Enhanced Pulsatile Growth Hormone Secretion and Altered Metabolic Hormones by in Vivo Hexarelin Treatment in Streptozotocin-Induced Diabetic Rats



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
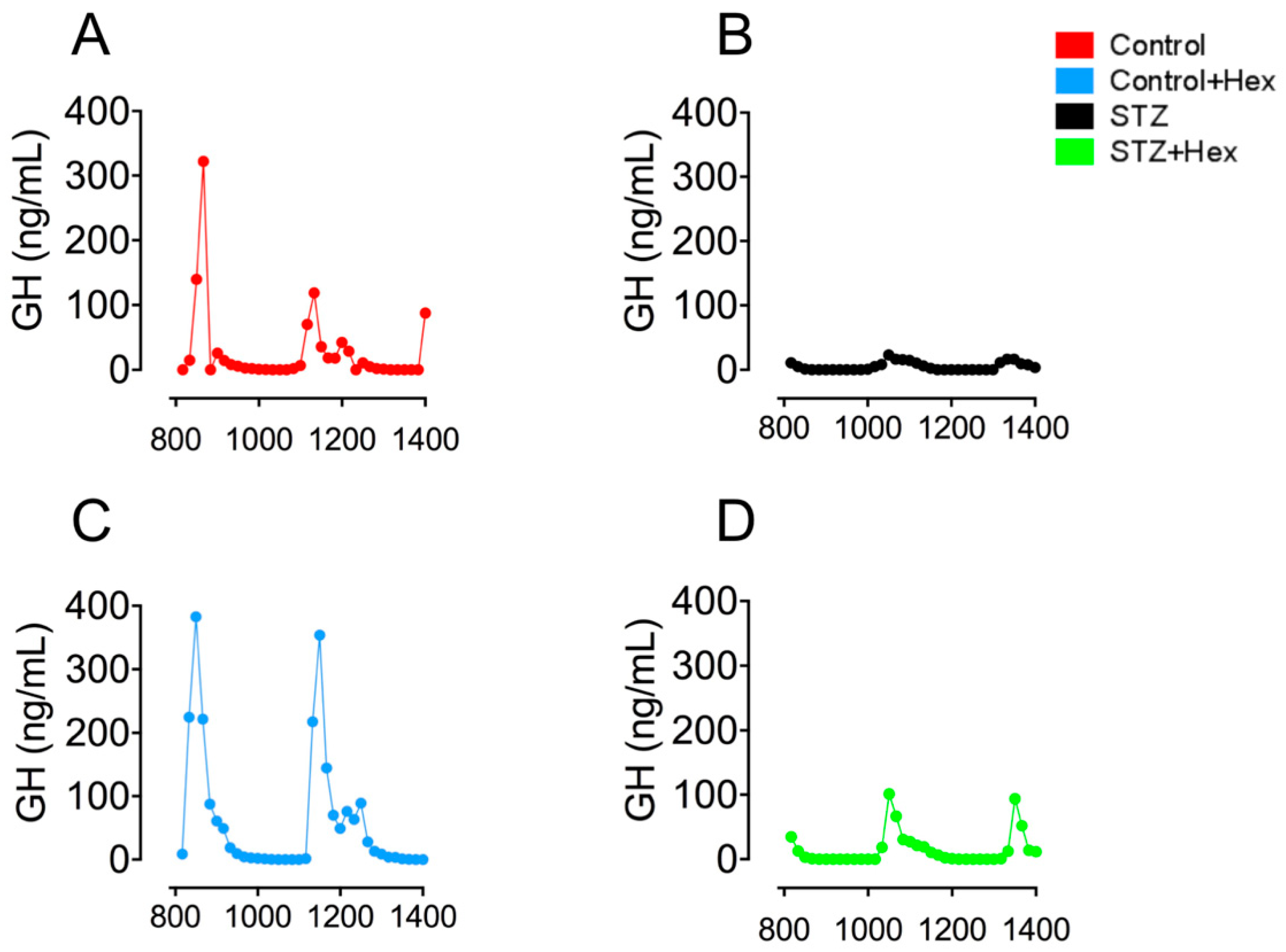
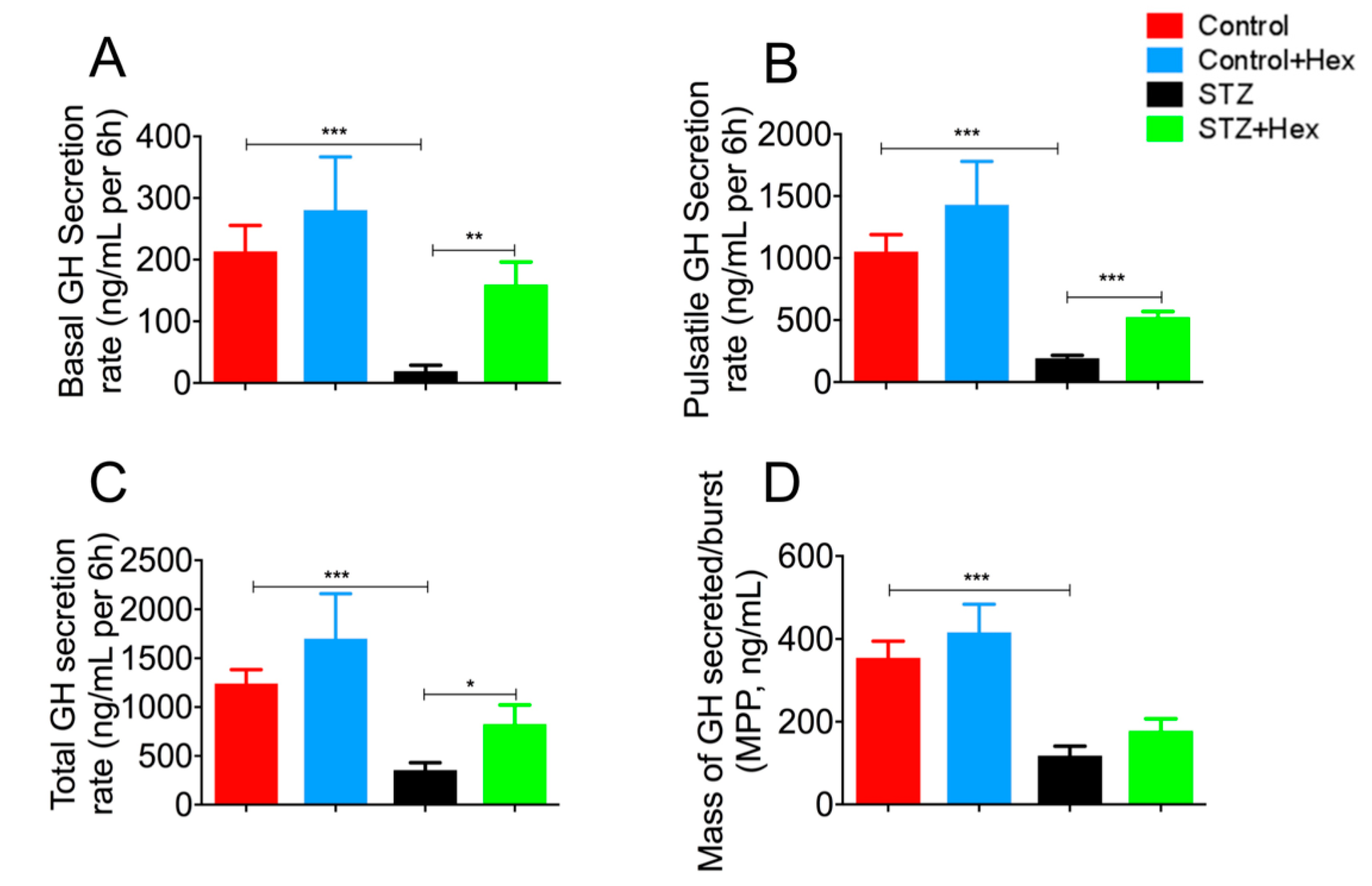
2.1. Pulsatile GH Secretion in STZ- and Hex-Treated Rats
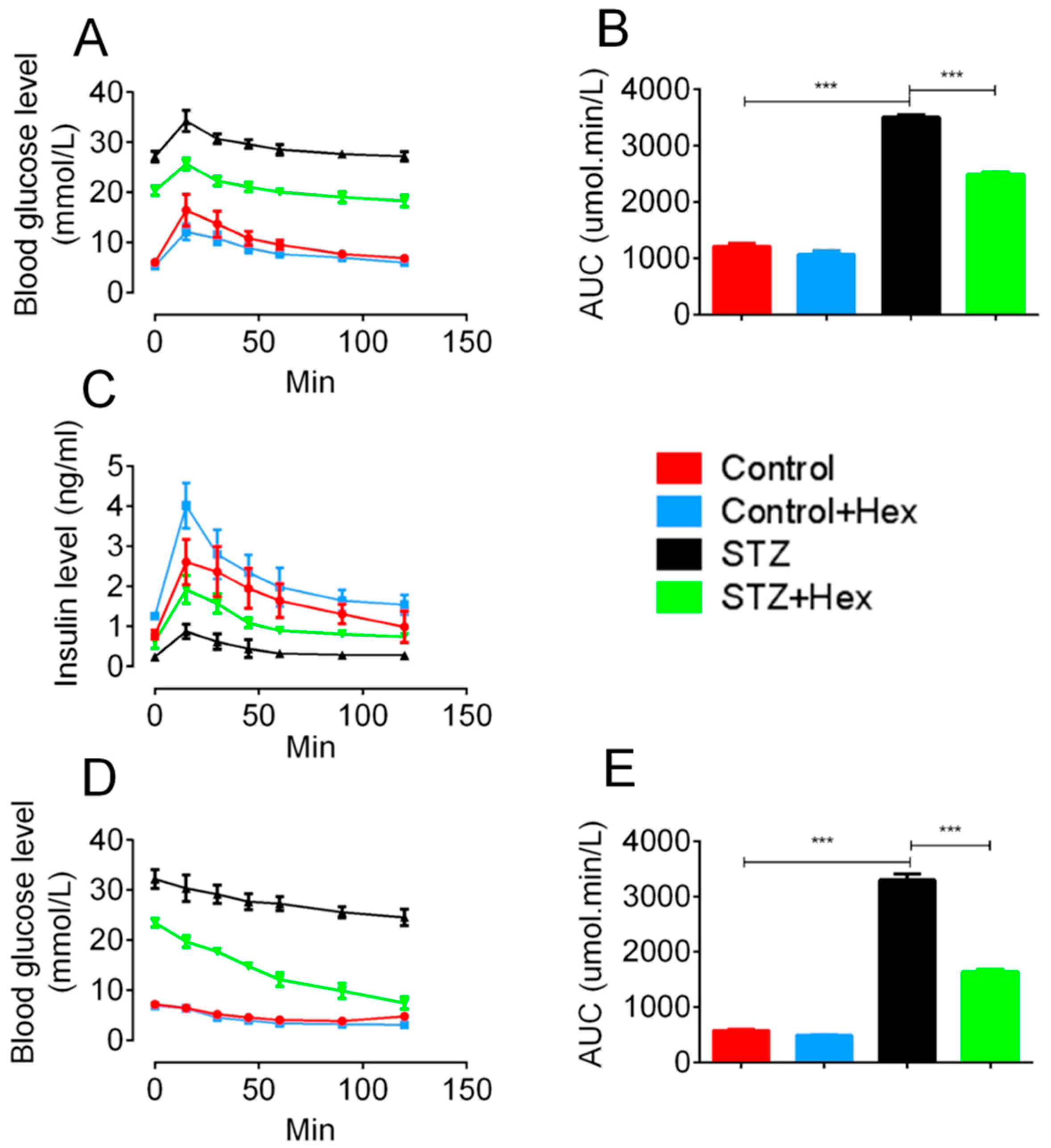
2.2. Glucose Tolerance Test (GTT) and Insulin Tolerance Test (ITT) in STZ- and Hex-Treated Rats
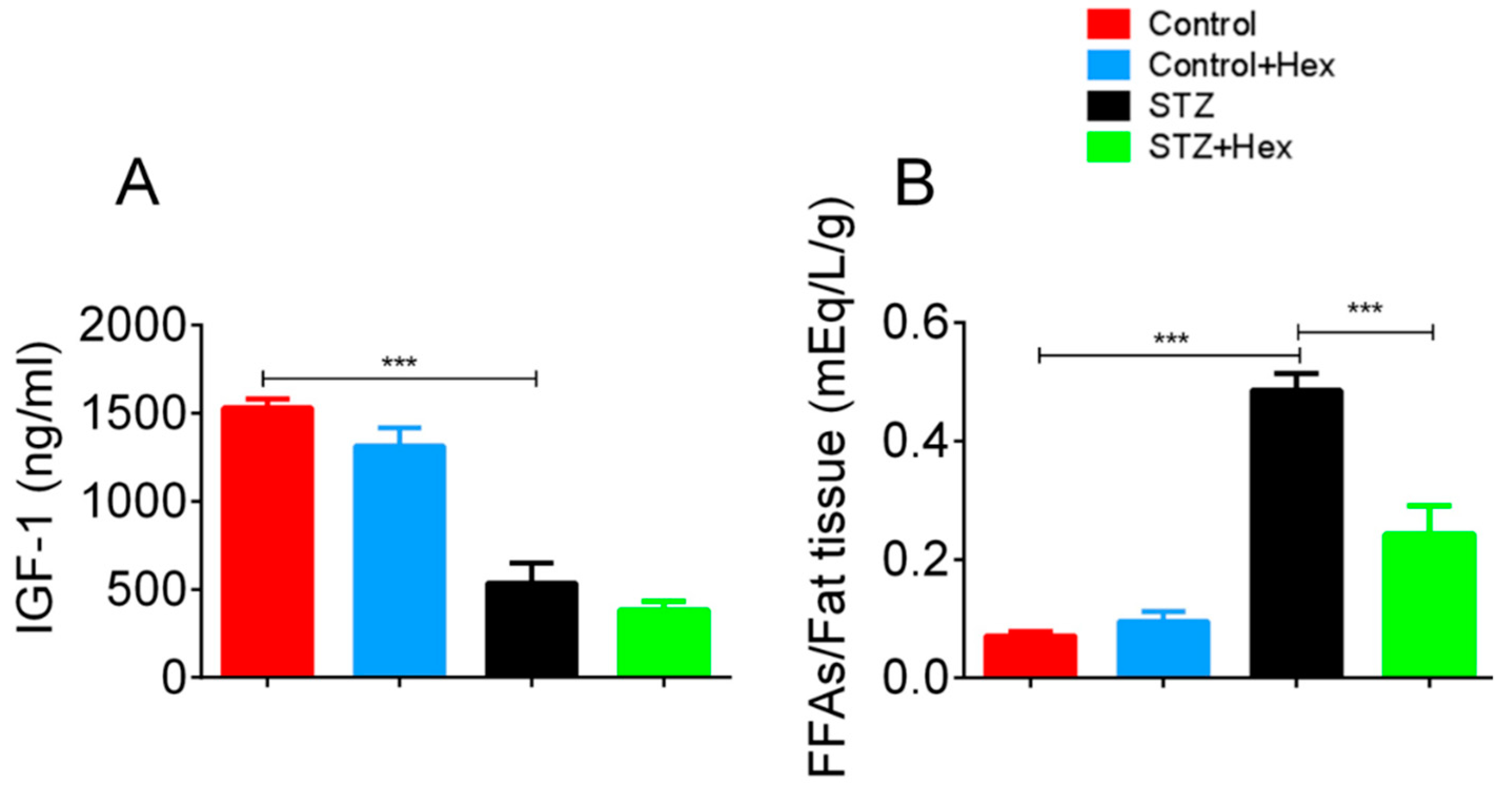
2.3. Circulating Levels of IGF-1 and FFAs in STZ- and Hex-Treated Rats
3. Discussion
4. Materials and Methods
4.1. Diabetic Rat Model
4.2. Blood Collection
4.3. Analysis of Pulsatile Growth Hormone Secretion by Sensitive Sandwich ELISA
4.4. Deconvolution Analysis
4.5. Glucose Tolerance Test (GTT) and Insulin Tolerance Test (ITT)
4.6. Euthanization
4.7. Glucose and Hormone Analysis
4.8. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Giustina, A.; Veldhuis, J.D. Pathophysiology of the neuroregulation of growth hormone secretion in experimental animals and the human. Endocr. Rev. 1998, 19, 717–797. [Google Scholar] [CrossRef] [PubMed]
- Olchovsky, D.; Bruno, J.F.; Wood, T.L.; Gelato, M.C.; Leidy, J.W., Jr.; Gilbert, J.M., Jr.; Berelowitz, M. Altered pituitary growth hormone (GH) regulation in streptozotocin-diabetic rats: A combined defect of hypothalamic somatostatin and GH-releasing factor. Endocrinology 1990, 126, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Underwood, L.E.; D’Ercole, A.J.; Clemmons, D.R.; Van Wyk, J.J. Paracrine functions of somatomedins. Clin. Endocrinol. Metab. 1986, 15, 59–77. [Google Scholar] [CrossRef]
- Roelfsema, V.; Clark, R.G. The growth hormone and insulin-like growth factor axis: Its manipulation for the benefit of growth disorders in renal failure. J. Am. Soc. Nephrol. JASN 2001, 12, 1297–1306. [Google Scholar] [PubMed]
- Davidson, M.B. Effect of growth hormone on carbohydrate and lipid metabolism. Endocr. Rev. 1987, 8, 115–131. [Google Scholar] [CrossRef] [PubMed]
- Randle, P.J.; Garland, P.B.; Hales, C.N.; Newsholme, E.A. The glucose fatty-acid cycle. Its role in insulin sensitivity and the metabolic disturbances of diabetes mellitus. Lancet 1963, 1, 785–789. [Google Scholar] [CrossRef]
- Ndon, J.A.; Giustina, A.; Wehrenberg, W.B. Hypothalamic regulation of impaired growth hormone secretion in diabetic rats. Neuroendocrinology 1992, 55, 500–505. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Caro, J.; Gonzalez, C.; Jolin, T. Diurnal variations of plasma growth hormone, thyrotropin, thyroxine, and triiodothyronine in streptozotocin-diabetic and food-restricted rats. Endocrinology 1984, 115, 2227–2232. [Google Scholar] [CrossRef] [PubMed]
- Tannenbaum, G.S. Growth hormone secretory dynamics in streptozotocin diabetes: Evidence of a role for endogenous circulating somatostatin. Endocrinology 1981, 108, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Bluet-Pajot, M.T.; Durand, D.; Kordon, C. Influence of streptozotocin-induced diabetes on growth hormone secretion in the rat. Neuroendocrinology 1983, 36, 307–309. [Google Scholar] [CrossRef] [PubMed]
- Locatelli, V.; Miyoshi, H.; Bestetti, G.; Rossi, G.L.; Muller, E.E. Effect of growth hormone-releasing stimuli in streptozotocin diabetic rats. Brain Res. 1985, 341, 35–40. [Google Scholar] [CrossRef]
- Murao, K.; Sato, M.; Mizobuchi, M.; Nimi, M.; Ishida, T.; Takahara, J. Acute effects of hypoglycemia and hyperglycemia on hypothalamic growth hormone-releasing hormone and somatostatin gene expression in the rat. Endocrinology 1994, 134, 418–423. [Google Scholar] [CrossRef] [PubMed]
- Nieves-Rivera, F.; Kerrigan, J.R.; Krieg, R.J., Jr.; Egan, J.; Hwang, L.J.; Truumees, E.; Veldhuis, J.D.; Evans, W.S.; Rogol, A.D. Altered growth hormone (GH) secretion in vivo and in vitro in the diabetes-prone bb/worcester rat. Growth Regul. 1993, 3, 235–244. [Google Scholar] [PubMed]
- Baes, M.; Vale, W.W. Characterization of the glucose-dependent release of growth hormone-releasing factor and somatostatin from superfused rat hypothalami. Neuroendocrinology 1990, 51, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Welsh, J.B.; Szabo, M. Impaired suppression of growth hormone release by somatostatin in cultured adenohypophyseal cells of spontaneously diabetic bb/w rats. Endocrinology 1988, 123, 2230–2234. [Google Scholar] [CrossRef] [PubMed]
- Baxter, R.C.; Brown, A.S.; Turtle, J.R. Association between serum insulin, serum somatomedin and liver receptors for human growth hormone in streptozotocin diabetes. Horm. Metab. Res. 1980, 12, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Bhathena, S.J.; Voyles, N.R.; Smith, S.; Recant, L. Decreased glucagon receptors in diabetic rat hepatocytes. Evidence for regulation of glucagon receptors by hyperglucagonemia. J. Clin. Investig. 1978, 61, 1488–1497. [Google Scholar] [CrossRef] [PubMed]
- Rahim, A.; O’Neill, P.A.; Shalet, S.M. Growth hormone status during long-term hexarelin therapy. J. Clin. Endocrinol. Metab. 1998, 83, 1644–1649. [Google Scholar] [CrossRef] [PubMed]
- Hataya, Y.; Akamizu, T.; Takaya, K.; Kanamoto, N.; Ariyasu, H.; Saijo, M.; Moriyama, K.; Shimatsu, A.; Kojima, M.; Kangawa, K.; et al. A low dose of ghrelin stimulates growth hormone (GH) release synergistically with GH-releasing hormone in humans. J. Clin. Endocrinol. Metab. 2001, 86, 4552. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.G.; Carlsson, L.M.; Trojnar, J.; Robinson, I.C. The effects of a growth hormone-releasing peptide and growth hormone-releasing factor in conscious and anaesthetized rats. J. Neuroendocrinol. 1989, 1, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Pandya, N.; DeMott-Friberg, R.; Bowers, C.Y.; Barkan, A.L.; Jaffe, C.A. Growth hormone (GH)-releasing peptide-6 requires endogenous hypothalamic GH-releasing hormone for maximal GH stimulation. J. Clin. Endocrinol. Metab. 1998, 83, 1186–1189. [Google Scholar] [CrossRef] [PubMed]
- Maheshwari, H.G.; Rahim, A.; Shalet, S.M.; Baumann, G. Selective lack of growth hormone (GH) response to the GH-releasing peptide hexarelin in patients with gh-releasing hormone receptor deficiency. J. Clin. Endocrinol. Metab. 1999, 84, 956–959. [Google Scholar] [PubMed]
- Gondo, R.G.; Aguiar-Oliveira, M.H.; Hayashida, C.Y.; Toledo, S.P.; Abelin, N.; Levine, M.A.; Bowers, C.Y.; Souza, A.H.; Pereira, R.M.; Santos, N.L.; et al. Growth hormone-releasing peptide-2 stimulates GH secretion in GH-deficient patients with mutated GH-releasing hormone receptor. J. Clin. Endocrinol. Metab. 2001, 86, 3279–3283. [Google Scholar] [CrossRef] [PubMed]
- Guillaume, V.; Magnan, E.; Cataldi, M.; Dutour, A.; Sauze, N.; Renard, M.; Razafindraibe, H.; Conte-Devolx, B.; Deghenghi, R.; Lenaerts, V.; et al. Growth hormone (GH)-releasing hormone secretion is stimulated by a new GH-releasing hexapeptide in sheep. Endocrinology 1994, 135, 1073–1076. [Google Scholar] [CrossRef] [PubMed]
- Tannenbaum, G.S.; Lapointe, M.; Beaudet, A.; Howard, A.D. Expression of growth hormone secretagogue-receptors by growth hormone-releasing hormone neurons in the mediobasal hypothalamus. Endocrinology 1998, 139, 4420–4423. [Google Scholar] [CrossRef] [PubMed]
- Dickson, S.L.; Luckman, S.M. Induction of c-fos messenger ribonucleic acid in neuropeptide y and growth hormone (GH)-releasing factor neurons in the rat arcuate nucleus following systemic injection of the GH secretagogue, GH-releasing peptide-6. Endocrinology 1997, 138, 771–777. [Google Scholar] [CrossRef] [PubMed]
- Hewson, A.K.; Dickson, S.L. Systemic administration of ghrelin induces fos and EGR-1 proteins in the hypothalamic arcuate nucleus of fasted and fed rats. J. Neuroendocrinol. 2000, 12, 1047–1049. [Google Scholar] [CrossRef] [PubMed]
- Drisko, J.E.; Faidley, T.D.; Zhang, D.; McDonald, T.J.; Nicolich, S.; Hora, D.F.; Cunningham, P.; Li, C.; Rickes, E.; McNamara, L.; et al. Administration of a nonpeptidyl growth hormone secretagogue, L-163, 255, changes somatostatin pattern, but has no effect on patterns of growth hormone-releasing factor in the hypophyseal-portal circulation of the conscious pig. Proc. Soc. Exp. Biol. Med. (N. Y.) 1999, 222, 70–77. [Google Scholar] [CrossRef]
- Cohen, R.M.; Abplanalp, W.A. Resistance of growth hormone secretion to somatostatin in men with type I diabetes mellitus. Diabetes 1991, 40, 1251–1258. [Google Scholar] [CrossRef] [PubMed]
- Ismail, I.S.; Scanlon, M.F.; Peters, J.R. Cholinergic control of growth hormone (GH) responses to GH-releasing hormone in insulin dependent diabetics: Evidence for attenuated hypothalamic somatostatinergic tone and decreased GH autofeedback. Clin. Endocrinol. 1993, 38, 149–157. [Google Scholar] [CrossRef]
- Serri, O.; Brazeau, P. Growth hormone responsiveness in vivo and in vitro to growth hormone releasing factor in the spontaneously diabetic bb wistar rat. Neuroendocrinology 1987, 46, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Busiguina, S.; Argente, J.; Garcia-Segura, L.M.; Chowen, J.A. Anatomically specific changes in the expression of somatostatin, growth hormone-releasing hormone and growth hormone receptor mRNA in diabetic rats. J. Neuroendocrinol. 2000, 12, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Joanny, P.; Peyre, G.; Steinberg, J.; Guillaume, V.; Pesce, G.; Becquet, D.; Oliver, C. Effect of diabetes on in vivo and in vitro hypothalamic somatostatin release. Neuroendocrinology 1992, 55, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Sohn, S.; Lee, M.; Jung, J.; Kineman, R.D.; Park, S. Differential responses of the growth hormone axis in two rat models of streptozotocin-induced insulinopenic diabetes. J. Endocrinol. 2006, 188, 263–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diz Chaves, Y.; Spuch Calvar, C.; Perez Tilve, D.; Mallo Ferrer, F. GH responses to GHRH and GHRP-6 in streptozotocin (STZ)-diabetic rats. Life Sci. 2003, 73, 3375–3385. [Google Scholar] [CrossRef] [PubMed]
- Tannenbaum, G.S.; Bowers, C.Y. Interactions of growth hormone secretagogues and growth hormone-releasing hormone/somatostatin. Endocrine 2001, 14, 21–27. [Google Scholar] [CrossRef]
- Diz-Chaves, Y.; Spuch, C.; Perez, D.; Mallo, F. Ghrelin improves growth hormone responses to growth hormone-releasing hormone in a streptozotocin-diabetic model of delayed onset. J. Endocrinol. Investig. 2007, 30, 298–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansen, P.B.; Segev, Y.; Landau, D.; Phillip, M.; Flyvbjerg, A. Growth hormone (GH) hypersecretion and GH receptor resistance in streptozotocin diabetic mice in response to a GH secretagogue. Exp. Diabesity Res. 2003, 4, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Ardestani, A.; Paroni, F.; Azizi, Z.; Kaur, S.; Khobragade, V.; Yuan, T.; Frogne, T.; Tao, W.; Oberholzer, J.; Pattou, F.; et al. MST1 is a key regulator of beta cell apoptosis and dysfunction in diabetes. Nat. Med. 2014, 20, 385–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Zhang, X.; Chen, J.; Lin, C.; Shao, R.; Yan, C.; Chen, C. Hexarelin protects rodent pancreatic beta-cells function from cytotoxic effects of streptozotocin involving mitochondrial signalling pathways in vivo and in vitro. PLoS ONE 2016, 11, e0149730. [Google Scholar]
- Lehmann, J.M.; Moore, L.B.; Smith-Oliver, T.A.; Wilkison, W.O.; Willson, T.M.; Kliewer, S.A. An antidiabetic thiazolidinedione is a high affinity ligand for peroxisome proliferator-activated receptor gamma (PPAR γ). J. Biol. Chem. 1995, 270, 12953–12956. [Google Scholar] [CrossRef] [PubMed]
- Lambe, K.G.; Tugwood, J.D. A human peroxisome-proliferator-activated receptor-γ is activated by inducers of adipogenesis, including thiazolidinedione drugs. Eur. J. Biochem. FEBS 1996, 239, 1–7. [Google Scholar] [CrossRef]
- Yki-Jarvinen, H. Thiazolidinediones. N. Engl. J. Med. 2004, 351, 1106–1118. [Google Scholar] [CrossRef] [PubMed]
- Avallone, R.; Demers, A.; Rodrigue-Way, A.; Bujold, K.; Harb, D.; Anghel, S.; Wahli, W.; Marleau, S.; Ong, H.; Tremblay, A. A growth hormone-releasing peptide that binds scavenger receptor CD36 and ghrelin receptor up-regulates sterol transporters and cholesterol efflux in macrophages through a peroxisome proliferator-activated receptor gamma-dependent pathway. Mol. Endocrinol. 2006, 20, 3165–3178. [Google Scholar] [CrossRef] [PubMed]
- Masaoka, T.; Suzuki, H.; Hosoda, H.; Ota, T.; Minegishi, Y.; Nagata, H.; Kangawa, K.; Ishii, H. Enhanced plasma ghrelin levels in rats with streptozotocin-induced diabetes. FEBS Lett. 2003, 541, 64–68. [Google Scholar] [CrossRef] [Green Version]
- Teppala, S.; Shankar, A. Association between serum IGF-1 and diabetes among U.S. Adults. Diabetes Care 2010, 33, 2257–2259. [Google Scholar] [CrossRef] [PubMed]
- Argente, J.; Garcia-Segura, L.M.; Pozo, J.; Chowen, J.A. Growth hormone-releasing peptides: Clinical and basic aspects. Horm. Res. 1996, 46, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Colao, A.; Barkan, A.L.; Scarpa, R. Growth hormone/insulin-like growth factor-i system and connective tissues: Basic aspects and clinical implications. Rheumatic Dis. Clin. N. Am. 2005, 31, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Nagaya, N.; Uematsu, M.; Kojima, M.; Ikeda, Y.; Yoshihara, F.; Shimizu, W.; Hosoda, H.; Hirota, Y.; Ishida, H.; Mori, H.; et al. Chronic administration of ghrelin improves left ventricular dysfunction and attenuates development of cardiac cachexia in rats with heart failure. Circulation 2001, 104, 1430–1435. [Google Scholar] [CrossRef] [PubMed]
- Dang, A.Q.; Faas, F.H.; Jethmalani, S.M.; Carter, W.J. Decreased incorporation of long-chain fatty acids into erythrocyte phospholipids of STZ-D rats. Diabetes 1991, 40, 1645–1651. [Google Scholar] [CrossRef] [PubMed]
- Muller, D.; Nitsch, R.M.; Wurtman, R.J.; Hoyer, S. Streptozotocin increases free fatty acids and decreases phospholipids in rat brain. J. Neural Transm. 1998, 105, 1271–1281. [Google Scholar] [CrossRef] [PubMed]
- Scheuermann-Freestone, M.; Madsen, P.L.; Manners, D.; Blamire, A.M.; Buckingham, R.E.; Styles, P.; Radda, G.K.; Neubauer, S.; Clarke, K. Abnormal cardiac and skeletal muscle energy metabolism in patients with type 2 diabetes. Circulation 2003, 107, 3040–3046. [Google Scholar] [CrossRef] [PubMed]
- DeFronzo, R.A.; Gunnarsson, R.; Bjorkman, O.; Olsson, M.; Wahren, J. Effects of insulin on peripheral and splanchnic glucose metabolism in noninsulin-dependent (type II) diabetes mellitus. J. Clin. Investig. 1985, 76, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Reaven, G.M.; Chang, H.; Ho, H.; Jeng, C.Y.; Hoffman, B.B. Lowering of plasma glucose in diabetic rats by antilipolytic agents. Am. J. Physiol. 1988, 254, E23–E30. [Google Scholar] [CrossRef] [PubMed]
- Flarsheim, C.E.; Grupp, I.L.; Matlib, M.A. Mitochondrial dysfunction accompanies diastolic dysfunction in diabetic rat heart. Am. J. Physiol.-Heart Circul. Physiol. 1996, 271, H192–H202. [Google Scholar] [CrossRef] [PubMed]
- Steyn, F.J.; Huang, L.; Ngo, S.T.; Leong, J.W.; Tan, H.Y.; Xie, T.Y.; Parlow, A.F.; Veldhuis, J.D.; Waters, M.J.; Chen, C. Development of a method for the determination of pulsatile growth hormone secretion in mice. Endocrinology 2011, 152, 3165–3171. [Google Scholar] [CrossRef] [PubMed]
- Keenan, D.M.; Chattopadhyay, S.; Veldhuis, J.D. Composite model of time-varying appearance and disappearance of neurohormone pulse signals in blood. J. Theor. Biol. 2005, 236, 242–255. [Google Scholar] [CrossRef] [PubMed]
- Keenan, D.M.; Iranmanesh, A.; Veldhuis, J.D. Analytical construct of reversible desensitization of pituitary-testicular signaling: Illustrative application in aging. Am. J. Physiol. Regul. Integr. Compar. Physiol. 2011, 300, R349–R360. [Google Scholar] [CrossRef] [PubMed]
- Veldhuis, J.D.; Johnson, M.L.; Veldhuis, O.L.; Straume, M.; Pincus, S.M. Impact of pulsatility on the ensemble orderliness (approximate entropy) of neurohormone secretion. Am. J. Physiol. Regul. Integr. Compar. Physiol. 2001, 281, R1975–R1985. [Google Scholar] [CrossRef] [PubMed]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Yang, J.-K.; Chen, C. Enhanced Pulsatile Growth Hormone Secretion and Altered Metabolic Hormones by in Vivo Hexarelin Treatment in Streptozotocin-Induced Diabetic Rats. Int. J. Mol. Sci. 2018, 19, 3067. https://doi.org/10.3390/ijms19103067
Zhang X, Yang J-K, Chen C. Enhanced Pulsatile Growth Hormone Secretion and Altered Metabolic Hormones by in Vivo Hexarelin Treatment in Streptozotocin-Induced Diabetic Rats. International Journal of Molecular Sciences. 2018; 19(10):3067. https://doi.org/10.3390/ijms19103067
Chicago/Turabian StyleZhang, Xinli, Jin-Kui Yang, and Chen Chen. 2018. "Enhanced Pulsatile Growth Hormone Secretion and Altered Metabolic Hormones by in Vivo Hexarelin Treatment in Streptozotocin-Induced Diabetic Rats" International Journal of Molecular Sciences 19, no. 10: 3067. https://doi.org/10.3390/ijms19103067