Synthesis of Indoleacetic Acid, Gibberellic Acid and ACC-Deaminase by Mortierella Strains Promote Winter Wheat Seedlings Growth under Different Conditions


 and
and
Abstract
:1. Introduction
2. Results
2.1. Identification and Optimal Growth Temperature of Fungal Isolates
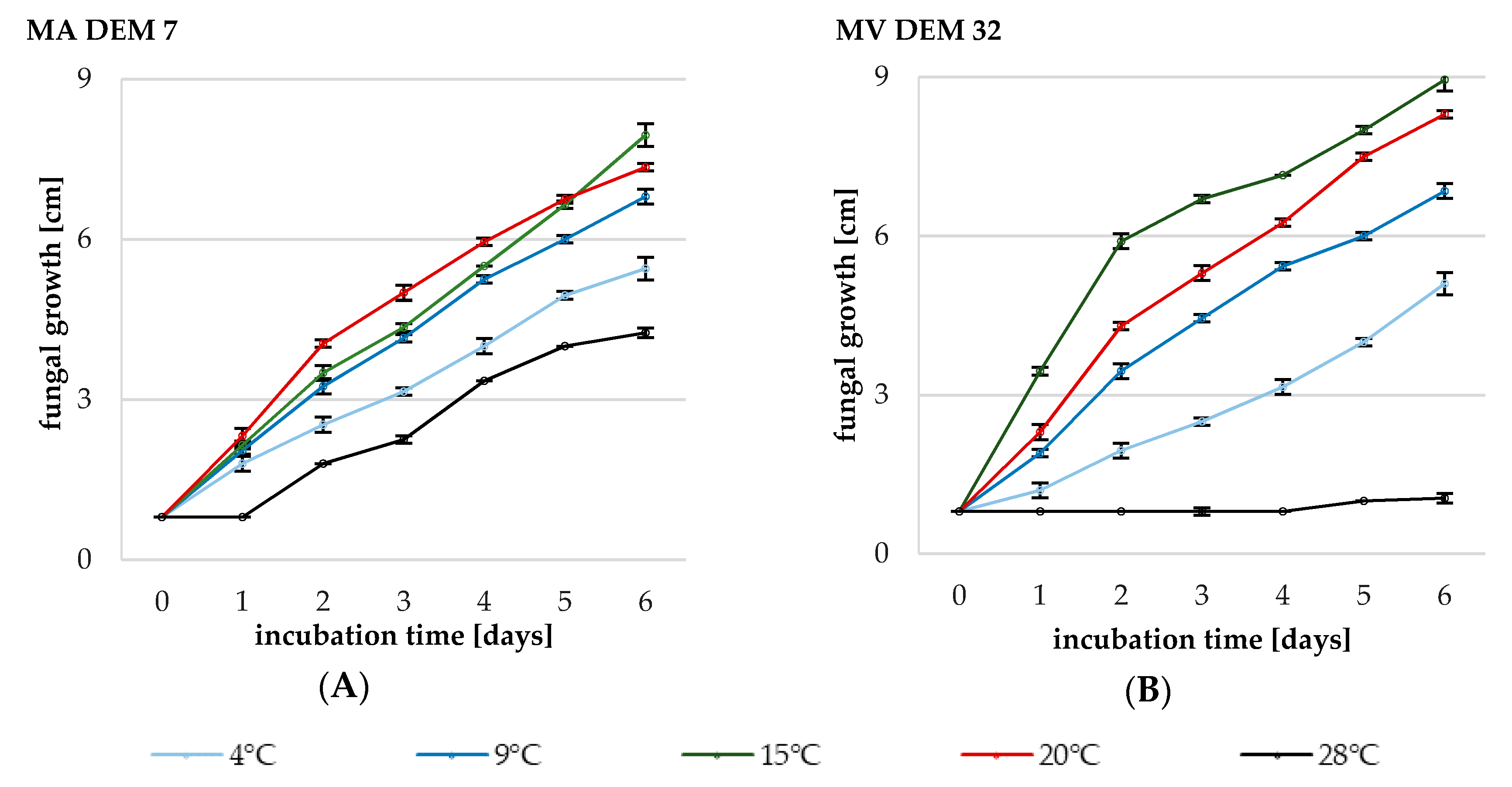
2.2. The Effect of Temperature on the Radial Growth of Mortierella Strains
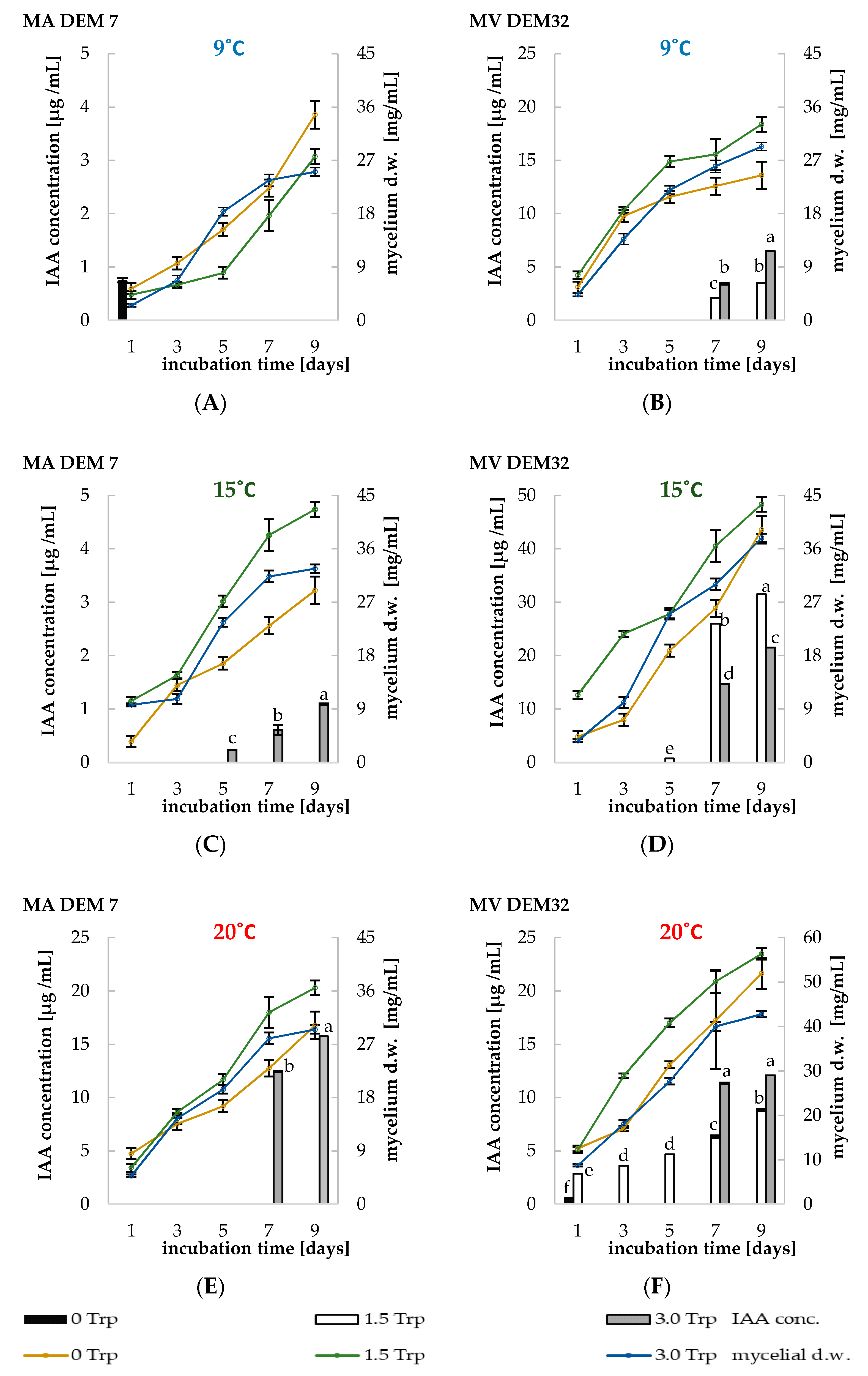
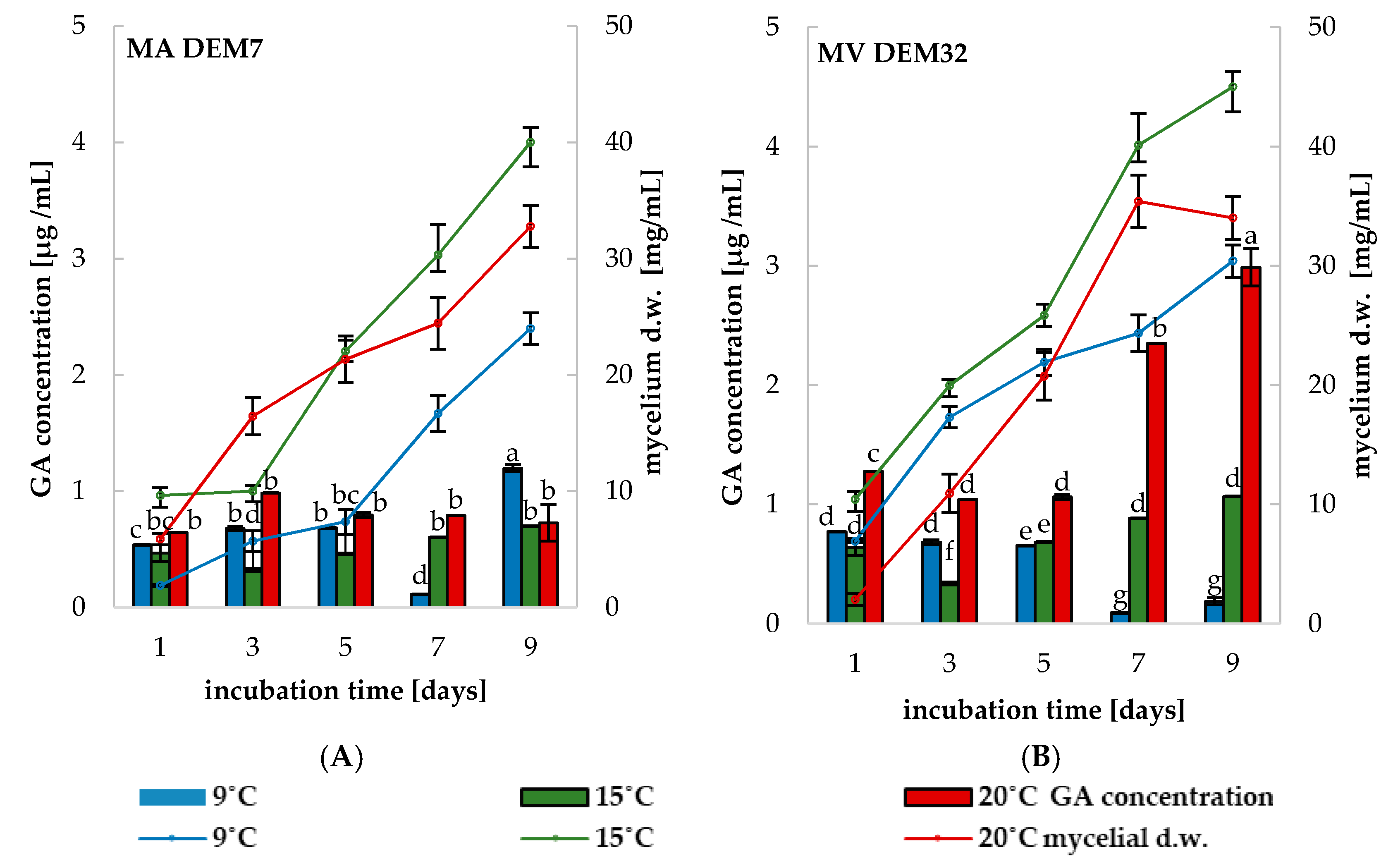
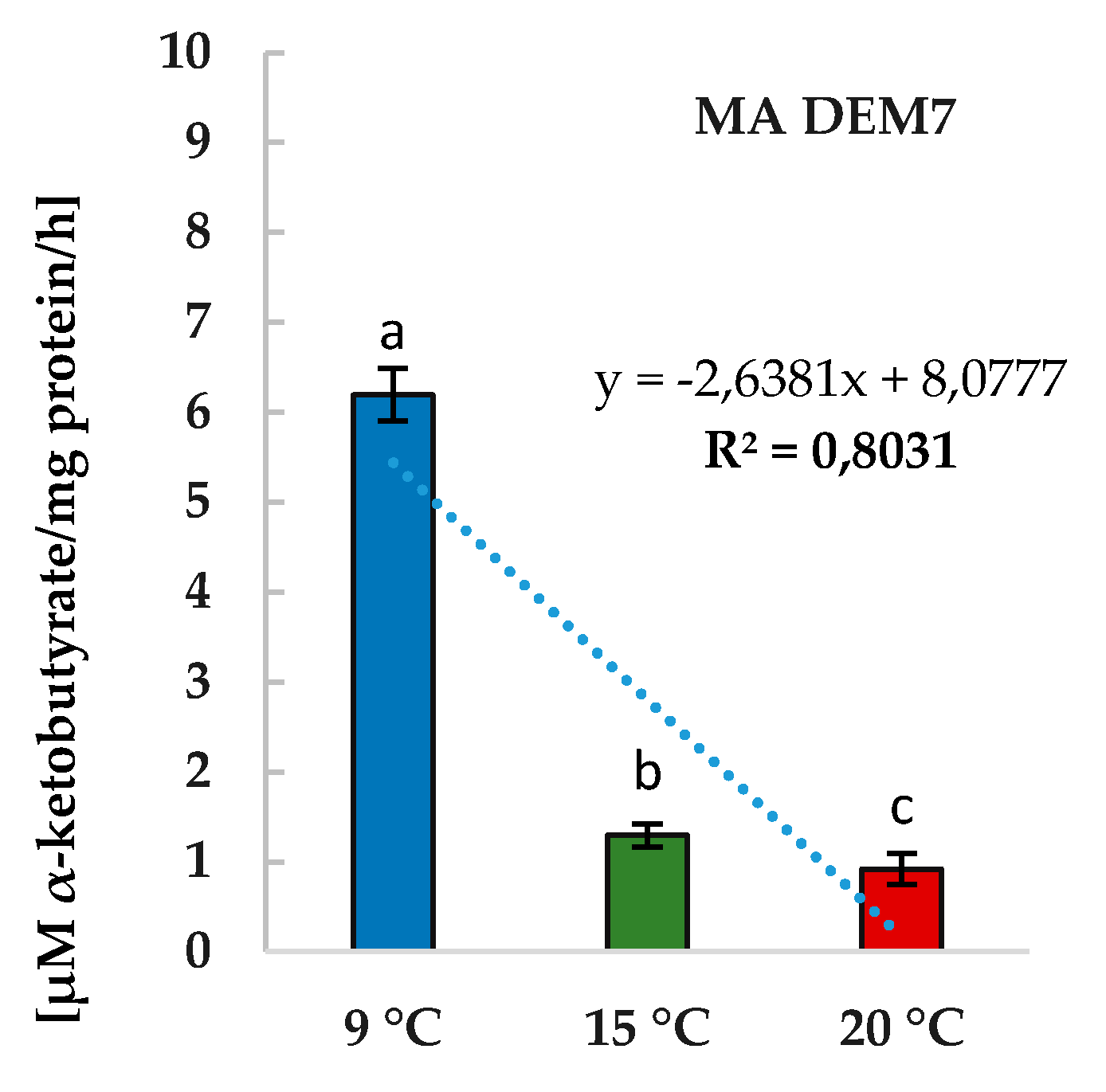
2.3. Phytoregulators Activity Assayed at Different Temperature Conditions
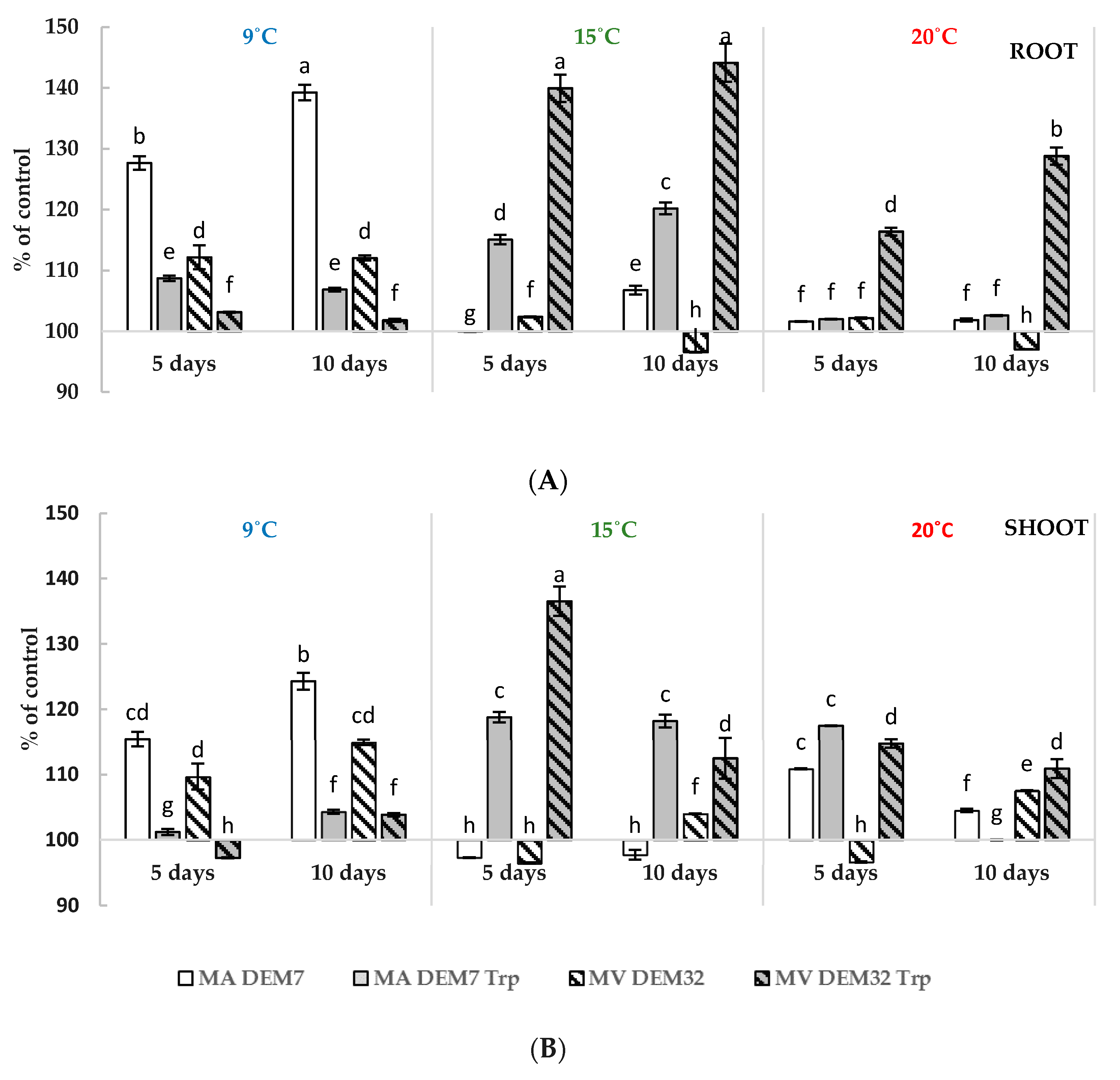
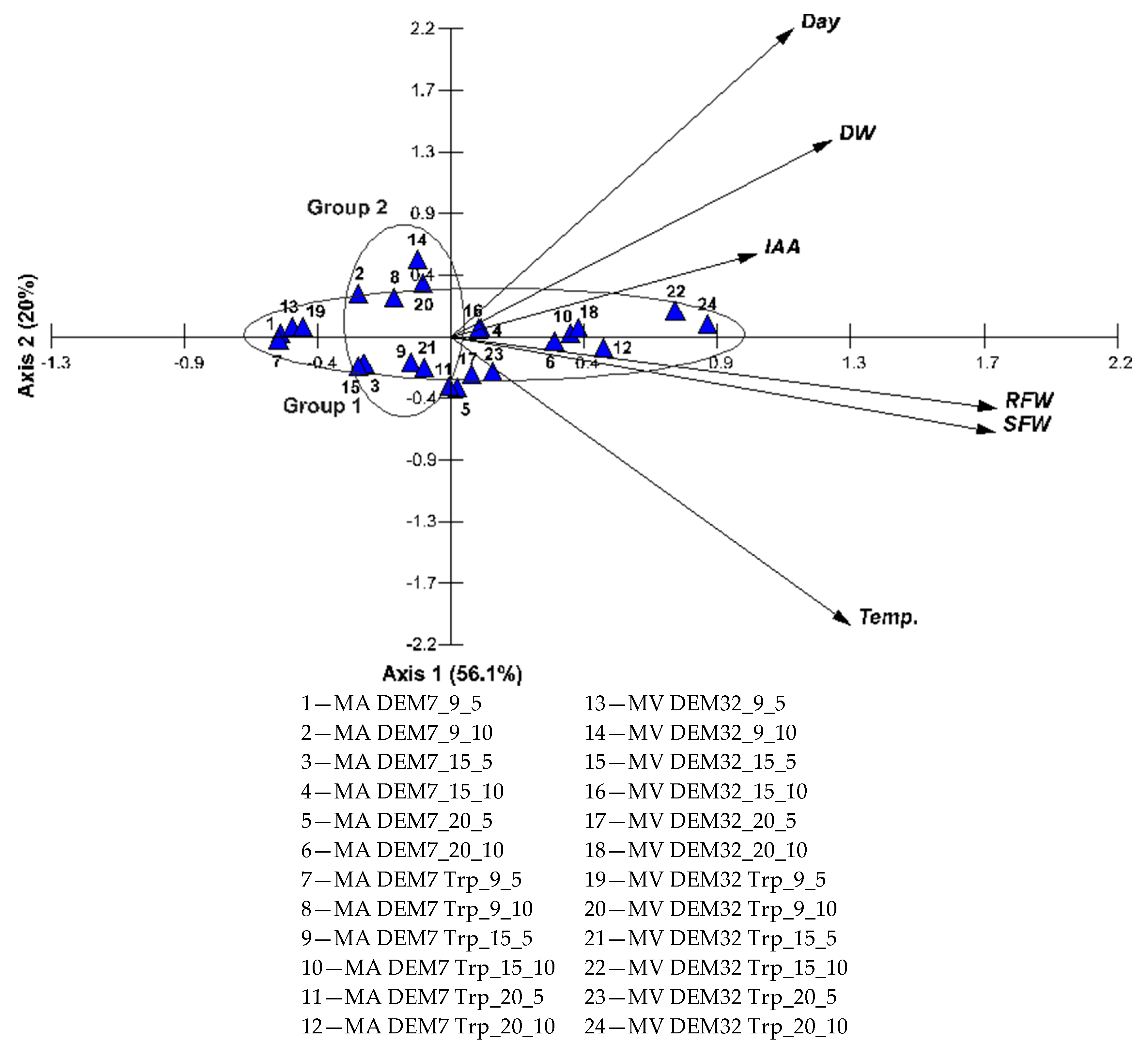
2.4. Growth of Winter Wheat Seedlings with Mortierella Strains under Various Conditions
3. Discussion
4. Materials and Methods
4.1. Microscopic and Molecular Identification of Fungal Isolates
4.2. Genomic DNA Extraction, Primers, and Sequencing
4.3. Determination of Optimal Fungal Growth Temperature
4.4. Maintenance of Strains
4.5. In Vitro Screening of Mortierella Strains for Phytoregulators Synthesis
4.5.1. Preparation of Fungal Inoculum
4.5.2. The Phytohormones and ACC-Deaminase Formation
4.6. Winter Wheat Seedlings Inoculation with Mortierella Strains—Experimental Design
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| IAA | indoleacetic acid |
| GA | gibberellic acid |
| ACC deaminase | 1-aminocyclopropane-1-carboxylate deaminase |
| ITS | internal transcribed spacer |
| PGPF | plant growth promoting fungi |
| PDA | potato dextrose agar |
| CDM | Chapek-Dox modified medium |
References
- Nagy, L.G.; Petkovits, T.; Kovács, G.M.; Voigt, K.; Vágvölgyi, C.; Papp, T. Where is the unseen fungal diversity hidden? A study of Mortierella reveals a large contribution of reference collections to the identification of fungal environmental sequences. New Phytol. 2011, 191, 789–794. [Google Scholar] [CrossRef] [PubMed]
- Yadav, D.R.; Kim, S.W.; Adhikari, M.; Um, Y.H.; Kim, H.S.; Kim, C.; Lee, H.B.; Lee, Y.S. Three New Records of Mortierella Species Isolated from Crop Field Soil in Korea. Mycobiology 2015, 43, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, V.N.; Vaz, A.B.M.; Rosa, C.A.; Rosa, L.H. Diversity and distribution of fungal communities in lakes of Antarctica. FEMS Microbiol. Ecol. 2012, 82, 459–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melo, I.S.; Santos, S.N.; Rosa, L.H.; Parma, M.M.; Silva, L.J.; Queiroz, S.C.; Pellizari, V.H. Isolation and biological activities of an endophytic Mortierella alpina strain from the Antarctic moss Schistidium antarctici. Extremophiles 2014, 18, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.T.; Lee, H.B. Characterization of a zygomycete fungus, Mortierella minutissima from freshwater of Yeongsan River in Korea. Korean J. Mycol. 2016, 44, 346–349. [Google Scholar] [CrossRef]
- Salt, G.A. The Incidence of Root-Surface Fungi on Naturally Regenerated Picea sitchensis Seedlings in Southeast Alaska. For. Int. J. For. Res. 1977, 50, 113–115. [Google Scholar] [CrossRef]
- Schmidt, S.K.; Wilson, K.L.; Meyer, A.F.; Gebauer, M.M.; King, A.J. Phylogeny and ecophysiology of opportunistic “snow molds” from a subalpine forest ecosystem. Microb. Ecol. 2008, 56, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.M.; Singh, S.K.; Yadav, L.S.; Singh, P.N.; Ravindra, R. Filamentous soil fungi from Ny-Ålesund, Spitsbergen, and screening for extracellular enzymes. Arctic 2012, 65, 45–55. [Google Scholar] [CrossRef]
- Ali, S.; Alias, A.; Siang, H.Y.; Smykla, J.; Pang, K.L.; Guo, S.Y.; Convey, P. Studies on diversity of soil microfungi in the Hornsund area, Spitsbergen. Pol. Polar Res. 2013, 34, 39–54. [Google Scholar] [CrossRef] [Green Version]
- Kurek, E.; Korniłowicz-Kowalska, T.; Słomka, A.; Melke, J. Characteristics of soil filamentous fungi communities isolated from micro-relief forms in the high Arctic tundra (Bellsund region, Spitzbergen). Pol. Polar Res. 2007, 28, 57–73. [Google Scholar]
- Wang, P.; Liu, Y.; Yin, Y.; Jin, H.; Wang, S.; Xu, F.; Zhao, S.; Geng, X. Diversity of microorganisms isolated from the soil sample surround Chroogomphus rutilus in the Beijing Region. Int. J. Biol. Sci. 2011, 7, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Crop monitory in Europe. Available online: http://publications.jrc.ec.europa.eu/repository/bitstream/JRC99692/lbam15011enn.pdf (accessed on 23 November 2015).
- Paulsen, G.M.; Heyne, E.G. Grain Production of Winter Wheat after Spring Freeze Injury 1. Agron. J. 1982, 75, 705–770. [Google Scholar] [CrossRef]
- Shaharoona, B.; Bibi, R.M.; Arshad, Z.A.; Ul-Hassan, Z. 1-Aminocylopropane-1-carboxylate (acc)-deaminase rhizobacteria extenuates acc-induced classical triple response in etiolated pea seedlings. Pak. J. Bot. 2006, 38, 1491–1499. [Google Scholar]
- Fu, S.F.; Wei, J.Y.; Chen, H.W.; Liu, Y.Y.; Lu, H.Y.; Chou, J.Y. Indole-3-acetic acid: A widespread physiological code in interactions of fungi with other organisms. Plant Signal. Behav. 2015, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Quiroz-Villareal, S.; Hernández, N.Z.; Luna-Romero, I.; Amora-Lazcano, E.; Rodríguez-Dorantes, A. Assessment of plant growth promotion by rhizobacteria supplied with tryptophan as phytohormone production elicitor on Axonopus affinis. Agric. Sci. Res. J. 2012, 2, 574–580. [Google Scholar]
- Maor, R.; Haskin, S.; Levi-Kedmi, H.; Sharon, A. In planta production of indole-3-acetic acid by Colletotrichum gloeosporioides f. sp. aeschynomene. Appl. Environ. Microb. 2004, 70, 1852–1854. [Google Scholar] [CrossRef]
- Kumar, A.; Kumar, A.; Devi, S.; Patil, S.; Payal, Ch.; Negi, S. Isolation, screening and characterization of bacteria from Rhizospheric soils for different plant growth promotion (PGP) activities: An in vitro study. Recent Res. Sci. Technol. 2012, 4, 01–05. [Google Scholar]
- Khan, A.L.; Halo, B.A.; Elyassi, A.; Ali, A.; Al-Hosni, K.; Hussain, J.; Al-Harrasi, A.; Lee, I.J. Indole acetic acid and ACC deaminase fromendophytic bacteria improves the growth of Solanum lycopersicum. Electron. J. Biotechnol. 2016, 21, 58–64. [Google Scholar] [CrossRef]
- Jaroszuk-Ściseł, J.; Kurek, E.; Trytek, M. Efficiency of indoleacetic acid, gibberellic acid and ethylene synthesized in vitro by Fusarium culmorum strains with different effects on cereal growth. Biologia 2014, 69, 281–292. [Google Scholar] [CrossRef]
- Khan, A.L.; Hamayun, M.; Kim, Y.-H.; Kang, S.M.; Lee, J.H.; Lee, I.N. Gibberellins producing endophytic Aspergillus fumigatus sp. LH02 influenced endogenous phytohormonal levels, isoflavonoids production and plant growth in salinity stress. Process Biochem. 2011, 46, 440–447. [Google Scholar] [CrossRef]
- Kurek, E.; Ozimek, E.; Sobiczewski, P.; Słomka, A.; Jaroszuk-Ściseł, J. Effect of Pseudomonas luteola on mobilization of phosphorus and growth of young apple trees (Ligol)—pot experiment. Sci. Hortic. 2013, 164, 270–276. [Google Scholar] [CrossRef]
- Osorio, N.W.; Habte, M. Synergistic influence of an arbuscular mycorrhizal fungus and a P solubilizing fungus on growth and P uptake of Leucaena leucocephala in an oxisol. Arid Land Res. Manag. 2001, 263–274. [Google Scholar] [CrossRef]
- Tamayo-Velez, A.; Osorio, N.W. Co-inoculation with an arbuscular mycorrhizal fungus and a phosphate-solubilizing fungus promotes the plant growth and phosphate uptake of avocado plantlets in a nursery. Botany 2017, 95, 539–545. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.S.; Tang, K.; Guo, S.X. The plant growth-promoting fungus (PGPF) Alternaria sp. A13 markedly enhances Salvia miltiorrhiza root growth and active ingredient accumulation under greenhouse and field conditions. Int. J. Mol. Sci. 2018, 19, 270. [Google Scholar] [CrossRef] [PubMed]
- Dyal, S.D.; Narine, S.S. Implications for the use of Mortierella fungi in the industrial production of essential fatty acids. Food Res. Int. 2005, 38, 445–467. [Google Scholar] [CrossRef]
- Grantina-Ievina, L.; Berzina, A.N.; Nikolajeva, V.; Mekss, P.; Muiznieks, I. Production of fatty acids by Mortierella and Umbelopsis species isolated from temperate climate soils. Environ. Exp. Biol. 2014, 12, 15–27. [Google Scholar]
- Nie, Z.K.; Deng, Z.T.; Zhang, A.H.; Ji, X.J.; Huang, H. Efficient arachidonic acid-rich oil production by Mortierella alpina through a three-stage fermentation strategy. Bioprocess Biosyst. Eng. 2014, 37, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chen, W.; Feng, Y.; Ren, Y.; Gu, Z. Genome characterization of the oleaginous fungus Mortierella alpina. PLoS ONE 2011, 6, e28319. [Google Scholar] [CrossRef] [PubMed]
- Domsch, K.H.; Gams, W.; Anderson, T.-H. Compendium of Soil Fungi, 2nd ed.; IHW-Verlag: Eching, Germany, 2007; pp. 1–672. ISBN 0122204026. [Google Scholar]
- Skirgiełło, A.; Zadara, M.; Ławrynowicz, M. Fungi (Mycota); Panstwowe Wydawnictwo Naukowe: Warsaw-Krakow, Poland, 1979; Volume X, pp. 218–321. [Google Scholar]
- Watanabe, T. Pictorial atlas of soil fungi and seed fungi. In Morphologies of Cultured Fungi and Key to Species, 3rd ed.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2010; ISBN 9781439804193-CAT# K10231. [Google Scholar]
- Wagner, L.; Stielow, B.; Hoffmann, K.; Petkovits, T.; Papp, T.; Vágvölgyi, C.; de Hoog, G.S.; Verkley, G.; Voigt, K. Persoonia A comprehensive molecular phylogeny of the Mortierellales (Mortierellomycotina) based on nuclear ribosomal DNA. Persoonia 2013, 30, 77–93. [Google Scholar] [CrossRef] [PubMed]
- Del Frate, G.; Caretta, G. Fungi isolated from Antarctic material. Polar Biol. 1990, 11, 1–7. [Google Scholar] [CrossRef]
- Brückner, B. Regulation of gibberellin formation by the fungus Gibberella fujikuroi. In Secondary Metabolites: Their Function and Evolution; Chadwick, D.J., Whelan, J., Eds.; Ciba Foundation Symposium, Wiley: Chichester, UK, 1992; Volume 171, pp. 129–143. [Google Scholar]
- Mosa, W.F.A.E.-G.; Sas-Paszt, L.; Frąc, M.; Trzciński, P. Microbial Products and Biofertilizers in Improving Growth and Productivity of Apple—A Review. Pol. J. Microbiol. 2016, 65, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Wani, Z.A.; Kumar, A.; Sultan, P.; Bindu, K.; Riyaz-Ul Hassan, S.; Ashraf, N. Mortierella alpina CS10E4, an oleaginous fungal endophyte of Crocus sativus L. enhances apocarotenoid biosynthesis and stress tolerance in the host plant. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wu, X.; Li, G.; Qin, P. Interactions between arbuscular mycorrhizal fungi and phosphate-solubilizing fungus (Mortierella sp.) and their effects on Kostelelzkya virginica growth and enzyme activities of rhizosphere and bulk soils at different salinities. Biol. Fertil. Soils 2011, 47, 543–554. [Google Scholar] [CrossRef]
- Ul Hassan, T.; Bano, A. The stimulatory effects of L-tryptophan and plant growth promoting rhizobacteria (PGPR) on soil health and physiology of wheat. J. Soil Sci. Plant Nutr. 2015, 15, 190–201. [Google Scholar] [CrossRef]
- Robinson, M.; Riov, J.; Sharon, A. Indole-3-acetic acid biosynthesis in Colletotrichum gloeosporioides f. sp. aeschynomene. Appl. Environ. Microbiol. 1998, 64, 5030–5032. [Google Scholar] [PubMed]
- Li, F.; Chen, L.; Redmile-Gordon, M.; Zhang, J.; Zhang, C.; Ning, Q.; Li, W. Mortierella elongata’s roles in organic agriculture and crop growth promotion in a mineral soil. Land. Degrad. Dev. 2018. [Google Scholar] [CrossRef]
- Bottini, R.; Cassán, F.; Piccoli, P. Gibberellin production by bacteria and its involvement in plant growth promotion and yield increase. Appl. Microbiol. Biotechnol. 2004, 65, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Egamberdieva, D.; Wirth, S.J.; Alqarawi, A.A.; Abd_Allah, E.F.; Hashem, A. Phytohormones and beneficial microbes: Essential components for plants to balance stress and fitness. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Hasan, H.A.H. Gibberellin and auxin production by plant root-fungi and their biosynthesis under salinity-calcium interaction. Acta Microbiol. Immunol. Hung. 2002, 48, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.; Ghosh, P.K.; Pramanik, K.; Mitra, S.; Soren, T.; Pandey, S.; Mondal, M.H.; Maiti, T.K. A halotolerant Enterobacter sp. displaying ACC deaminase activity promotes rice seedling growth under salt stress. Res. Microbiol. 2018, 169, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Davies, P.J. Plant Hormones, 2nd ed.; Physiology, Biochemistry and Molecular Biology; Kluwer Academic Publishers: Boston, MA, USA, 1995; p. 835. ISBN 978-0-7923-2985-5. [Google Scholar]
- Dash, S.; Gupta, N. Microbial bioinoculants and their role in plant growth and development. Int. J. Biotechnol. Mol. Biol. Res. 2011, 2, 232–251. [Google Scholar] [CrossRef]
- Su, Y.Y.; Qi, Y.L.; Cai, L. Induction of sporulation in plant pathogenic fungi. Mycology 2012, 3, 195–200. [Google Scholar] [CrossRef]
- Bohacz, J.; Korniłłowicz-Kowalska, T. Species diversity of keratinophilic fungi in various soil types. Cent. Eur. J. Biol. 2012, 2, 259–266. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: Guide to Methods and Applications; Innis, M., Gelfand, D., Eds.; Academic Press: Orlando, FL, USA, 1990; pp. 315–322. [Google Scholar]
- Alef, K.; Nannipieri, P. Methods in Applied Soil Microbiology and Biochemistry; Academic Press: New York, NY, USA, 1995; ISBN 9780125138406. [Google Scholar]
- Glickmann, E.; Dessaux, Y. A critical examination of the specificity of the Salkowski reagent for indolic compounds produced by phytopathogenic bacteria. Appl. Environ. Microbiol. 1995, 61, 793–796. [Google Scholar] [PubMed]
- Pilet, P.E.; Chollet, R. Colorimetric determination of indole-3-acetic acid. CR Acad. Sci. Ser. D 1970, 271, 1675–1678. [Google Scholar]
- Brückner, B.; Blechschmidt, D.; Recknagel, R.D. Optimalization of nutrient medium for biosynthesis of gibberellic acid. J. Basic Microb. 1991, 31, 243–250. [Google Scholar] [CrossRef]
- Belimov, A.A.; Dodd, I.C.; Safronova, V.I.; Shaposhnikov, A.I.; Azarova, T.S.; Makarova, N.M. Rhizobacteria that produce auxins and contain 1-amino-cyclopropane-1-carboxylic acid deaminase decrease amino acid concentrations in the rhizosphere and improve growth and yield of well-watered and water-limited potato (Solanum tuberosum). Ann. Appl. Biol. 2015, 167, 11–25. [Google Scholar] [CrossRef]
- Bradford, M. Rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Jaroszuk-Ściseł, J.; Kurek, E. Hydrolysis of fungal and plant cell walls by enzymatic complexes from cultures of Fsariumu isolates with different aggressiveness to rye (Secale cereale). Arch. Microbiol. 2012, 194, 653–665. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature | MA DEM7 | MV DEM32 |
|---|---|---|
| 4 °C | 0.77 ± 0.06 c | 0.71 ± 0.02 c |
| 9 °C | 1.0 ± 0.05 b | 1.0 ± 0.04 b |
| 15 °C | 1.19 ± 0.08 a | 1.35 ± 0.09 a |
| 20 °C | 1.09 ± 0.05 ab | 1.25 ± 0.06 ab |
| 28 °C | 0.7 ± 0.02 d | 0.041 ± 0.00 d |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ozimek, E.; Jaroszuk-Ściseł, J.; Bohacz, J.; Korniłłowicz-Kowalska, T.; Tyśkiewicz, R.; Słomka, A.; Nowak, A.; Hanaka, A. Synthesis of Indoleacetic Acid, Gibberellic Acid and ACC-Deaminase by Mortierella Strains Promote Winter Wheat Seedlings Growth under Different Conditions. Int. J. Mol. Sci. 2018, 19, 3218. https://doi.org/10.3390/ijms19103218
Ozimek E, Jaroszuk-Ściseł J, Bohacz J, Korniłłowicz-Kowalska T, Tyśkiewicz R, Słomka A, Nowak A, Hanaka A. Synthesis of Indoleacetic Acid, Gibberellic Acid and ACC-Deaminase by Mortierella Strains Promote Winter Wheat Seedlings Growth under Different Conditions. International Journal of Molecular Sciences. 2018; 19(10):3218. https://doi.org/10.3390/ijms19103218
Chicago/Turabian StyleOzimek, Ewa, Jolanta Jaroszuk-Ściseł, Justyna Bohacz, Teresa Korniłłowicz-Kowalska, Renata Tyśkiewicz, Anna Słomka, Artur Nowak, and Agnieszka Hanaka. 2018. "Synthesis of Indoleacetic Acid, Gibberellic Acid and ACC-Deaminase by Mortierella Strains Promote Winter Wheat Seedlings Growth under Different Conditions" International Journal of Molecular Sciences 19, no. 10: 3218. https://doi.org/10.3390/ijms19103218