SPINT2 in the regulation of HGFAC is still a matter of debate [16]. Evidence indicates that only HAI-1/SPINT1 can complex with activated HGFAC on the surface of epithelial cells that are expressing both HAIs, suggesting that HAI-2/SPINT2 does not function as an HGFAC inhibitor on the cell surface [40]. Furthermore, HAI-2/SPINT2 immunoreactivity is exclusively observed in the cytoplasm [16,41,42], likely residing in the endoplasmic reticulum [43]. Those findings suggest a primary role of HAI-2/SPINT2 for regulation of serine proteinase activities in the endoplasmic reticulum and intracellular secretory pathways. Thus, HAI-2/SPINT2 may not be a physiological inhibitor of serum HGFAC. On the other hand, HAI-2/SPINT2 may function as an HGFAC inhibitor to suppress HGF-MET signaling in cancer tissues [44]. Further studies are required to define the roles of HAI-2/SPINT2 in HGF-MET signaling in vivo.
Thrombin catalyzes proHGFAC in the presence of heparin, which appears to be inhibited by PCI. More importantly, PCI directly inhibits HGFAC in the absence of heparin [36]. The in vivo role of PCI in the regulation of HGFAC activity was further demonstrated in a partial hepatectomy model using human PCI-transgenic mice that mimic PCI expression in humans [45]. The PCI-HGFAC complex was detectable in human plasma and the concentration of the complex was significantly higher in hepatocellular carcinoma patients (60 ± 20 pM) than in normal individuals (27 ± 10 pM) [36]. Therefore, PCI serves as a major circulating inhibitor of HGFAC.
On the other hand, HAI-1/SPINT1 is a cell surface inhibitor of HGFAC that regulates pericellular HGFAC activity [16,40]. It also regulates cell surface activators of proHGF, such as matriptase and hepsin [14,16]. In contrast to PCI that forms an irreversible complex with HGFAC, the inhibition of HGFAC by HAI-1/SPINT1 is mediated by a reversible complex formation of KD1 with the catalytic site of active HGFAC [40]. It is noteworthy that the cell surface HAI-1/SPINT1-HGFAC complex can be released by the cleavage of HAI-1/SPINT1 between KD2 and the transmembrane domain, followed by recovery of substantial HGFAC activity (around 70% of the initial activity) in the pericellular space [40]. This phenomenon is caused by markedly decreased affinity of the shed form HAI-1/SPINT1 (sHAI-1) compared to that of the membrane-anchored form to active HGFAC (Figure 3) [46]. This observation implies paradoxical functions of HAI-1/SPINT1 in regard to pericellular HGFAC activity. HAI-1/SPINT1 is a cell surface inhibitor of HGFAC, but may serve as a reservoir of active HGFAC supplying pericellular HGFAC activity if the ectodomain shedding of HAI-1/SPINT1 is increased. This shedding is metalloproteinase-dependent and accelerated by IL-1 stimulation [40]. Membrane-type 1 matrix metalloproteinase (MT1-MMP, also known as MMP14) and MMP-7 reportedly cleave Gly451/Leu452 of HAI-1/SPINT1 to release the low affinity form sHAI-1 having both KD1 and KD2 [47,48]. ADAMs (a disintegrin and metalloprotease) may also be involved in shedding, which should be clarified in future studies. It should be noted that a short form of sHAI-1 that has only KD1 efficiently inhibits HGFAC, indicating that KD1 is the functional domain for the inhibition of HGFAC; KD2 with a free N-terminus interferes with the binding of KD1 to HGFAC [46,49,50].
6. Impaired HGFAC Function and Diseases
Active HGF is an important effector of cellular survival and migration as well as anti-fibrotic actions [3,4,5]. Consequently, impaired proHGF activation in damaged tissue may significantly disturb subsequent tissue repair. For example, Hgfac knockout mice are more susceptible to acetic acid-induced or dextran sulfate sodium-induced acute colitis compared to wild-type mice, and initial restitution by epithelial cells on the ulcer surface was significantly delayed in the mutant mice [25]. This restitution step likely requires HGF activity (through MET signaling) to potently stimulate proliferation and migration of gastrointestinal epithelial cells [4]. In fact, epithelial cells undergoing restitution on gastrointestinal ulcers showed enhanced MET phosphorylation in human tissue sections [51]. In accordance with these observations, the relationship between a single nucleotide polymorphism (SNP) in the HGFAC gene and gastrointestinal mucosal disorders has been suggested in humans. A reverse genetic study using electronic medical records indicated a positive relationship between missense SNP in the HGFAC gene (rs16844401; Arg516Gln in HGFAC isoform 1 that is identical to Arg509Gln in isoform 2 [11]) and a clinical diagnosis related to gastrointestinal bleeding [52]. In addition, a genome-wide screening study indicated another SNP (rs2073505; synonymous codon) may increase susceptibility for inflammatory bowel disease [53].
Experimental evidence suggests that an imbalance between HGFAC and HAIs may be involved in idiopathic pulmonary fibrosis (IPF) [54]. Lung fibroblasts normally express proHGF, HGFAC, HAI-1/SPINT1 and HAI-2/SPINT2. In contrast, those from IPF patients have a reduced capacity to activate proHGF in vitro, a consequence of a low expression level of HGFAC and high levels of HAI-1/SPINT1 and HAI-2/SPINT2 [55]. Similar imbalances were also suggested in a mouse bleomycin-induced lung injury model, in which lung Spint1 mRNA expression and HAI-1/SPINT1 content in the bronchoalveolar lavage fluid were markedly increased compared to control mice. Thus, during fibrosis, an imbalance between HGFAC and HAI-1 became apparent, accompanying decreased proHGF activation [56]. In a study of chronic skin wounds, the beneficial effects of increased HGF and HGFAC expression could be blocked by increased expression of HAI-1/SPINT1 [57]. In addition, enhanced immunoreactivity of HAI-1/SPINT1 occurs in the gastrointestinal epithelium at the edge of ulcers [40], periportal hepatocytes of advanced primary biliary cholangitis (PBC), fulminant hepatitis [38] and also in the interlobular biliary epithelium of PBC patients [58]. However, it remains to be clarified whether augmented HAI-1/SPINT1 expression in fact suppresses the HGFAC-mediated proHGF activation in these cells. Another interesting relationship was observed between HGFAC expression and epilepsy. Decreased cortical HGFAC mRNA levels were correlated with increased frequency of temporal lobe epilepsy [59].
On the other hand, excess and persistent HGFAC activity likely occurs in tumor tissues, as tissue destruction, inflammation and enhanced procoagulant activity are unavoidable in solid tumors [60,61,62]. Considering the important roles of HGF-MET signaling in invasive growth, drug resistance and maintaining the stem cell phenotype of tumor cells, targeting proHGF activation may provide additional options to overcome the malignant phenotypes of tumor cells. The significance of HGFAC in tumor tissues has been reviewed elsewhere [15,61]. Although cell surface proHGF-activating TTSPs are primary targets to suppress excess proHGF activation in solid tumors [63], additional targeting of HGFAC would also be critically required [11,16,63].
7. Conclusions and Future Perspectives
There exist redundant mechanisms for activation of proHGF in vivo, including serum serine proteinases such as HGFAC, factor XIIa and XIa, as well as cellular TTSPs, such as matriptase and hepsin [16]. However, as summarized above, HGFAC is a primary activator of proHGF at the site of tissue injury, promoting accelerated healing of the damaged tissue. Moreover, HGFAC generated in injured tissues may function as alarmin that relays a signal to quiescent tissue stem cells distal to the injured site to stimulate their transition into a primed GAlert state [30]. Therefore, pretreatment with recombinant HGFAC may be a promising option to accelerate the repair process after a surgical procedure. Furthermore, recombinant HGFAC may also have relevance in therapies of patients with refractory wounds or persistent inflammation.
In summary, HGFAC is a proteinase that links tissue injury with subsequent regeneration and repair of tissues. The information described in this review provides opportunities to develop novel targeted approaches to accelerate tissue repair and treatment of refractory wounds.
Author Contributions
All authors contributed to this review with equal effort.
Funding
This research was funded by the Japan Society for the Promotion of Science KAKENHI (Grant No. 16H05175 to H.K. and 17K08764 to T.F.).
Acknowledgments
The authors wish to thank Takeshi Shimomura and Makiko Kawaguchi for their helpful comments.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| HGF | Hepatocyte growth factor |
| HGFAC | Hepatocyte growth factor activator |
| KLK | Kallikrein-related peptidase |
| uPA | Urokinase-type plasminogen activator |
| HAT | Human airway trypsin-like protease |
| FBS | Fetal bovine serum |
| MSP | Macrophage stimulating protein |
| HMGB1 | High mobility group box 1 |
| PCI | Protein C inhibitor |
| HAI-1 | Hepatocyte growth factor activator inhibitor type 1 |
| SPINT1 | Serine peptidase inhibitor, Kunitz type 1 |
| HAI-2 | Hepatocyte growth factor activator inhibitor type 2 |
| SPINT2 | Serine peptidase inhibitor, Kunitz type 2 |
| KD | Kunitz domain |
| MANEC | N-terminus with eight-cysteines |
| PKD | Polycystic kidney disease |
| LDL-Ra | Low density lipoprotein receptor class A |
| MMP | Matrix metalloprotease |
| MT1-MMP | Membrane type-1 matrix metalloprotease |
References
- Gohda, E.; Tsubouchi, H.; Nakayama, H.; Hirono, S.; Sakiyama, O.; Takahashi, K.; Miyazaki, H.; Hashimoto, S.; Daikuhara, Y. Purification and partial characterization of hepatocyte growth factor from plasma of a patient with fulminant hepatic failure. J. Clin. Investig. 1988, 81, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Weidner, K.M.; Arakaki, N.; Hartmann, G.; Vandekerckhove, J.; Weingart, S.; Rieder, H.; Fonatsch, C.; Tsubouchi, H.; Hishida, T.; Daikuhara, Y.; et al. Evidence for the identity of human scatter factor and human hepatocyte growth factor. Proc. Natl. Acad. Sci. USA 1991, 88, 7001–7005. [Google Scholar] [CrossRef] [PubMed]
- Tsubouchi, H.; Gohda, E.; Strain, A.J.; Daikuhara, Y. The role of HGF-SF in animal and human hepatic physiology and pathology. EXS 1993, 65, 251–274. [Google Scholar] [PubMed]
- Ido, A.; Numata, M.; Kodama, M.; Tsubouchi, H. Mucosal repair and growth factors: Recombinant human hepatocyte growth factor as an innovative therapy for inflammatory bowel disease. J. Gastroenterol. 2005, 40, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Sakai, K.; Nakamura, T.; Matsumoto, K. Hepatocyte growth factor twenty years on: Much more than a growth factor. J. Gastroenterol. Hepatol. 2011, 26 (Suppl. 1), 188–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, K.; Umitsu, M.; De Silva, D.M.; Roy, A.; Bottaro, D.P. Hepatocyte growth factor/MET in cancer progression and biomarker discovery. Cancer Sci. 2017, 108, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Naka, D.; Ishii, T.; Yoshiyama, Y.; Miyazawa, K.; Hara, H.; Hishida, T.; Kidamura, N. Activation of hepatocyte growth factor by proteolytic conversion of a single chain form to a heterodimer. J. Biol. Chem. 1992, 267, 20114–20119. [Google Scholar] [PubMed]
- Kataoka, H.; Kawaguchi, M. Hepatocyte growth factor activator (HGFA): Pathophysiological functions in vivo. FEBS J. 2010, 277, 2230–2237. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, K.; Shimomura, T.; Naka, D.; Kitamura, N. Proteolytic activation of hepatocyte growth factor in response to tissue injury. J. Biol. Chem. 1994, 269, 8966–8970. [Google Scholar] [PubMed]
- Miyazawa, K. Hepatocyte growth factor activator (HGFA): A serine protease that links tissue injury to activation of hepatocyte growth factor. FEBS J. 2010, 277, 2208–2214. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, H.; Shimomura, T. HGF activator (HGFA) and its inhibitors HAI-1 and HAI-2: Key players in tissue repair and cancer. In Extracellular Targeting of Cell Signaling in Cancer; Janetka, J.W., Benson, R.M., Eds.; Wiley: West Sussex, UK, 2018; pp. 69–90. ISBN 978-1-119-30018-2. [Google Scholar]
- Naldini, L.; Tamagnone, L.; Vigna, E.; Sachs, M.; Hartmann, G.; Birchmeier, W.; Daikuhara, Y.; Tsubouchi, H.; Blasi, F.; Comoglio, P.M. Extracellular proteolytic cleavage by urokinase is required for activation of hepatocyte growth factor/scatter factor. EMBO J. 1992, 11, 4825–4833. [Google Scholar] [CrossRef] [PubMed]
- Naldini, L.; Vigna, E.; Bardelli, A.; Follenzi, A.; Galimi, F.; Comoglio, P.M. Biological activation of pro-HGF (hepatocyte growth factor) by urokinase is controlled by a stoichiometric reaction. J. Biol. Chem. 1995, 270, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Owen, K.A.; Qiu, D.; Alves, J.; Schumacher, A.M.; Kilpatrick, L.M.; Li, J.; Harris, J.L.; Ellis, V. Pericellular activation of hepatocyte growth factor by the transmembrane serine proteases matriptase and hepsin, but not by the membrane-associated protease uPA. Biochem. J. 2010, 426, 219–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawaguchi, M.; Kataoka, H. Mechanisms of hepatocyte growth factor activation in cancer tissues. Cancers 2014, 6, 1890–1904. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, H.; Kawaguchi, M.; Fukushima, T.; Shimomura, T. Hepatocyte growth factor activator inhibitors (HAI-1 and HAI-2): Emerging key players in epithelial integrity and cancer. Pathol. Int. 2018, 68, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, T.; Ochiai, M.; Kondo, J.; Morimoto, Y. A novel protease obtained from FBS-containing culture supernatant, that processes single chain form hepatocyte growth factor to two chain form in serum-free culture. Cytotechnology 1992, 8, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, K.; Shimomura, T.; Kitamura, A.; Kondo, J.; Morimoto, Y.; Kitamura, N. Molecular cloning and sequence analysis of the cDNA for a human serine protease reponsible for activation of hepatocyte growth factor. Structural similarity of the protease precursor to blood coagulation factor XII. J. Biol. Chem. 1993, 268, 10024–10028. [Google Scholar] [PubMed]
- Donate, L.E.; Gherardi, E.; Srinivasan, N.; Sowdhamini, R.; Aparicio, S.; Blundell, T.L. Molecular evolution and domain structure of plasminogen-related growth factors (HGF/SF and HGF1/MSP). Protein Sci. 1994, 3, 2378–2394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazawa, K.; Wang, Y.; Minoshima, S.; Shimizu, N.; Kitamura, N. Structural organization and chromosomal localization of the human hepatocyte growth factor activator gene—Phylogenetic and functional relationship with blood coagulation factor XII, urokinase, and tissue-type plasminogen activator. Eur. J. Biochem. 1998, 258, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, T.; Kondo, J.; Ochiai, M.; Naka, D.; Miyazawa, K.; Morimoto, Y.; Kitamura, N. Activation of the zymogen of hepatocyte growth factor activator by thrombin. J. Biol. Chem. 1993, 268, 22927–22932. [Google Scholar] [PubMed]
- Kawaguchi, M.; Orikawa, H.; Baba, T.; Fukushima, T.; Kataoka, H. Hepatocyte growth factor activator is a serum activator of single-chain precursor macrophage-stimulating protein. FEBS J. 2009, 276, 3481–3490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, H.P.; Zhou, Y.Q.; Zhang, R.; Wang, M.H. MSP-RON signalling in cancer: Pathogenesis and therapeutic potential. Nat. Rev. Cancer 2013, 13, 466–481. [Google Scholar] [CrossRef] [PubMed]
- Sugie, S.; Mukai, S.; Yamasaki, K.; Kamibeppu, T.; Tsukino, H.; Kamoto, T. Plasma macrophage-stimulating protein and hepatocyte growth factor levels are associated with prostate cancer progression. Hum. Cell 2016, 29, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Itoh, H.; Naganuma, S.; Takeda, N.; Miyata, S.; Uchinokura, S.; Fukushima, T.; Uchiyama, S.; Tanaka, H.; Nagaike, K.; Shimomura, T.; et al. Regeneration of injured intestinal mucosa is impaired in hepatocyte growth factor activator-deficient mice. Gastroenterology 2004, 127, 1423–1435. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, K.; Shimomura, T.; Kitamura, N. Activation of hepatocyte growth factor in the injured tissues is mediated by hepatocyte growth factor activator. J. Biol. Chem. 1996, 271, 3615–3618. [Google Scholar] [CrossRef] [PubMed]
- Mukai, S.; Fukushima, T.; Naka, D.; Tanaka, H.; Osada, Y.; Kataoka, H. Activation of hepatocyte growth factor activator zymogen (pro-HGFA) by human kallikrein 1-related peptidases. FEBS J. 2008, 275, 1003–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, M.; Arakaki, N.; Tsubouchi, H.; Takada, H.; Daikuhara, Y. Enhancement of human hepatocyte growth factor production by interleukin-1 alpha and -1 beta and tumor necrosis factor-alpha by fibroblasts in culture. J. Biol. Chem. 1993, 56, 8140–8145. [Google Scholar]
- Tang, W.; Miyazawa, K.; Kitamura, N. Hepatocyte growth factor remains as an inactive single chain after partial hepatectomy or unilateral nephrectomy. FRBS Lett. 1995, 3, 220–224. [Google Scholar] [CrossRef]
- Rodgers, J.T.; Schroeder, M.D.; Ma, C.; Rando, T.A. HGFA Is an injury-regulated systemic factor that induces the transition of stem cells into GAlert. Cell Rep. 2017, 19, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, J.T.; King, K.Y.; Brett, J.O.; Cromie, M.J.; Charville, G.W.; Maguire, K.K.; Brunson, C.; Mastey, N.; Liu, L.; Tsai, C.R.; et al. mTORC1 controls the adaptive transition of quiescent stem cells from G0 to GAlert. Nature 2014, 510, 393–396. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Soto-Gutierrez, A.; Baptista, P.M.; Spee, B. Biotechnology challenges to in vitro maturation of hepatic stem cells. Gastroenterology 2018, 154, 1258–1272. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, N.; Kawakami, Y.; Maruyama, I.; Lotz, M. HMGB proteins and arthritis. Hum. Cell 2018, 31, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Dziki, J.L.; Hussey, G.; Badylak, S.F. Alarmins of the extracellular space. Semin. Immunol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.; Espirito Santo, A.I.; Zwingenberger, S.; Cai, L.; Vogl, T.; Feldmann, M.; Horwood, N.J.; Chan, J.K.; Nanchahal, J. Fully reduced HMGB1 accelerates the regeneration of multiple tissues by transitioning stem cells to GAlert. Proc. Natl. Acad. Sci. USA 2018, 115, E4463–E4472. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K. Hepatocyte growth factor activator (HGFA): Its regulation by protein C inhibitor. FEBS J. 2010, 277, 2223–2229. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, T.; Denda, K.; Kitamura, A.; Kawaguchi, T.; Kito, M.; Kondo, J.; Kagaya, S.; Qin, L.; Takata, H.; Miyazawa, K.; et al. Hepatocyte growth factor activator inhibitor, a novel Kunitz-type serine protease inhibitor. J. Biol. Chem. 1997, 272, 6370–6376. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, H.; Suganuma, T.; Shimomura, T.; Itoh, H.; Kitamura, N.; Nabeshima, K.; Koono, M. Distribution of hepatocyte growth factor activator inhibitor type 1 (HAI-1) in human tissues: Cellular surface localization of HAI-1 in simple columnar epithelium and its modulated expression in injured and regenerative tissues. J. Histochem. Cytochem. 1999, 47, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, T.; Qin, L.; Shimomura, T.; Kondo, J.; Matsumoto, K.; Denda, K.; Kitamura, N. Purification and cloning of hepatocyte growth factor activator inhibitor type 2, a Kunitz-type serine protease inhibitor. J. Biol. Chem. 1997, 272, 27558–27564. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, H.; Shimomura, T.; Kawaguchi, T.; Hamasuna, R.; Itoh, H.; Kitamura, N.; Miyazawa, K.; Koono, M. Hepatocyte growth factor activator inhibitor type 1 is a specific cell surface binding protein of hepatocyte growth factor activator (HGFA) and regulates HGFA activity in the pericellular microenvironment. J. Biol. Chem. 2000, 275, 40453–40462. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.P.; Kao, C.Y.; Chang, S.C.; Chiu, Y.L.; Chen, Y.J.; Chen, M.G.; Chang, C.C.; Lin, Y.W.; Chiang, C.P.; Wang, J.K.; et al. Tissue distribution and subcellular localizations determine in vivo functional relationship among prostasin, matriptase, HAI-1, and HAI-2 in human skin. PLoS ONE 2018, 13, e0192632. [Google Scholar] [CrossRef]
- Yamamoto, K.; Kawaguchi, M.; Shimomura, T.; Izumi, A.; Konari, K.; Honda, A.; Lin, C.Y.; Johnson, M.D.; Yamashita, Y.; Fukushima, T.; et al. Hepatocyte growth factor activator inhibitor type-2 (HAI-2)/SPINT2 contributes to invasive growth of oral squamous cell carcinoma cells. Oncotarget 2018, 9, 11691–11706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, B.R.; Steffensen, S.D.; Nielsen, N.V.; Friis, S.; Godiksen, S.; Bornholdt, J.; Soendergaard, C.; Nonboe, A.W.; Andersen, M.N.; Poulsen, S.S.; et al. Hepatocyte growth factor activator inhibitor-2 prevents shedding of matriptase. Exp. Cell Res. 2013, 319, 918–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukushima, T.; Kawaguchi, M.; Yamamoto, K.; Yamashita, F.; Izumi, A.; Kaieda, T.; Takezaki, Y.; Itoh, H.; Takeshima, H.; Kataoka, H. Aberrant methylation and silencing of the SPINT2 gene in high-grade gliomas. Cancer Sci. 2018. [Google Scholar] [CrossRef] [PubMed]
- Hamada, T.; Kamada, H.; Hayashi, T.; Nishioka, J.; Gabazza, E.C.; Isaji, S.; Uemoto, S.; Suzuki, K. Protein C inhibitor regulates hepatocyte growth factor activator-mediated liver regeneration in mice. Gut 2008, 57, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Denda, K.; Shimomura, T.; Kawaguchi, T.; Miyazawa, K.; Kitamura, N. Functional characterization of Kunitz domains in hepatocyte growth factor activator inhibitor type 1. J. Biol. Chem. 2002, 277, 14053–14059. [Google Scholar] [CrossRef] [PubMed]
- Domoto, T.; Takino, T.; Guo, L.; Sato, H. Cleavage of hepatocyte growth factor activator inhibitor-1 by membrane-type MMP-1 activates matriptase. Cancer Sci. 2012, 103, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, T.; Kimura, Y.; Hirano, H.; Higashi, S. Matrix metalloproteinase-7 induces homotypic tumor cell aggregation via proteolytic cleavage of the membrane-bound Kunitz-type inhibitor HAI-1. J. Biol. Chem. 2017, 292, 20769–20784. [Google Scholar] [CrossRef] [PubMed]
- Eigenbrot, C.; Ganesan, R.; Kirchhofer, D. Hepatocyte growth factor activator (HGFA): Molecular structure and interactions with HGFA inhibitor-1 (HAI-1). FEBS J. 2010, 277, 2215–2222. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Yuan, C.; Jensen, J.K.; Zhao, B.; Jiang, Y.; Jiang, L.; Huang, M. The crystal structure of a multidomain protease inhibitor (HAI-1) reveals the mechanism of its auto-inhibition. J. Biol. Chem. 2017, 292, 10744. [Google Scholar] [CrossRef] [PubMed]
- Nagai, M.; Takahashi, N.; Miyazawa, K.; Kawaguchi, M.; Chijiiwa, K.; Kataoka, H. Activation of MET receptor tyrosine kinase in ulcer surface epithelial cells undergoing restitution. Pathol. Int. 2008, 58, 462–464. [Google Scholar] [CrossRef] [PubMed]
- Mosley, J.D.; Van Driest, S.L.; Weeke, P.E.; Delaney, J.T.; Wells, Q.S.; Bastarache, L.; Roden, D.M.; Denny, J.C. Integrating EMR-linked and in vivo functional genetic data to identify new genotype-phenotype associations. PLoS ONE 2014, 9, e100322. [Google Scholar] [CrossRef] [PubMed]
- Ellinghaus, D.; Jostins, L.; Spain, S.L.; Cortes, A.; Bethune, J.; Han, B.; Park, Y.R.; Raychaudhuri, S.; Pouget, J.G.; Hubenthal, M.; et al. Analysis of five chronic inflammatory diseases identifies 27 new associations and highlights disease-specific patterns at shared loci. Nat. Genet. 2016, 48, 510–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menou, A.; Duitman, J.; Crestani, B. The Impaired proteases and anti-proteases balance in Idiopathic Pulmonary Fibrosis. Matrix Biol. 2018, 68–69, 382–403. [Google Scholar] [CrossRef] [PubMed]
- Marchand-Adam, S.; Fabre, A.; Mailleux, A.A.; Marchal, J.; Quesnel, C.; Kataoka, H.; Aubier, M.; Dehoux, M.; Soler, P.; Crestani, B. Defect of pro-hepatocyte growth factor activation by fibroblasts in idiopathic pulmonary fibrosis. Am. J. Respr. Crit. Care Med. 2006, 174, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Phin, S.; Marchand-Adam, S.; Fabre, A.; Marchal-Somme, J.; Bantsimba-Malanda, C.; Kataoka, H.; Soler, P.; Crestani, B. Imbalance in the pro-hepatocyte growth factor activation system in bleomycin-induced lung fibrosis in mice. Am. J. Respir. Cell Mol. Biol. 2010, 42, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Conway, K.; Ruge, F.; Price, P.; Harding, K.G.; Jiang, W.G. Hepatocyte growth factor regulation: An integral part of why wounds become chronic. Wound Repair Regen. 2007, 15, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, M.; Ikeda, H.; Kataoka, H.; Nakanuma, Y. Augmented expression of hepatocytes growth factor activator inhibitor type 1 (HAI-1) in intrahepatic small bile ducts in primary biliary cirrhosis. Virchows Arch. 2006, 449, 462–471. [Google Scholar] [CrossRef] [PubMed]
- McCallum, A.P.; Gallek, M.J.; Ramey, W.; Manziello, A.; Witte, M.H.; Bernas, M.J.; Labiner, D.M.; Weinand, M.E. Cortical gene expression correlates of temporal lobe epileptogenicity. Pathophysiology 2016, 23, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, H.; Hamasuna, R.; Itoh, H.; Kitamura, N.; Koono, M. Activation of hepatocyte growth factor/scatter factor in colorectal carcinoma. Cancer Res. 2000, 60, 6148–6159. [Google Scholar] [PubMed]
- Kataoka, H.; Miyata, S.; Uchinokura, S.; Itoh, H. Roles of hepatocyte growth factor (HGF) activator and HGF activator inhibitor in the pericellular activation of HGF/scatter factor. Cancer Metast. Rev. 2003, 22, 223–236. [Google Scholar] [CrossRef]
- Dvorak, H.F. Tumors: Wounds that do not heal-redux. Cancer Immunol. Res. 2015, 3, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Janetka, J.W.; Galemmo, R.A., Jr. Inhibitors of the growth-factor activating proteases matriptase, hepsin and HGFA: Strategy for rational drug design and optimization. In Extracellular Targeting of Cell Signaling in Cancer; Janetka, J.W., Benson, R.M., Eds.; Wiley: West Sussex, UK, 2018; pp. 247–275. ISBN 978-1-119-30018-2. [Google Scholar]
Figure 1.
Tissue injury-induced activations of proHGF by HGFAC in response to tissue injury. Thrombin efficiently activates plasma-derived proHGFAC. KLK-4 and -5 also activate proHGFAC as well. Activated HGFAC is also released into the bloodstream and serves as an “alarmin” for tissue stem cells of non-injured tissue to prepare for the regeneration phase. Injured cell-derived HMGB1 also serves as a similar alarmin through CXCR4, and HGF-MET signaling upregulates CXCR4 expression in stem cells.
Figure 1.
Tissue injury-induced activations of proHGF by HGFAC in response to tissue injury. Thrombin efficiently activates plasma-derived proHGFAC. KLK-4 and -5 also activate proHGFAC as well. Activated HGFAC is also released into the bloodstream and serves as an “alarmin” for tissue stem cells of non-injured tissue to prepare for the regeneration phase. Injured cell-derived HMGB1 also serves as a similar alarmin through CXCR4, and HGF-MET signaling upregulates CXCR4 expression in stem cells.

Figure 2.
Effects of Hgfac deletion on liver regeneration and survival of C57BL6 mice after partial hepatectomy (a) and CCl4 treatment (b). (a) Effect of Hgfac deletion on liver weight gain after 70% partial hepatectomy. N = 40 for wild-type mice and 28 for Hgfac−/− mice on the 7th day. No significant differences between two groups (two way repeated-measures analysis of variance). (b) Effect of Hgfac deletion on the survival of mice after CCl4 administration (2.5 μL/g, intraperitoneal injection). Kaplan-Meier survival curve is shown. *, p = 0.0216 (log-rank test); N = 21 for each group. All mice were maintained, treated, and sacrificed in accordance with the protocols and regulations of Miyazaki University Institutional Animal Care and Use Committee.
Figure 2.
Effects of Hgfac deletion on liver regeneration and survival of C57BL6 mice after partial hepatectomy (a) and CCl4 treatment (b). (a) Effect of Hgfac deletion on liver weight gain after 70% partial hepatectomy. N = 40 for wild-type mice and 28 for Hgfac−/− mice on the 7th day. No significant differences between two groups (two way repeated-measures analysis of variance). (b) Effect of Hgfac deletion on the survival of mice after CCl4 administration (2.5 μL/g, intraperitoneal injection). Kaplan-Meier survival curve is shown. *, p = 0.0216 (log-rank test); N = 21 for each group. All mice were maintained, treated, and sacrificed in accordance with the protocols and regulations of Miyazaki University Institutional Animal Care and Use Committee.

Figure 3.
Molecular structure of HAI-1/SPINT1 and HAI-2/SPINT2. Whereas the membrane-form HAI-1/SPINT1 and sHAI-1 only with KD1 show high affinity to HGFAC, sHAI-1 having both KD1 and KD2, shows decreased affinity to HGFAC. MANEC, N-terminus with eight-cysteines domain; PKD-like, polycystic kidney disease domain-like domain; LDL-Ra, low density lipoprotein receptor class A domain; KD, Kunitz domain; MMP, matrix metalloprotease.
Figure 3.
Molecular structure of HAI-1/SPINT1 and HAI-2/SPINT2. Whereas the membrane-form HAI-1/SPINT1 and sHAI-1 only with KD1 show high affinity to HGFAC, sHAI-1 having both KD1 and KD2, shows decreased affinity to HGFAC. MANEC, N-terminus with eight-cysteines domain; PKD-like, polycystic kidney disease domain-like domain; LDL-Ra, low density lipoprotein receptor class A domain; KD, Kunitz domain; MMP, matrix metalloprotease.


{kind=link}
{kind=link}
{kind=link}
| Proteinase | Localization | RA | RA+ | Producing Organ/Tissue |
|---|---|---|---|---|
| HGFAC | blood | 1.0 | 5.0 | Liver, (brain, GI tract, etc.) |
| Factor XIIa | blood | 0.02 | 0.7 | Liver |
| Factor XIa | blood | 0.015 | NA | Liver |
| Matriptase | cell surface | 2.07 | NA | Epithelial tissues |
| Hepsin | cell surface | 0.074 | NA | Liver, Kidney, Inner ear |
| HAT | cell surface | 0.02–0.06 | NA | Respiratory epithelium |
RA, relative activity; RA+, relative activity in the presence of high molecular weight dextran sulfate; GI tract, gastrointestinal tract; HAT, human airway trypsin-like protease; NA, not applicable.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Fukushima, T.; Uchiyama, S.; Tanaka, H.; Kataoka, H. Hepatocyte Growth Factor Activator: A Proteinase Linking Tissue Injury with Repair. Int. J. Mol. Sci. 2018, 19, 3435. https://doi.org/10.3390/ijms19113435
AMA Style
Fukushima T, Uchiyama S, Tanaka H, Kataoka H. Hepatocyte Growth Factor Activator: A Proteinase Linking Tissue Injury with Repair. International Journal of Molecular Sciences. 2018; 19(11):3435. https://doi.org/10.3390/ijms19113435
Chicago/Turabian StyleFukushima, Tsuyoshi, Shuichiro Uchiyama, Hiroyuki Tanaka, and Hiroaki Kataoka. 2018. "Hepatocyte Growth Factor Activator: A Proteinase Linking Tissue Injury with Repair" International Journal of Molecular Sciences 19, no. 11: 3435. https://doi.org/10.3390/ijms19113435
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.