Comparative Analysis of the Integument Transcriptomes between stick Mutant and Wild-Type Silkworms

 , and
, and
Abstract
:
1. Introduction
2. Results
2.1. Comparison of the Mechanical Properties of Cuticle between the Wild-Type and sk Mutant
2.2. Transcriptomic Sequence Analysis and Detection of Differentially-Expressed Genes (DEGs)
2.3. Validation of RNA-Seq Data by qRT-PCR
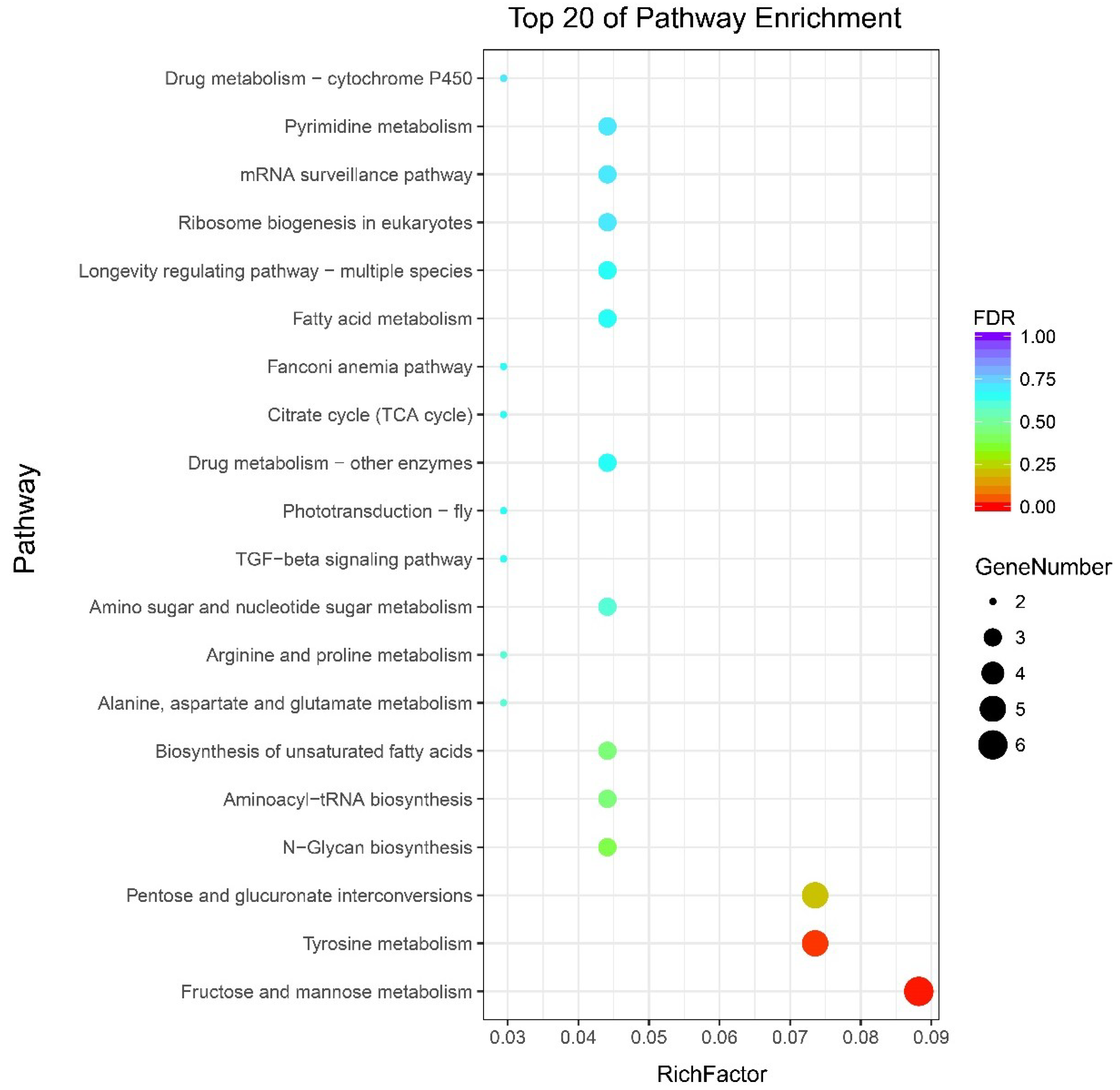
2.4. GO and KEGG Pathway Enrichment Analysis of DEGs
2.5. DEGs Specifically Related to Chitin Binding and Chitin Metabolism
2.6. DEGs Related to Hydrolase and Transferase
2.7. DEGs Related to Defense and Immune Responses
3. Discussion
3.1. The Cuticle of sk Mutants Shows a Higher Degree of Stiffness and Less Damping Oscillation Compared to the Wild-Type
3.2. Cuticular Proteins and Chitinase Are Associated with Larval Exoskeleton Shape Formation
3.3. Downregulated Genes Related to Trehalose Metabolism
3.4. Less Active Antimicrobial Peptides in sk Mutants
4. Materials and Methods
4.1. Silkworm Strains and Tissue Collection
4.2. Mechanical Properties of Cuticles
4.3. RNA Extraction and RNA-Seq
4.4. Novel Transcripts Prediction and Normalization of Gene Expression Levels and DEGs Screening
4.5. GO and KEGG Enrichment Analysis of DEGs
4.6. Quantitative Real-Time Reverse Transcription PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| L5D3 | Day 3 of the fifth instar |
| L5D2 | Day 2 of the fifth instar |
| GO | Gene ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| CPAP | Cuticle Proteins Analogous to Peritrophin |
| ChtBD2 | Peritrophin A-type chitin-binding domain |
| DEGs | Differentially-expressed genes |
| DMA | Dynamic mechanical analysis |
| DEGs | Differentially-expressed genes |
| PCA | Principal component analysis |
| FDR | Multiple hypotheses test the corrected p value |
| qRT-PCR | Quantitative real-time reverse transcription PCR |
| CBD | Chitin-binding domains |
| PM | Peritrophic membrane |
| PMPs | Peritrophic matrix proteins |
References
- Johnston, L.A.; Gallant, P. Control of growth and organ size in Drosophila. Bioessays 2002, 24, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Demontis, F.; Perrimon, N. Integration of Insulin receptor/Foxo signaling and dMyc activity during muscle growth regulates body size in Drosophila. Development 2009, 136, 983–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohni, R.; Riesgo-Escovar, J.; Oldham, S.; Brogiolo, W.; Stocker, H.; Andruss, B.F.; Beckingham, K.; Hafen, E. Autonomous control of cell and organ size by CHICO, a Drosophila homolog of vertebrate IRS1-4. Cell 1999, 97, 865–875. [Google Scholar] [CrossRef]
- Colombani, J.; Bianchini, L.; Layalle, S.; Pondeville, E.; Dauphin-Villemant, C.; Antoniewski, C.; Carre, C.; Noselli, S.; Leopold, P. Antagonistic actions of ecdysone and insulins determine final size in Drosophila. Science 2005, 310, 667–670. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Bi, H.L.; Zhang, Z.J.; Yang, Y.; Li, K.; Huang, Y.P.; Zhang, Y.; He, L. BmHpo mutation induces smaller body size and late stage larval lethality in the silkworm, Bombyx mori. Insect Sci. 2018. [Google Scholar] [CrossRef] [PubMed]
- Angelini, D.R.; Kaufman, T.C. Comparative developmental genetics and the evolution of arthropod body plans. Annu. Rev. Genet. 2005, 39, 95–119. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, S.S.; Arakane, Y.; Specht, C.A.; Moussian, B.; Boyle, D.L.; Park, Y.; Kramer, K.J.; Beeman, R.W.; Muthukrishnan, S. Knickkopf protein protects and organizes chitin in the newly synthesized insect exoskeleton. Proc. Natl. Acad. Sci. USA 2011, 108, 17028–17033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luschnig, S.; Batz, T.; Armbruster, K.; Krasnow, M.A. Serpentine and vermiform encode matrix proteins with chitin binding and deacetylation domains that limit tracheal tube length in Drosophila. Curr. Biol. 2006, 16, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Moussian, B.; Soding, J.; Schwarz, H.; Nusslein-Volhard, C. Retroactive, a membrane-anchored extracellular protein related to vertebrate snake neurotoxin-like proteins, is required for cuticle organization in the larva of Drosophila melanogaster. Dev. Dyn. 2005, 233, 1056–1063. [Google Scholar] [CrossRef] [PubMed]
- Moussian, B.; Tang, E.; Tonning, A.; Helms, S.; Schwarz, H.; Nusslein-Volhard, C.; Uv, A.E. Drosophila Knickkopf and Retroactive are needed for epithelial tube growth and cuticle differentiation through their specific requirement for chitin filament organization. Development 2006, 133, 163–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araujo, S.J.; Aslam, H.; Tear, G.; Casanova, J. Mummy/cystic encodes an enzyme required for chitin and glycan synthesis, involved in trachea, embryonic cuticle and CNS development—Analysis of its role in Drosophila tracheal morphogenesis. Dev. Biol. 2005, 288, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.; Middlebrooks, B.W.; Alexander, S.; Wasserman, S.A. Mutation of TweedleD, a member of an unconventional cuticle protein family, alters body shape in Drosophila. Proc. Natl. Acad. Sci. USA 2006, 103, 16794–16799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tajiri, R.; Ogawa, N.; Fujiwara, H.; Kojima, T. Mechanical Control of Whole Body Shape by a Single Cuticular Protein Obstructor-E in Drosophila melanogaster. PLoS Genet. 2017, 13, e1006548. [Google Scholar] [CrossRef] [PubMed]
- Clark, K.A.; Bland, J.M.; Beckerle, M.C. The Drosophila muscle LIM protein, Mlp84B, cooperates with D-titin to maintain muscle structural integrity. J. Cell Sci. 2007, 120, 2066–2077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Sullivan, K.M.; Beckingham, K. Drosophila calmodulin mutants with specific defects in the musculature or in the nervous system. Genetics 2003, 165, 1255–1268. [Google Scholar] [PubMed]
- Merzendorfer, H.; Zimoch, L. Chitin metabolism in insects: Structure, function and regulation of chitin synthases and chitinases. J. Exp. Biol. 2003, 206, 4393–4412. [Google Scholar] [CrossRef] [PubMed]
- Arakane, Y.; Hogenkamp, D.G.; Zhu, Y.C.; Kramer, K.J.; Specht, C.A.; Beeman, R.W.; Kanost, M.R.; Muthukrishnan, S. Characterization of two chitin synthase genes of the red flour beetle, Tribolium castaneum, and alternate exon usage in one of the genes during development. Insect Biochem. Mol. Biol. 2004, 34, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, G.H.; Smartt, C.T.; Kiley, L.M.; Christensen, B.M. Cloning and characterization of a chitin synthase cDNA from the mosquito Aedes aegypti. Insect Biochem. Mol. Biol. 2000, 30, 1213–1222. [Google Scholar] [CrossRef]
- Tellam, R.L.; Vuocolo, T.; Johnson, S.E.; Jarmey, J.; Pearson, R.D. Insect chitin synthase cDNA sequence, gene organization and expression. Eur. J. Biochem. 2000, 267, 6025–6043. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, J.; Park, Y.; Zhu, K.Y. Identification and characterization of two chitin synthase genes in African malaria mosquito, Anopheles gambiae. Insect Biochem. Mol. Biol. 2012, 42, 674–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arakane, Y.; Muthukrishnan, S.; Kramer, K.J.; Specht, C.A.; Tomoyasu, Y.; Lorenzen, M.D.; Kanost, M.; Beeman, R.W. The Tribolium chitin synthase genes TcCHS1 and TcCHS2 are specialized for synthesis of epidermal cuticle and midgut peritrophic matrix. Insect Mol. Biol. 2005, 14, 453–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arakane, Y.; Specht, C.A.; Kramer, K.J.; Muthukrishnan, S.; Beeman, R.W. Chitin synthases are required for survival, fecundity and egg hatch in the red flour beetle, Tribolium castaneum. Insect Biochem. Mol. Biol. 2008, 38, 959–962. [Google Scholar] [CrossRef] [PubMed]
- Bolognesi, R.; Arakane, Y.; Muthukrishnan, S.; Kramer, K.J.; Terra, W.R.; Ferreira, C. Sequences of cDNAs and expression of genes encoding chitin synthase and chitinase in the midgut of Spodoptera frugiperda. Insect Biochem. Mol. Biol. 2005, 35, 1249–1259. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.S.; Tang, B.; Chen, X.; Tian, H.; Zhang, W. Molecular cloning, expression pattern and comparative analysis of chitin synthase gene B in Spodoptera exigua. Comp. Biochem. Phys. Part B 2008, 149, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, W.; Fang, Y.; Kong, L.; Li, X.; Sima, Y.; Xu, S. Chitin synthase A: A novel epidermal development regulation gene in the larvae of Bombyx mori. Mol. Biol. Rep. 2014, 41, 4177–4186. [Google Scholar] [CrossRef] [PubMed]
- Moussian, B.; Schwarz, H.; Bartoszewski, S.; Nusslein-Volhard, C. Involvement of chitin in exoskeleton morphogenesis in Drosophila melanogaster. J. Morphol. 2005, 264, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Ostrowski, S.; Dierick, H.A.; Bejsovec, A. Genetic control of cuticle formation during embryonic development of Drosophila melanogaster. Genetics 2002, 161, 171–182. [Google Scholar] [PubMed]
- Zhang, J.; Liu, X.; Zhang, J.; Li, D.; Sun, Y.; Guo, Y.; Ma, E.; Zhu, K.Y. Silencing of two alternative splicing-derived mRNA variants of chitin synthase 1 gene by RNAi is lethal to the oriental migratory locust, Locusta migratoria manilensis (Meyen). Insect Biochem. Mol. Biol. 2010, 40, 824–833. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Arakane, Y.; Banerjee, D.; Beeman, R.W.; Kramer, K.J.; Muthukrishnan, S. Domain organization and phylogenetic analysis of the chitinase-like family of proteins in three species of insects. Insect Biochem. Mol. Biol. 2008, 38, 452–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merzendorfer, H. Chitin synthesis inhibitors: Old molecules and new developments. Insect Sci. 2013, 20, 121–138. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, X.; Arakane, Y.; Muthukrishnan, S.; Kramer, K.J.; Ma, E.; Zhu, K.Y. Comparative genomic analysis of chitinase and chitinase-like genes in the African malaria mosquito (Anopheles gambiae). PLoS ONE 2011, 6, e19899. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Lu, P.; Wang, Y.; Yin, L.; Ma, H.; Ma, G.; Chen, K.; He, Y. In silico identification of novel chitinase-like proteins in the silkworm, Bombyx mori, genome. J. Insect Sci. 2012, 12, 150. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Arakane, Y.; Beeman, R.W.; Kramer, K.J.; Muthukrishnan, S. Functional specialization among insect chitinase family genes revealed by RNA interference. Proc. Natl. Acad. Sci. USA 2008, 105, 6650–6655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Zhang, J.; Wang, Y.; Liu, X.; Ma, E.; Sun, Y.; Li, S.; Zhu, K.Y.; Zhang, J. Two chitinase 5 genes from Locusta migratoria: Molecular characteristics and functional differentiation. Insect Biochem. Mol. Biol. 2015, 58, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Cornman, R.S.; Togawa, T.; Dunn, W.A.; He, N.; Emmons, A.C.; Willis, J.H. Annotation and analysis of a large cuticular protein family with the R&R Consensus in Anopheles gambiae. BMC Genom. 2008, 9, 22. [Google Scholar]
- Futahashi, R.; Okamoto, S.; Kawasaki, H.; Zhong, Y.S.; Iwanaga, M.; Mita, K.; Fujiwara, H. Genome-wide identification of cuticular protein genes in the silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 2008, 38, 1138–1146. [Google Scholar] [CrossRef] [PubMed]
- Cornman, R.S.; Willis, J.H. Annotation and analysis of low-complexity protein families of Anopheles gambiae that are associated with cuticle. Insect Mol. Biol. 2009, 18, 607–622. [Google Scholar] [CrossRef] [PubMed]
- Rebers, J.E.; Riddiford, L.M. Structure and expression of a Manduca sexta larval cuticle gene homologous to Drosophila cuticle genes. J. Mol. Biol. 1988, 203, 411–423. [Google Scholar] [CrossRef]
- Togawa, T.; Augustine Dunn, W.; Emmons, A.C.; Willis, J.H. CPF and CPFL, two related gene families encoding cuticular proteins of Anopheles gambiae and other insects. Insect Biochem. Mol. Biol. 2007, 37, 675–688. [Google Scholar] [CrossRef] [PubMed]
- Jasrapuria, S.; Arakane, Y.; Osman, G.; Kramer, K.J.; Beeman, R.W.; Muthukrishnan, S. Genes encoding proteins with peritrophin A-type chitin-binding domains in Tribolium castaneum are grouped into three distinct families based on phylogeny, expression and function. Insect Biochem. Mol. Biol. 2010, 40, 214–227. [Google Scholar] [CrossRef] [PubMed]
- Jasrapuria, S.; Specht, C.A.; Kramer, K.J.; Beeman, R.W.; Muthukrishnan, S. Gene families of cuticular proteins analogous to peritrophins (CPAPs) in Tribolium castaneum have diverse functions. PLoS ONE 2012, 7, e49844. [Google Scholar] [CrossRef] [PubMed]
- Kucharski, R.; Maleszka, J.; Maleszka, R. Novel cuticular proteins revealed by the honey bee genome. Insect Biochem. Mol. Biol. 2007, 37, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Arakane, Y.; Lomakin, J.; Gehrke, S.H.; Hiromasa, Y.; Tomich, J.M.; Muthukrishnan, S.; Beeman, R.W.; Kramer, K.J.; Kanost, M.R. Formation of rigid, non-flight forewings (elytra) of a beetle requires two major cuticular proteins. PLoS Genet. 2012, 8, e1002682. [Google Scholar] [CrossRef] [PubMed]
- Noh, M.Y.; Muthukrishnan, S.; Kramer, K.J.; Arakane, Y. Tribolium castaneum RR-1 cuticular protein TcCPR4 is required for formation of pore canals in rigid cuticle. PLoS Genet. 2015, 11, e1004963. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith, M.R.; Shimada, T.; Abe, H. The genetics and genomics of the silkworm, Bombyx mori. Annu. Rev. Entomol. 2005, 50, 71–100. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.; Xiong, G.; Wang, R.X.; He, S.Z.; Chen, J.; Tong, X.L.; Hu, H.; Li, C.L.; Gai, T.T.; Xin, Y.Q.; et al. Mutation of a cuticular protein, BmorCPR2, alters larval body shape and adaptability in silkworm, Bombyx mori. Genetics 2014, 196, 1103–1115. [Google Scholar] [CrossRef] [PubMed]
- Xiong, G.; Tong, X.; Gai, T.; Li, C.; Qiao, L.; Monteiro, A.; Hu, H.; Han, M.; Ding, X.; Wu, S.; et al. Body Shape and Coloration of Silkworm Larvae Are Influenced by a Novel Cuticular Protein. Genetics 2017, 207, 1053–1066. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.X.; Tong, X.L.; Gai, T.T.; Li, C.L.; Qiao, L.; Hu, H.; Han, M.J.; Xiang, Z.H.; Lu, C.; Dai, F.Y. A serine protease homologue Bombyx mori scarface induces a short and fat body shape in silkworm. Insect Mol. Biol. 2018, 27, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Banno, Y.; Fujii, H.; Kawaguchi, Y.; Yamamoto, K.; Nishikawa, K.; Nishisaka, A.; Tamura, K.; Eguchi, S. A Guide to the Silkworm Mutants: 2005 Gene Name and Gene Symbol; Kyusyu University: Fukuoka, Japan, 2005; pp. 1–30. [Google Scholar]
- Jones, D.S. Dynamic mechanical analysis of polymeric systems of pharmaceutical and biomedical significance. Int. J. Pharm. 1999, 179, 167–178. [Google Scholar] [CrossRef]
- Bourne, Y.; Henrissat, B. Glycoside hydrolases and glycosyltransferases: Families and functional modules. Curr. Opin. Struct. Biol. 2001, 11, 593–600. [Google Scholar] [CrossRef]
- Henrissat, B.; Davies, G. Structural and sequence-based classification of glycoside hydrolases. Curr. Opin. Struct. Biol. 1997, 7, 637–644. [Google Scholar] [CrossRef]
- Kalf, G.F.; Rieder, S.V. The purification and properties of trehalase. J. Biol. Chem. 1958, 230, 691–698. [Google Scholar] [PubMed]
- Hehre, E.J.; Sawai, T.; Brewer, C.F.; Nakano, M.; Kanda, T. Trehalase: Stereocomplementary hydrolytic and glucosyl transfer reactions with α- and β-D-glucosyl fluoride. Biochemistry 1982, 21, 3090–3097. [Google Scholar] [CrossRef] [PubMed]
- Mori, H.; Lee, J.H.; Okuyama, M.; Nishimoto, M.; Ohguchi, M.; Kim, D.; Kimura, A.; Chiba, S. Catalytic reaction mechanism based on α-secondary deuterium isotope effects in hydrolysis of trehalose by European honeybee trehalase. Biosci. Biotechnol. Biochem. 2009, 73, 2466–2473. [Google Scholar] [CrossRef] [PubMed]
- Negishi, M.; Pedersen, L.G.; Petrotchenko, E.; Shevtsov, S.; Gorokhov, A.; Kakuta, Y.; Pedersen, L.C. Structure and function of sulfotransferases. Arch. Biochem. Biophys. 2001, 390, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Rath, V.L.; Verdugo, D.; Hemmerich, S. Sulfotransferase structural biology and inhibitor discovery. Drug Discov. Today 2004, 9, 1003–1011. [Google Scholar] [CrossRef]
- Fukuda, M.; Hiraoka, N.; Akama, T.O.; Fukuda, M.N. Carbohydrate-modifying sulfotransferases: Structure, function, and pathophysiology. J. Biol. Chem. 2001, 276, 47747–47750. [Google Scholar] [CrossRef] [PubMed]
- Axen, A.; Carlsson, A.; Engstrom, A.; Bennich, H. Gloverin, an antibacterial protein from the immune hemolymph of Hyalophora pupae. Eur. J. Biochem. 1997, 247, 614–619. [Google Scholar] [CrossRef] [PubMed]
- Hara, S.; Yamakawa, M. Moricin, a novel type of antibacterial peptide isolated from the silkworm, Bombyx mori. J. Biol. Chem. 1995, 270, 29923–29927. [Google Scholar] [PubMed]
- Hara, S.; Yamakawa, M. A novel antibacterial peptide family isolated from the silkworm, Bombyx mori. Biochem. J. 1995, 310, 651–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.H.; Park, B.S.; Yun, E.Y.; Je, Y.H.; Woo, S.D.; Kang, S.W.; Kim, K.Y.; Kang, S.K. Cloning and expression of a novel gene encoding a new antibacterial peptide from silkworm, Bombyx mori. Biochem. Biophys. Res. Commun. 1998, 246, 388–392. [Google Scholar] [CrossRef] [PubMed]
- Morishima, I.; Suginaka, S.; Ueno, T.; Hirano, H. Isolation and structure of cecropins, inducible antibacterial peptides, from the silkworm, Bombyx mori. Comp. Biochem. Physiol. B 1990, 95, 551–554. [Google Scholar] [CrossRef]
- Sugiyama, M.; Kuniyoshi, H.; Kotani, E.; Taniai, K.; Kadono-Okuda, K.; Kato, Y.; Yamamoto, M.; Shimabukuro, M.; Chowdhury, S.; Xu, J.; et al. Characterization of a Bombyx mori cDNA encoding a novel member of the attacin family of insect antibacterial proteins. Insect Biochem. Mol. Biol. 1995, 25, 385–392. [Google Scholar] [CrossRef]
- Arakane, Y.; Lomakin, J.; Beeman, R.W.; Muthukrishnan, S.; Gehrke, S.H.; Kanost, M.R.; Kramer, K.J. Molecular and functional analyses of amino acid decarboxylases involved in cuticle tanning in Tribolium castaneum. J. Biol. Chem. 2009, 284, 16584–16594. [Google Scholar] [CrossRef] [PubMed]
- Lomakin, J.; Arakane, Y.; Kramer, K.J.; Beeman, R.W.; Kanost, M.R.; Gehrke, S.H. Mechanical properties of elytra from Tribolium castaneum wild-type and body color mutant strains. J. Insect Physiol. 2010, 56, 1901–1906. [Google Scholar] [CrossRef] [PubMed]
- Lomakin, J.; Huber, P.A.; Eichler, C.; Arakane, Y.; Kramer, K.J.; Beeman, R.W.; Kanost, M.R.; Gehrke, S.H. Mechanical properties of the beetle elytron, a biological composite material. Biomacromolecules 2011, 12, 321–335. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Zhang, L.; Xiang, Z.; He, N. Expression profile of cuticular genes of silkworm, Bombyx mori. BMC Genom. 2010, 11, 173. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.L.; Tian, S.; Yang, H.; Zhou, X.; Xu, S.P.; Zhang, Z.Y.; Gong, J.; Hou, Y.; Xia, Q.Y. Genome-wide identification of chitin-binding proteins and characterization of BmCBP1 in the silkworm, Bombyx mori. Insect Sci. 2017. [Google Scholar] [CrossRef] [PubMed]
- Tellam, R.L.; Wijffels, G.; Willadsen, P. Peritrophic matrix proteins. Insect Biochem. Mol. Biol. 1999, 29, 87–101. [Google Scholar] [CrossRef]
- Dixit, R.; Arakane, Y.; Specht, C.A.; Richard, C.; Kramer, K.J.; Beeman, R.W.; Muthukrishnan, S. Domain organization and phylogenetic analysis of proteins from the chitin deacetylase gene family of Tribolium castaneum and three other species of insects. Insect Biochem. Mol. Biol. 2008, 38, 440–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barry, M.K.; Triplett, A.A.; Christensen, A.C. A peritrophin-like protein expressed in the embryonic tracheae of Drosophila melanogaster. Insect Biochem. Mol. Biol. 1999, 29, 319–327. [Google Scholar] [CrossRef]
- Behr, M.; Hoch, M. Identification of the novel evolutionary conserved obstructor multigene family in invertebrates. FEBS Lett. 2005, 579, 6827–6833. [Google Scholar] [CrossRef] [PubMed]
- Petkau, G.; Wingen, C.; Jussen, L.C.; Radtke, T.; Behr, M. Obstructor-A is required for epithelial extracellular matrix dynamics, exoskeleton function, and tubulogenesis. J. Biol. Chem. 2012, 287, 21396–21405. [Google Scholar] [CrossRef] [PubMed]
- Qu, M.; Ren, Y.; Liu, Y.; Yang, Q. Studies on the chitin/chitosan binding properties of six cuticular proteins analogous to peritrophin 3 from Bombyx mori. Insect Mol. Biol. 2017, 26, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, G.R.; Kale, G.F. The chemistry of insect hemolymph. II. Trehalose and other carbohydrates. J. Gen. Physiol. 1957, 40, 833–847. [Google Scholar] [CrossRef] [PubMed]
- Paul, M.J.; Primavesi, L.F.; Jhurreea, D.; Zhang, Y. Trehalose metabolism and signaling. Annu. Rev. Plant Biol. 2008, 59, 417–441. [Google Scholar] [CrossRef] [PubMed]
- Elbein, A.D.; Pan, Y.T.; Pastuszak, I.; Carroll, D. New insights on trehalose: A multifunctional molecule. Glycobiology 2003, 13, 17R–27R. [Google Scholar] [CrossRef] [PubMed]
- Cohen, E. Chitin synthesis and inhibition: A revisit. Pest Manag. Sci. 2001, 57, 946–950. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.C.; Xia, Q.Y.; Qian, J.F.; Liu, C.; Lin, Y.; Zha, X.F.; Xiang, Z.H. Mining single nucleotide polymorphisms from EST data of silkworm, Bombyx mori, inbred strain Dazao. Insect Biochem. Mol. Biol. 2004, 34, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.M.; Kusakabe, T.; Lee, J.M.; Tatsuke, T.; Kawaguchi, Y.; Kang, M.W.; Kang, S.W.; Kim, K.A.; Nho, S.K. Structure and expression analysis of the cecropin-E gene from the silkworm, Bombyx mori. Biosci. Biotechnol. Biochem. 2008, 72, 1992–1998. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Cheng, T.; Ye, M.; Deng, X.; Yi, H.; Huang, Y.; Tan, X.; Han, D.; Wang, B.; Xiang, Z.; et al. Functional divergence among silkworm antimicrobial peptide paralogs by the activities of recombinant proteins and the induced expression profiles. PLoS ONE 2011, 6, e18109. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhu, F.; Cao, Y.; Qiao, S. Novel expression vector for secretion of cecropin AD in Bacillus subtilis with enhanced antimicrobial activity. Antimicrob. Agents Chemother. 2009, 53, 3683–3689. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Wang, C.; Guo, X.; Li, W.; Wang, Y.; He, Q. Broad activity against porcine bacterial pathogens displayed by two insect antimicrobial peptides moricin and cecropin B. Mol. Cells 2013, 35, 106–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.; Jeong, K.W.; Lee, J.; Shin, A.; Kim, J.K.; Lee, J.; Lee, D.G.; Kim, Y. Structure-activity relationships of cecropin-like peptides and their interactions with phospholipid membrane. BMB Rep. 2013, 46, 282–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeLucca, A.J.; Bland, J.M.; Jacks, T.J.; Grimm, C.; Cleveland, T.E.; Walsh, T.J. Fungicidal activity of cecropin A. Antimicrob. Agents Chemother. 1997, 41, 481–483. [Google Scholar] [CrossRef] [PubMed]
- Eade, C.R.; Wood, M.P.; Cole, A.M. Mechanisms and modifications of naturally occurring host defense peptides for anti-HIV microbicide development. Curr. HIV Res. 2012, 10, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.B.; Mei, H.F.; Pu, Q.H.; Shen, J.; Lu, X.M.; Chu, F.J.; Zhu, J.Y. Effects of Musca domestica cecropin on the adhesion and migration of human hepatocellular carcinoma BEL-7402 cells. Biol. Pharm. Bull. 2013, 36, 938–943. [Google Scholar] [CrossRef] [PubMed]
- Hui, L.; Leung, K.; Chen, H.M. The combined effects of antibacterial peptide cecropin A and anti-cancer agents on leukemia cells. Anticancer Res. 2002, 22, 2811–2816. [Google Scholar] [PubMed]
- Diaz-Achirica, P.; Ubach, J.; Guinea, A.; Andreu, D.; Rivas, L. The plasma membrane of Leishmania donovani promastigotes is the main target for CA(1-8)M(1-18), a synthetic cecropin A-melittin hybrid peptide. Biochem. J. 1998, 330, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, Y.; Furukawa, S.; Tanaka, H.; Yamakawa, M. Expression of antimicrobial peptide genes encoding Enbocin and Gloverin isoforms in the silkworm, Bombyx mori. Biosci. Biotechnol. Biochem. 2007, 71, 2233–2241. [Google Scholar] [CrossRef] [PubMed]
- Hemmi, H.; Ishibashi, J.; Hara, S.; Yamakawa, M. Solution structure of moricin, an antibacterial peptide, isolated from the silkworm Bombyx mori. FEBS Lett. 2002, 518, 33–38. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GO ID | Gene Ontology Term | GeneRatio | BgRatio | p Value | FDR |
|---|---|---|---|---|---|
| Molecular Function | |||||
| GO:0004553 | Hydrolase activity, hydrolyzing O-glycosyl compounds | 13/323 | 82/5764 | 0.56 × 10−3 | 0.04 |
| GO:0008146 | Sulfotransferase activity | 6/323 | 20/5764 | 0.59 × 10−3 | 0.04 |
| GO:0016782 | Transferase activity, transferring sulfur-containing groups | 6/323 | 21/5764 | 0.78 × 10−3 | 0.04 |
| GO:0016798 | Hydrolase activity, acting on glycosyl bonds | 13/323 | 85/5764 | 0.80 × 10−3 | 0.04 |
| GO:0022857 | Transmembrane transporter activity | 35/323 | 385/5764 | 0.27 × 10−2 | 0.11 |
| GO:0016705 | Oxidoreductase activity, acting on paired donors, with incorporation or reduction of molecular oxygen | 13/323 | 99/5764 | 0.33 × 10−2 | 0.11 |
| GO:0008061 | Chitin binding | 8/323 | 46/5764 | 0.36 × 10−2 | 0.11 |
| GO:0005215 | Transporter activity | 38/323 | 438/5764 | 0.40 × 10−2 | 0.11 |
| GO:0005506 | Iron ion binding | 12/323 | 101/5764 | 0.01 | 0.20 |
| GO:0004559 | α-mannosidase activity | 4/323 | 17/5764 | 0.01 | 0.20 |
| Cellular Component | |||||
| GO:0005576 | Extracellular region | 20/118 | 144/1987 | 0.19 × 10−3 | 0.01 |
| GO:0033178 | Proton-transporting two-sector ATPase complex, catalytic domain | 2/118 | 11/1987 | 0.14 | 1.00 |
| GO:0030133 | Transport vesicle | 1/118 | 10/1987 | 0.46 | 1.00 |
| GO:0030684 | Preribosome | 1/118 | 10/1987 | 0.46 | 1.00 |
| GO:0030054 | Cell junction | 1/118 | 11/1987 | 0.49 | 1.00 |
| GO:0098796 | Membrane protein complex | 5/118 | 78/1987 | 0.50 | 1.00 |
| GO:0016469 | Proton-transporting two-sector ATPase complex | 2/118 | 28/1987 | 0.50 | 1.00 |
| GO:0031012 | Extracellular matrix | 1/118 | 12/1987 | 0.52 | 1.00 |
| GO:0098797 | Plasma membrane protein complex | 1/118 | 12/1987 | 0.52 | 1.00 |
| GO:0005694 | Chromosome | 2/118 | 29/1987 | 0.52 | 1.00 |
| Biological Process | |||||
| GO:0009605 | Response to external stimulus | 9/231 | 10/3710 | 1.15 × 10−10 | 5.38 × 10−9 |
| GO:0009607 | Response to biotic stimulus | 9/231 | 10/3710 | 1.15 × 10−10 | 5.38 × 10−9 |
| GO:0009617 | Response to bacterium | 9/231 | 10/3710 | 1.15 × 10−10 | 5.38 × 10−9 |
| GO:0042742 | Defense response to bacterium | 9/231 | 10/3710 | 1.15 × 10−10 | 5.38 × 10−9 |
| GO:0043207 | Response to external biotic stimulus | 9/231 | 10/3710 | 1.15 × 10−10 | 5.38 × 10−9 |
| GO:0051707 | Response to other organism | 9/231 | 10/3710 | 1.15 × 10−10 | 5.38 × 10−9 |
| GO:0098542 | Defense response to other organism | 9/231 | 10/3710 | 1.15 × 10−10 | 5.38 × 10−9 |
| GO:0006952 | Defense response | 9/231 | 12/3710 | 2.26 × 10−9 | 9.26 × 10−8 |
| GO:0051704 | Multi-organism process | 9/231 | 14/3710 | 1.84 × 10−8 | 6.71 × 10−7 |
| GO:0002376 | Immune system process | 8/231 | 16/3710 | 1.67 × 10−6 | 4.98 × 10−5 |
| GO:0006955 | Immune response | 8/231 | 16/3710 | 1.67 × 10−6 | 4.98 × 10−5 |
| GO:0015074 | DNA integration | 10/231 | 47/3710 | 0.48 × 10−3 | 0.01 |
| GO:0006030 | Chitin metabolic process | 9/231 | 47/3710 | 0.20 × 10−2 | 0.04 |
| GO:0006040 | Amino sugar metabolic process | 9/231 | 47/3710 | 0.20 × 10−2 | 0.04 |
| GO:1901071 | Glucosamine-containing compound metabolic process | 9/231 | 47/3710 | 0.20 × 10−2 | 0.04 |
| Name | Gene ID | Dazao1 FPKM | Dazao2 FPKM | Dazao3 FPKM | sk1 FPKM | sk2 FPKM | sk3 FPKM | log2 FoldChange | FDR |
|---|---|---|---|---|---|---|---|---|---|
| Chitin binding and chitin metabolic | |||||||||
| Chitin binding peritrophin A domain | novel.3284 | 21.23 | 53.76 | 14.25 | 7.81 | 0 | 4.59 | −2.81 | 0.02 |
| Chitin binding peritrophin A domain | novel.4995 | 2197.11 | 810.02 | 2889.63 | 109.39 | 172.40 | 303.74 | −3.33 | 3.38 × 10−9 |
| Chitin binding peritrophin A domain | novel.6300 | 36.52 | 75.27 | 58.77 | 17.58 | 19.42 | 8.02 | −1.92 | 0.40 × 10−2 |
| Chitin binding peritrophin A domain | novel.8828 | 709.16 | 957.87 | 943.91 | 409.22 | 526.90 | 559.34 | −0.80 | 0.02 |
| Chitin binding peritrophin A domain | novel.311 | 0 | 0.90 | 4.45 | 16.60 | 20.64 | 12.61 | 3.23 | 0.02 |
| Chitin binding peritrophin A domain | novel.5689 | 427.20 | 414.87 | 353.52 | 774.50 | 807.35 | 798.89 | 1.00 | 3.32 × 10−5 |
| Chitin binding peritrophin A domain | novel.6971 | 907.04 | 1027.76 | 1040.09 | 1899.62 | 3166.25 | 1856.81 | 1.22 | 0.51 × 10−3 |
| BmCPAP1-D | BGIBMGA006382 | 27.18 | 52.87 | 26.71 | 362.34 | 350.86 | 679.69 | 3.71 | 1.53 × 10−16 |
| Chitinase | BGIBMGA008709 | 35.67 | 29.57 | 35.62 | 21.49 | 173.61 | 209.75 | 2.00 | 0.03 |
| Hydrolase | |||||||||
| Glycosyl hydrolase family 1 | novel.2031 | 13.59 | 10.75 | 16.92 | 3.91 | 1.21 | 1.15 | −2.67 | 0.04 |
| Glycosyl hydrolase family 1 | novel.12594 | 107.01 | 114.69 | 73.02 | 48.83 | 32.78 | 33.24 | −1.35 | 0.48 × 10−2 |
| Glycosyl hydrolase family 1 | novel.12616 | 124.00 | 19.71 | 1.78 | 0 | 0 | 0 | −7.90 | 0.02 |
| Glycosyl hydrolase family 31 | BGIBMGA013995 | 86.63 | 116.49 | 103.30 | 51.76 | 40.06 | 46.99 | −1.14 | 0.70 × 10−2 |
| Glycosyl hydrolases family 38 C-terminal domain | novel.1424 | 189.39 | 150.54 | 199.47 | 55.67 | 74.06 | 30.95 | −1.75 | 9.54 × 10−5 |
| Glycosyl hydrolases family 38 N-terminal domain | novel.15227 | 9.34 | 5.38 | 8.01 | 0 | 0 | 0 | −5.22 | 0.02 |
| α-1,2-mannosidase | BGIBMGA002426 | 879.01 | 478.49 | 1114.00 | 144.55 | 122.62 | 338.12 | −2.03 | 0.29 × 10−3 |
| Mannosyl-oligosaccharide α-1,2-mannosidase IA | BGIBMGA002486 | 62.85 | 52.87 | 96.17 | 21.49 | 7.28 | 25.22 | −1.96 | 0.29 × 10−2 |
| Trehalase 1B | BGIBMGA005665 | 84.93 | 63.62 | 60.55 | 24.42 | 30.35 | 25.22 | −1.39 | 0.42 × 10−2 |
| β-glucosidase | BGIBMGA010811 | 0 | 0 | 0 | 4.88 | 9.71 | 6.88 | 5.46 | 0.01 |
| Destabilase | novel.6630 | 16.14 | 16.13 | 20.48 | 82.04 | 110.48 | 61.89 | 2.27 | 1.97 × 10−6 |
| α galactosidase A | novel.8493 | 255.64 | 443.54 | 332.15 | 1616.39 | 1016.16 | 1565.68 | 2.03 | 8.92 × 10−9 |
| Transferase | |||||||||
| Sulfotransferase family | novel.1630 | 327.83 | 408.60 | 273.38 | 614.32 | 675.01 | 706.05 | 0.98 | 0.14 × 10−2 |
| Sulfotransferase family | novel.2592 | 545.24 | 826.15 | 708.83 | 197.29 | 154.18 | 205.17 | −1.90 | 3.15 × 10−10 |
| Sulfotransferase domain | novel.6336 | 81.53 | 98.56 | 138.03 | 40.04 | 48.56 | 43.55 | −1.27 | 0.59 × 10−2 |
| Sulfotransferase domain | novel.15388 | 177.50 | 225.80 | 206.59 | 0.98 | 2.43 | 0 | −7.52 | 4.53 × 10−15 |
| Heparan-sulfate 6-O-sulfotransferase 2 | BGIBMGA007552 | 32.27 | 17.92 | 36.51 | 5.86 | 6.07 | 12.61 | −1.83 | 0.04 |
| Amine sulfotransferase | BGIBMGA010842 | 2.55 | 2.69 | 2.67 | 13.67 | 34.00 | 8.02 | 2.81 | 0.02 |
| Antimicrobial peptides | |||||||||
| Cecropin A | BGIBMGA006280 | 454.37 | 282.25 | 518.26 | 12.70 | 26.71 | 30.95 | −4.17 | 3.68 × 10−21 |
| Cecropin A | BGIBMGA014285 | 241.20 | 233.87 | 281.39 | 3.91 | 23.07 | 0 | −4.82 | 0.34 × 10−2 |
| Cecropin B | BGIBMGA000023 | 163.06 | 60.93 | 153.16 | 0 | 0 | 0 | −9.27 | 4.63 × 10−11 |
| Cecropin B | BGIBMGA000036 | 172.41 | 38.53 | 126.45 | 2.93 | 10.93 | 24.07 | −3.16 | 0.28 × 10−3 |
| Cecropin CBM2 | BGIBMGA000021 | 44.16 | 8.06 | 35.62 | 0 | 0 | 0 | −7.17 | 1.98 × 10−5 |
| Cecropin-D | BGIBMGA000017 | 287.06 | 339.60 | 274.27 | 45.90 | 98.34 | 191.41 | −1.43 | 0.03 |
| Enbocin2 | BGIBMGA000039 | 82.38 | 37.63 | 75.69 | 2.93 | 8.50 | 12.61 | −3.05 | 1.74 × 10−5 |
| Enbocin3 | BGIBMGA000018 | 9.34 | 14.34 | 8.90 | 0 | 0 | 2.29 | −3.87 | 0.04 |
| Moricin-1 | BGIBMGA011495 | 95.97 | 87.81 | 58.77 | 29.30 | 18.21 | 24.07 | −1.75 | 0.45 × 10−3 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, D.; Hu, H.; Tong, X.; Han, M.; Wu, S.; Ding, X.; Dai, F.; Lu, C. Comparative Analysis of the Integument Transcriptomes between stick Mutant and Wild-Type Silkworms. Int. J. Mol. Sci. 2018, 19, 3158. https://doi.org/10.3390/ijms19103158
Tan D, Hu H, Tong X, Han M, Wu S, Ding X, Dai F, Lu C. Comparative Analysis of the Integument Transcriptomes between stick Mutant and Wild-Type Silkworms. International Journal of Molecular Sciences. 2018; 19(10):3158. https://doi.org/10.3390/ijms19103158
Chicago/Turabian StyleTan, Duan, Hai Hu, Xiaoling Tong, Minjin Han, Songyuan Wu, Xin Ding, Fangyin Dai, and Cheng Lu. 2018. "Comparative Analysis of the Integument Transcriptomes between stick Mutant and Wild-Type Silkworms" International Journal of Molecular Sciences 19, no. 10: 3158. https://doi.org/10.3390/ijms19103158