The Decrease in Human Endogenous Retrovirus-H Activity Runs in Parallel with Improvement in ADHD Symptoms in Patients Undergoing Methylphenidate Therapy

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
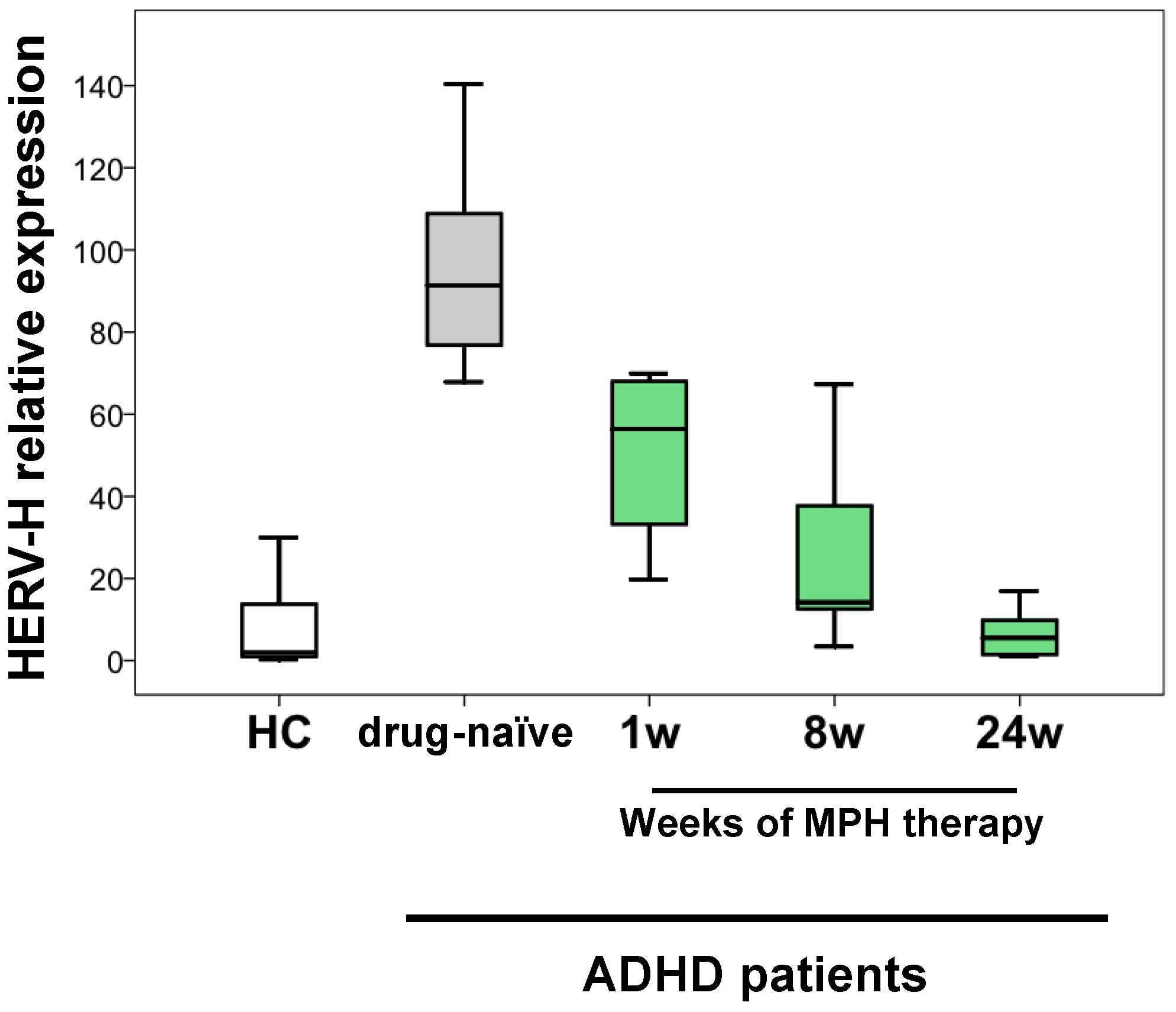
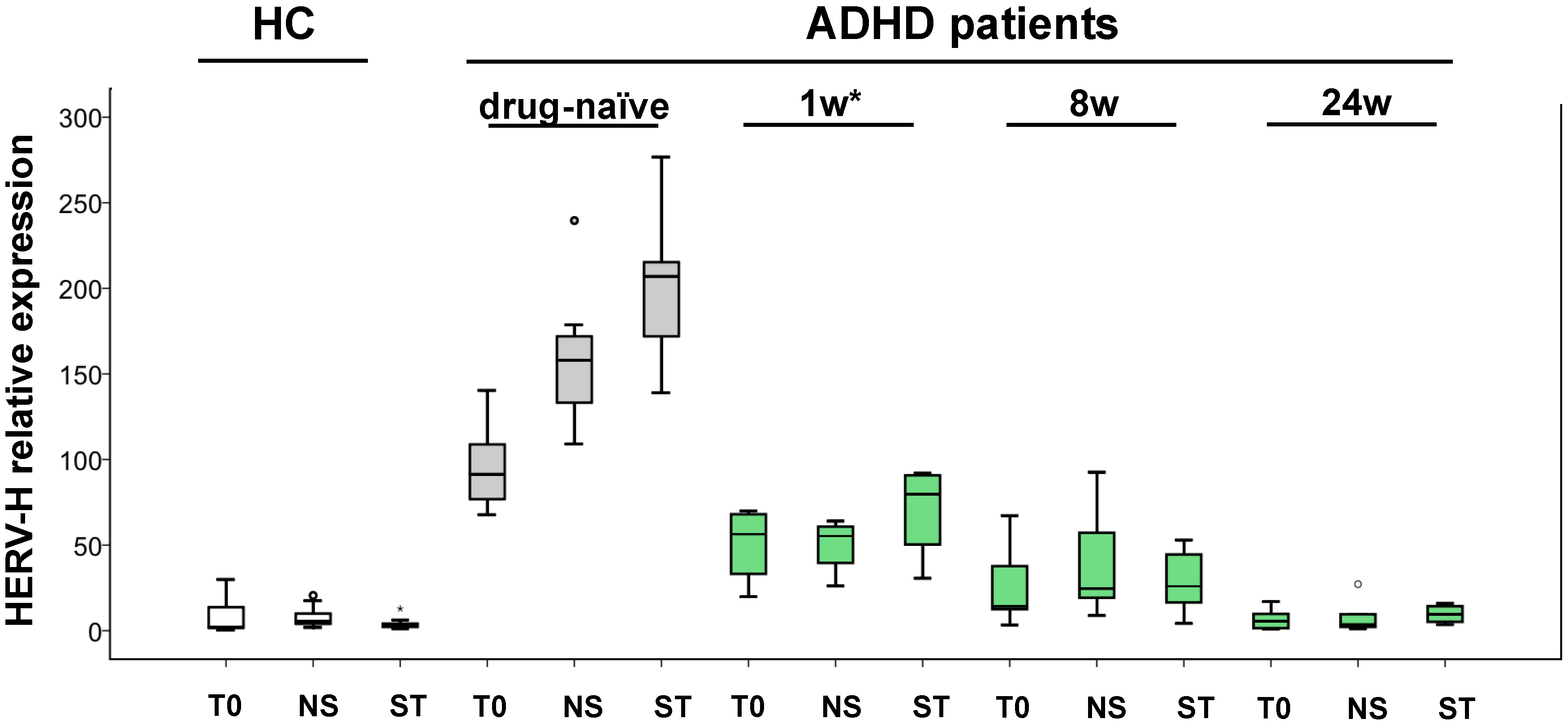
2. Results
3. Discussion
4. Materials and Methods
4.1. Participants
4.2. Clinical Assessment
4.3. Pharmacological Intervention
4.4. Samples Preparation and RT-PCR Analysis
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lander, E.S.; Linton, L.M.; Birren, B.; Nusbaum, C.; Zody, M.C.; Baldwin, J.; Devon, K.; Dewar, K.; Doyle, M.; FitzHugh, W.; et al. Initial sequencing and analysis of the human genome. Nature 2001, 409, 860921. [Google Scholar] [CrossRef]
- Hayward, A.; Katzourakis, A. Endogenous retroviruses. Curr. Biol. 2015, 25, 644–646. [Google Scholar] [CrossRef] [PubMed]
- Belshaw, R.; Pereira, V.; Katzourakis, A.; Talbot, G.; Paces, J.; Burt, A.; Tristem, M. Long-term reinfection of the human genome by endogenous retroviruses. Proc. Natl. Acad. Sci. USA 2004, 101, 4894–4899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katzourakis, A.; Rambaut, A.; Pybus, O.G. The evolutionary dynamics of endogenous retroviruses. Trends Microbiol. 2005, 13, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Feschotte, C.; Gilbert, C. Endogenous viruses: Insights into viral evolution and impact on host biology. Nat. Rev. Genet. 2012, 13, 283–296. [Google Scholar] [CrossRef] [PubMed]
- Bannert, N.; Kurth, R. The evolutionary dynamics of human endogenous retroviral families. Annu. Rev. Genom. Hum. Genet. 2006, 7, 149–173. [Google Scholar] [CrossRef] [PubMed]
- Bannert, N.; Kurth, R. Retroelements and the human genome: New perspectives on an old relation. Proc. Natl. Acad. Sci. USA 2004, 101, 14572–14579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- International Human Genome Sequencing Consortium. Finishing the euchromatic sequence of the human genome. Nature 2004, 431, 931–945. [Google Scholar] [CrossRef] [PubMed]
- Schulz, W.A.; Steinhoff, C.; Florl, A.R. Methylation of endogenous human retroelements in health and disease. Curr. Top. Microbiol. Immunol. 2006, 310, 211–250. [Google Scholar] [PubMed]
- Dewannieux, M.; Heidmann, T. Endogenous retroviruses: Acquisition, amplification and taming of genome invaders. Curr. Opin. Virol. 2013, 3, 646–656. [Google Scholar] [CrossRef] [PubMed]
- Seifarth, W.; Frank, O.; Zeilfelder, U.; Spiess, B.; Greenwood, A.D.; Hehlmann, R.; Leib-Mösch, C. Comprehensive analysis of human endogenous retrovirus transcriptional activity in human tissues with a retrovirus-specific microarray. J. Virol. 2005, 79, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Pérot, P.; Mugnier, N.; Montgiraud, C.; Gimenez, J.; Jaillard, M.; Bonnaud, B.; Mallet, F. Microarray-based sketches of the HERV transcriptome landscape. PLoS ONE 2012, 7, e40194. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.N.; Bieniasz, P.D. Reconstitution of an infectious human endogenous retrovirus. PLoS Pathog. 2007, 3, e10. [Google Scholar] [CrossRef] [PubMed]
- Jern, P.; Coffin, J.M. Effects of retroviruses on host genome function. Annu. Rev. Genet. 2008, 42, 709–732. [Google Scholar] [CrossRef] [PubMed]
- Balestrieri, E.; Argaw-Denboba, A.; Gambacurta, A.; Cipriani, C.; Bei, R.; Serafino, A.; Sinibaldi-Vallebona, P.; Matteucci, C. Human Endogenous Retrovirus K in the Crosstalk Between Cancer Cells Microenvironment and Plasticity: A New Perspective for Combination Therapy. Front. Microbiol. 2018, 9, 1448. [Google Scholar] [CrossRef] [PubMed]
- Bannert, N.; Hofmann, H.; Block, A.; Hohn, O. HERVs New Role in Cancer: From Accused Perpetrators to Cheerful Protectors. Front. Microbiol. 2018, 9, 178. [Google Scholar] [CrossRef] [PubMed]
- Volkman, H.E.; Stetson, D.B. The enemy within: Endogenous retroelements and autoimmune disease. Nat. Immunol. 2014, 15, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Marguerat, S.; Wang, W.Y.; Todd, J.A.; Conrad, B. Association of human endogenous retrovirus K-18 polymorphisms with type 1 diabetes. Diabetes 2004, 53, 852–854. [Google Scholar] [CrossRef] [PubMed]
- Küry, P.; Nath, A.; Créange, A.; Dolei, A.; Marche, P.; Gold, J.; Giovannoni, G.; Hartung, H.P.; Perron, H. Human Endogenous Retroviruses in Neurological Diseases. Trends Mol. Med. 2018, 24, 379–394. [Google Scholar] [CrossRef] [PubMed]
- Buslei, R.; Strissel, P.L.; Henke, C.; Schey, R.; Lang, N.; Ruebner, M.; Stolt, C.C.; Fabry, B.; Buchfelder, M.; Strick, R. Activation and regulation of endogenous retroviral genes in the human pituitary gland and related endocrine tumours. Neuropathol. Appl. Neurobiol. 2015, 41, 180–200. [Google Scholar] [CrossRef] [PubMed]
- Manghera, M.; Ferguson-Parry, J.; Lin, R.; Douville, R.N. NF-κB and IRF1 Induce Endogenous Retrovirus K Expression via Interferon-Stimulated Response Elements in Its 5’ Long Terminal Repeat. J. Virol. 2016, 90, 9338–9349. [Google Scholar] [CrossRef] [PubMed]
- Diem, O.; Schäffner, M.; Seifarth, W.; Leib-Mösch, C. Influence of antipsychotic drugs on human endogenous retrovirus (HERV) transcription in brain cells. PLoS ONE 2012, 7, e30054. [Google Scholar] [CrossRef] [PubMed]
- Mercorio, R.; Bonzini, M.; Angelici, L.; Iodice, S.; Delbue, S.; Mariani, J.; Apostoli, P.; Pesatori, A.C.; Bollati, V. Effects of metal-rich particulate matter exposure on exogenous and endogenous viral sequence methylation in healthy steel-workers. Environ. Res. 2017, 159, 452–457. [Google Scholar] [CrossRef] [PubMed]
- Toufaily, C.; Landry, S.; Leib-Mosch, C.; Rassart, E.; Barbeau, B. Activation of LTRs from different human endogenous retrovirus (HERV) families by the HTLV-1 tax protein and T-cell activators. Viruses 2011, 3, 2146–2159. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Hernandez, M.J.; Swanson, M.D.; Contreras-Galindo, R.; Cookinham, S.; King, S.R.; Noel, R.J., Jr.; Kaplan, M.H.; Markovitz, D.M. Expression of human endogenous retrovirus type K (HML-2) is activated by the Tat protein of HIV-1. J. Virol. 2012, 86, 7790–7805. [Google Scholar] [CrossRef] [PubMed]
- Daskalakis, M.; Brocks, D.; Sheng, Y.H.; Islam, M.S.; Ressnerova, A.; Assenov, Y.; Milde, T.; Oehme, I.; Witt, O.; Goyal, A.; et al. Reactivation of endogenous retroviral elements via treatment with DNMT- and HDAC-inhibitors. Cell Cycle 2018, 17, 811–822. [Google Scholar] [CrossRef] [PubMed]
- White, C.H.; Beliakova-Bethell, N.; Lada, S.M.; Breen, M.S.; Hurst, T.P.; Spina, C.A.; Richman, D.D.; Frater, J.; Magiorkinis, G.; Woelk, C.H. Transcriptional Modulation of Human Endogenous Retroviruses in Primary CD4+ T Cells Following Vorinostat Treatment. Front. Immunol. 2018, 9, 603. [Google Scholar] [CrossRef] [PubMed]
- Sinibaldi-Vallebona, P.; Matteucci, C.; Spadafora, C. Retrotransposon-encoded reverse transcriptase in the genesis, progression and cellular plasticity of human cancer. Cancers 2011, 3, 1141–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argaw-Denboba, A.; Balestrieri, E.; Serafino, A.; Cipriani, C.; Bucci, I.; Sorrentino, R.; Sciamanna, I.; Gambacurta, A.; Sinibaldi-Vallebona, P.; Matteucci, C. HERV-K activation is strictly required to sustain CD133+ melanoma cells with stemness features. J. Exp. Clin. Cancer Res. 2017, 36, 20. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Galindo, R.; Dube, D.; Fujinaga, K.; Kaplan, M.H.; Markovitz, D.M. Susceptibility of Human Endogenous Retrovirus Type K to Reverse Transcriptase Inhibitors. J. Virol. 2017, 91, E01309-17. [Google Scholar] [CrossRef] [PubMed]
- Cipriani, C.; Ricceri, L.; Matteucci, C.; De Felice, A.; Tartaglione, A.M.; Argaw-Denboba, A.; Pica, F.; Grelli, S.; Calamandrei, G.; SinibaldiVallebona, P.; et al. High expression of Endogenous Retroviruses from intrauterine life to adulthood in two mouse models of Autism Spectrum Disorders. Sci. Rep. 2018, 8, 629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, J.M.; Kim, H.S. Evolutionary implication of human endogenous retrovirus HERV-H family. J. Hum. Genet. 2004, 49, 215–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jern, P.; Sperber, G.O.; Ahlsén, G.; Blomberg, J. Sequence variability, gene structure, and expression of full-length human endogenous retrovirus H. J. Virol. 2005, 79, 6325–6337. [Google Scholar] [CrossRef] [PubMed]
- Guliyev, M.; Yilmaz, S.; Sahin, K.; Marakli, S.; Gozukirmizi, N. Human endogenous retrovirus-H insertion screening. Mol. Med. Rep. 2013, 7, 1305–1309. [Google Scholar] [CrossRef] [PubMed]
- Glinsky, G.V. Transposable Elements and DNA Methylation Create in Embryonic Stem Cells Human-Specific Regulatory Sequences Associated with Distal Enhancers and Noncoding RNAs. Genome Biol. Evol. 2015, 7, 1432–1454. [Google Scholar] [CrossRef] [PubMed]
- Matteucci, C.; Balestrieri, E.; Argaw-Denboba, A.; Sinibaldi-Vallebona, P. Human Endogenous Retroviruses role in cancer cell stemness. Semin. Cancer Biol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Brudek, T.; Christensen, T.; Aagaard, L.; Petersen, T.; Hansen, H.J.; Møller-Larsen, A. B cells and monocytes from patients with active multiple sclerosis exhibit increased surface expression of both HERV-H Env and HERV-W Env, accompanied by increased seroreactivity. Retrovirology 2009, 6, 104. [Google Scholar] [CrossRef] [PubMed]
- Balestrieri, E.; Arpino, C.; Matteucci, C.; Sorrentino, R.; Pica, F.; Alessandrelli, R.; Coniglio, A.; Curatolo, P.; Rezza, G.; Macciardi, F.; et al. HERVs expression in Autism Spectrum Disorders. PLoS ONE 2012, 7, e48831. [Google Scholar] [CrossRef] [PubMed]
- Balestrieri, E.; Cipriani, C.; Matteucci, C.; Capodicasa, N.; Pilika, A.; Korca, I.; Sorrentino, R.; Argaw-Denboba, A.; Bucci, I.; Miele, M.T.; et al. Transcriptional activity of human endogenous retrovirus in Albanian children with autism spectrum disorders. New Microbiol. 2016, 39, 228–231. [Google Scholar] [PubMed]
- Balestrieri, E.; Pitzianti, M.; Matteucci, C.; D’Agati, E.; Sorrentino, R.; Baratta, A.; Caterina, R.; Zenobi, R.; Curatolo, P.; Garaci, E.; et al. Human endogenous retroviruses and ADHD. World J. Biol. Psychiatry 2014, 15, 499–504. [Google Scholar] [CrossRef] [PubMed]
- American Psychiatric Association Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Press: Washington, DC, USA, 2013.
- Zametkin, A.J.; Rapoport, J.L. Neurobiology of attention deficit disorder with hyperactivity: Where have we come in 50 years? J. Am. Acad. Child Adolesc. Psychiatry 1987, 26, 676–686. [Google Scholar] [CrossRef] [PubMed]
- Seeman, P.; Madras, B.K. Anti-hyperactivity medication: Methylphenidate and amphetamine. Mol. Psychiatry 1998, 3, 386–396. [Google Scholar] [CrossRef] [PubMed]
- DuPaul, G.J.; Barkley, R.A.; Connor, D.F. Stimulants. In Attention-Deficit Hyperactivity Disorder: A Handbook for Diagnosis and Treatment, 2nd ed.; Barkley, R.A., Ed.; Guilford Press: New York, NY, USA, 1998. [Google Scholar]
- Castellanos, F.X.; Giedd, J.N.; Marsh, W.L.; Hamburger, S.D.; Vaituzis, A.C.; Dickstein, D.P.; Sarfatti, S.E.; Vauss, Y.C.; Snell, J.W.; Lange, N.; et al. Quantitative brain magnetic resonance imaging in attention-deficit hyperactivity disorder. Arch. Gen. Psychiatry 1996, 53, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Filipek, P.A.; Semrud-Clikeman, M.; Steingard, R.J.; Renshaw, P.F.; Kennedy, D.N.; Biederman, J. Volumetric MRI analysis comparing subjects having attention-deficit hyperactivity disorder with normal controls. Neurology 1997, 48, 589–601. [Google Scholar] [CrossRef] [PubMed]
- Cook, E.H., Jr.; Stein, M.A.; Krasowski, M.D.; Cox, N.J.; Olkon, D.M.; Kieffer, J.E.; Leventhal, B.L. Association of attention-deficit disorder and the dopamine transporter gene. Am. J. Hum. Genet. 1995, 56, 993–998. [Google Scholar] [PubMed]
- LaHoste, G.J.; Swanson, J.M.; Wigal, S.B.; Glabe, C.; Wigal, T.; King, N.; Kennedy, J.L. Dopamine D4 receptor gene polymorphism is associated with attention deficit hyperactivity disorder. Mol. Psychiatry 1996, 1, 121–124. [Google Scholar] [PubMed]
- Marrocco, R.T.; Davidson, M.C. Neurochemistry of attention. In The Attentive Brain; Parasuraman, R., Ed.; MIT Press: Cambridge, MA, USA, 1998. [Google Scholar]
- Riccio, C.A.; Waldrop, J.J.; Reynolds, C.R.; Lowe, P. Effects of stimulants on the continuous performance test (CPT): Implications for CPT use and interpretation. J. Neuropsychiatry Clin. Neurosci. 2001, 13, 326–335. [Google Scholar] [CrossRef] [PubMed]
- Schachar, R.; Tannock, R. Childhood hyperactivity and psychostimulants: A review of extended treatment studies. J. Child Adolesc. Psychopharmacol. 1993, 3, 81–97. [Google Scholar] [CrossRef] [PubMed]
- Swanson, J.M.; McBurnett, K.; Cooper, M.M. Effects of stimulant medication on children with attention deficit disorder: A “review of reviews”. Except. Child. 1993, 60, 154–162. [Google Scholar] [CrossRef]
- Pelham, W.E.; Burrows-MacLean, L.; Gnagy, E.M.; Fabiano, G.A.; Coles, E.K.; Wymbs, B.T.; Chacko, A.; Walker, K.S.; Wymbs, F.; Garefino, A.; et al. A Dose-Ranging Study of Behavioral and Pharmacological Treatment for Children with ADHD. J. Abnorm. Child Psychol. 2014, 42, 1019–1031. [Google Scholar] [CrossRef] [PubMed]
- D’Agati, E.; Pitzianti, M.; Balestrieri, E.; Matteucci, C.; SinibaldiVallebona, P.; Pasini, A. First evidence of HERV-H transcriptional activity reduction after methylphenidate treatment in a young boy with ADHD. New Microbiol. 2016, 39, 237–239. [Google Scholar] [PubMed]
- Pasini, A.; Sinibaldi, L.; Paloscia, C.; Douzgou, S.; Pitzianti, M.B.; Romeo, E.; Curatolo, P.; Pizzuti, A. Neurocognitive effects of methylphenidate on ADHD children with different DAT genotypes: A longitudinal open label trial. Eur. J. Paediatr. Neurol. 2013, 17, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Bonnefois, G.; Robaey, P.; Barrière, O.; Li, J.; Nekka, F. An Evaluation Approach for the Performance of Dosing Regimens in Attention-Deficit/Hyperactivity Disorder Treatment. J. Child Adolesc. Psychopharmacol. 2017, 27, 320–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, K.H.; Higgs, B.W.; Weis, S.; Song, J.; Llenos, I.C.; Dulay, J.R.; Yolken, R.H.; Webster, M.J. Effects of typical and atypical antipsychotic drugs on gene expression profiles in the liver of schizophrenia subjects. BMC Psychiatry 2009, 9, 57. [Google Scholar] [CrossRef] [PubMed]
- Dong, E.; Nelson, M.; Grayson, D.R.; Costa, E.; Guidotti, A. Clozapine and sulpiride but not haloperidol or olanzapine activate brain DNA demethylation. Proc. Natl. Acad. Sci. USA 2008, 105, 13614–13619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adriani, W.; Leo, D.; Guarino, M.; Natoli, A.; Di Consiglio, E.; De Angelis, G.; Traina, E.; Testai, E.; Perrone-Capano, C.; Laviola, G. Short-term effects of adolescent methylphenidate exposure on brain striatal gene expression and sexual/endocrine parameters in male rats. Ann. N. Y. Acad. Sci. 2006, 1074, 52–73. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Chen, C.; Yang, L.; Zhang, M.; Zhang, X.; Jia, J.; Wang, J.; Fu, Z.; Cui, X.; Ji, C.; et al. Distinct lncRNA expression profiles in the prefrontal cortex of SD rats after exposure to methylphenidate. Biomed. Pharmacother. 2015, 70, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Iskandar, B.J.; Rizk, E.; Meier, B.; Hariharan, N.; Bottiglieri, T.; Finnell, R.H.; Jarrard, D.F.; Banerjee, R.V.; Skene, J.H.; Nelson, A.; et al. Folate regulation of axonal regeneration in the rodent central nervous system through DNA methylation. J. Clin. Investig. 2010, 120, 1603–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, R.; Reif, A.; Scholz, C.J.; Weissflog, L.; Schmidt, B.; Lesch, K.P.; Jacob, C.; Reichert, S.; Heupel, J.; Volkert, J.; et al. A preliminary study on methylphenidate-regulated gene expression in lymphoblastoid cells of ADHD patients. World J. Biol. Psychiatry 2015, 16, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Hurst, T.P.; Magiorkinis, G. Epigenetic Control of Human Endogenous Retrovirus Expression: Focus on Regulation of Long-Terminal Repeats (LTRs). Viruses 2017, 9, 130. [Google Scholar] [CrossRef] [PubMed]
- Rowe, H.M.; Trono, D. Dynamic control of endogenous retroviruses during development. Virology 2011, 411, 273–287. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Sachs, F.; Ramsay, L.; Jacques, P.É.; Göke, J.; Bourque, G.; Ng, H.H. The retrovirus HERVH is a long noncoding RNA required for human embryonic stem cell identity. Nat. Struct. Mol. Biol. 2014, 21, 423–425. [Google Scholar] [CrossRef] [PubMed]
- Ohnuki, M.; Tanabe, K.; Sutou, K.; Teramoto, I.; Sawamura, Y.; Narita, M.; Nakamura, M.; Tokunaga, Y.; Nakamura, M.; Watanabe, A.; et al. Dynamic regulation of human endogenous retroviruses mediates factor-induced reprogramming and differentiation potential. Proc. Natl. Acad. Sci. USA 2014, 111, 12426–12431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyanagi-Aoi, M.; Ohnuki, M.; Takahashi, K.; Okita, K.; Noma, H.; Sawamura, Y.; Teramoto, I.; Narita, M.; Sato, Y.; Ichisaka, T.; et al. Differentiation-defective phenotypes revealed by large-scale analyses of human pluripotent stem cells. Proc. Natl. Acad. Sci. USA 2013, 110, 20569–20574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, T.; Biederman, J.; Wilens, T. Nonstimulant treatment of adult attention-deficit/hyperactivity disorder. Psychiatr. Clin. N. Am. 2004, 27, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Spencer, T.; Biederman, J.; Wilens, T.; Doyle, R.; Surman, C.; Prince, J.; Mick, E.; Aleardi, M.; Herzig, K.; Faraone, S. A large, double-blind, randomized clinical trial of methylphenidate in the treatment of adults with attention-deficit/hyperactivity disorder. Biol. Psychiatry 2005, 57, 456–463. [Google Scholar] [CrossRef] [PubMed]
- Storebø, O.J.; Pedersen, N.; Ramstad, E.; Kielsholm, M.L.; Nielsen, S.S.; Krogh, H.B.; Moreira-Maia, C.R.; Magnusson, F.L.; Holmskov, M.; Gerner, T.; et al. Methylphenidate for attention deficit hyperactivity disorder (ADHD) in children and adolescents—Assessment of adverse events in non-randomised studies. Cochrane Database Syst. Rev. 2018, 5, CD012069. [Google Scholar] [CrossRef]
- Nobile, M.; Alberti, B.; Zuddas, A. Conners’ Parents Rating Scales Revised; Adattamento Italiano, Giunti Organizzazioni Speciali: Firenze, Italy, 2007. [Google Scholar]
- Kaufman, J.; Birmaher, B.; Brent, D.; Rao, U.; Flynn, C.; Moreci, P.; Williamson, D.; Ryan, N. Schedule for Affective Disorders and Schizophrenia for School-Age Children-Present and Lifetime Version (K-SADS-PL): Initial reliability and validity data. J. Am. Acad. Child Adolesc. Psychiatry 1997, 36, 980–988. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| ADHD Patients (n = 7) | Healthy Controls (n = 12) | p Value | |
|---|---|---|---|
| Gender | males | males | 1 |
| Median age (range) years | 13 (7–17) | 11 (7–17) | 0.249 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiara, C.; Bernanda, P.M.; Claudia, M.; Elisa, D.; Tony, M.M.; Valentina, R.; Sandro, G.; Paolo, C.; Paola, S.-V.; Augusto, P.; et al. The Decrease in Human Endogenous Retrovirus-H Activity Runs in Parallel with Improvement in ADHD Symptoms in Patients Undergoing Methylphenidate Therapy. Int. J. Mol. Sci. 2018, 19, 3286. https://doi.org/10.3390/ijms19113286
Chiara C, Bernanda PM, Claudia M, Elisa D, Tony MM, Valentina R, Sandro G, Paolo C, Paola S-V, Augusto P, et al. The Decrease in Human Endogenous Retrovirus-H Activity Runs in Parallel with Improvement in ADHD Symptoms in Patients Undergoing Methylphenidate Therapy. International Journal of Molecular Sciences. 2018; 19(11):3286. https://doi.org/10.3390/ijms19113286
Chicago/Turabian StyleChiara, Cipriani, Pitzianti Maria Bernanda, Matteucci Claudia, D’Agati Elisa, Miele Martino Tony, Rapaccini Valentina, Grelli Sandro, Curatolo Paolo, Sinibaldi-Vallebona Paola, Pasini Augusto, and et al. 2018. "The Decrease in Human Endogenous Retrovirus-H Activity Runs in Parallel with Improvement in ADHD Symptoms in Patients Undergoing Methylphenidate Therapy" International Journal of Molecular Sciences 19, no. 11: 3286. https://doi.org/10.3390/ijms19113286