Abstract
Bacterial blight (BB) and bacterial leaf streak (BLS), caused by Xanthomonas oryzae pv. oryzae and Xanthomonas oryzae pv. oryzicola, respectively, are two devastating diseases in rice planting areas worldwide. It has been proven that adoption of rice resistance is the most effective, economic, and environment-friendly strategy to avoid yield loss caused by BB and BLS. As a model system for plant—pathogen interaction, the rice—X. oryzae pathosystem has been intensively investigated in the past decade. Abundant studies have shown that the resistance and susceptibility of rice to X. oryzae is determined by molecular interactions between rice genes or their products and various pathogen effectors. In this review, we briefly overviewed the literature regarding the diverse interactions, focusing on recent advances in uncovering mechanisms of rice resistance and X. oryzae virulence. Our analysis and discussions will not only be helpful for getting a better understanding of coevolution of the rice innate immunity and X. oryzae virulence, but it will also provide new insights for application of plant R genes in crop breeding.
1. Introduction
Phytopathogenic bacteria of the Xanthomonas genus are causative agents of numerous destructive diseases in more than 300 plant species, including some important crops like rice, citrus, and cotton [1,2]. X. oryzae pv. oryzae (Xoo) and X. oryzae pv. oryzicola (Xoc), two close pathovars of X. oryzae species, cause bacterial blight (BB) and bacterial leaf streak (BLS), respectively, in rice. These are among the major diseases in most rice planting areas worldwide, leading to reduced yield and poor quality of the affected plants [3]. Unlike nonpathogenic bacteria, Xanthomonas species attack plants by mainly relying on various effectors secreted through different types of protein secretion systems, particularly the type III secretion system (T3S), to suppress host immunity and obtain nutrients from plants. In X. oryzae, T3S effectors (T3SEs) are grouped into two types: transcription activator-like effectors (TALEs) and non-TALEs (also called Xanthomonas outer proteins, Xops) [2,4].
TALEs, formerly cited as AvrBs3/PthA proteins, are important virulence factors of most species in the Xanthomonas genus. In the past few years, tremendous research progresses have been achieved in TALE biology, particularly on TALE’s virulence contribution [5,6,7,8]. The TALE proteins are injected into plant cell, then enter the nucleus, playing roles as transcription factors by activating expression of target genes to promote susceptibility or suppress effector-triggered immunity (ETI) of plants [1,9,10]. For example, PthA from X. citri controls pustule formation and bacterial population density in plant leaf during citrus bacterial canker by activating CsLOB1, a member of the Lateral Organ Boundaries gene family [11]. Similarly, AvrBs3 from X. campestris pv. vesicatoria causes hypertrophy and facilitate bacterial spreading in pepper by activating the bHLH transcription factor UPA20 [12]. Major virulence TALEs (e.g., PthXo1 and AvrXa7) from Xoo activates the expression of multiple rice SWEET genes (e.g., OsSWEET11 and OsSWEET14) to promote susceptibility [13,14,15], and Tal7 from Xoc activates the expression of Os09g29100 to suppress avrXa7-induced resistance in rice [16]. Both Xoo and Xoc harbor large TALE repertoires, usually exceeding 8 in Xoo isolates and 20 in Xoc isolates, are significantly different from other pathovars in Xanthomonas [17,18,19].
Xops of Xanthomonas constitute another important group of virulence factors, which is mainly involved in manipulation of signaling in plant defense response. Several dozen Xops have been intensively studied. XopYMAFF311018 from the Xoo strain T7174 interacts with two receptor-like cytoplasmic kinases (RLCKs)—OsRLCK55 and OsRLCK185—and suppresses OsRLCK185 phosphorylation to interfere mitogen-activated protein kinase (MAPK) signaling [20]. XopPMAFF311018 targets ubiquitin E3 ligase, OsPUB44, and reduces ligase activity to suppress the PAMPs-triggered immunity (PTI) in rice [21]. It has also been shown that XopNKXO85, an important effector of Xoo strain KXO85, targets OsVOZ2 and OsXNP and utilizes OsVOZ2 for successful infection and increased virulence [22]. Additionally, XopZPXO99, XopFITCCBB0002, XopRMAFF311018/1375, and XopQBXO43 have all been reported to affect plant defense and significantly contribute to strain virulence [23,24,25,26,27]. Among the 23 investigated Xop genes in Xoc, avrBs2RS105 has been demonstrated to be required for full virulence of Xoc strains, while the xopAARS105 deletion enhanced strain virulence in rice JG30 [28]. It has been reported that the same Xop protein may contribute differently to virulence due to either different genetic backgrounds of strains or rice cultivars used for virulence assays.
The interaction between effectors and their targets is a cruel struggle between pathogen and host at molecular level. Rice–X. oryzae pathosystem is an excellent model for understanding effector biology, plant innate immunity, and other aspects of host–parasite interactions. In a long-time coevolution with pathogen, rice has evolved successful defense systems with the core of R genes to resist disease. The resistance strategies revealed so far of R genes adopted in rice–X. oryzae interactions mostly fit within two mechanisms: activating host innate immunity upon perception of the pathogen effectors and abolishing the host susceptibility through loss of interaction with effectors [29]. Interestingly, eight (Xa1, xa5, Xa10, xa13, Xa23, xa25, Xa27, xa41) out of 11 major rice resistance (R) genes cloned so far against X. oryzae mediate resistance associated with the TALEs in the course of rice–Xanthomonas interaction, which are grouped into three types of R genes, i.e., recessive, dominant nontranscriptional, and dominant TALE-dependent transcriptional resistance genes [30,31,32]. The other three cloned BB R genes are Xa3/Xa26, Xa4, and Xa21, which encode two types of receptor kinase proteins [33,34,35]. This review will focus on the diversified interactions between the major R genes or their products and the avirulence (Avr) proteins or virulent effectors in the rice–X. oryzae pathosystem to provide an overview of coevolution of rice innate immunity and the X. oryzae virulence.
2. Broad-Spectrum Resistance Mediated by Kinases
Plants have a precise immune system that is able to detect invading microbes and trigger a defense response [36]. The first line of innate immunity in plants, defined as PTI, is primarily made up of cell surface-localized pattern recognition receptors (PRRs), which can recognize a wide range of microbe- or pathogen-derived molecules, the so-called pathogen-associated molecular patterns (PAMPs), delivered into plant apoplast. This perception between PRRs and PAMPs triggers plant immune response (PTI) to resist a vast majority of microbes in the environment [36]. Receptor kinases (RKs) are the major components of PRRs and can alarm initial attacking from pathogens. For example, when rice is attacked by the fungus Magnaporthe oryzae, the host PRR OsCEBiP can recognize and bind the chitin from the fungus, forming a receptor complex with receptor-like kinases OsCERK1, and transduces signals to downstream proteins OsRacGEF1 and OsRac1 for innate immune responses [37,38].
The rice genome encodes more than 290 leucine-rich repeat (LRR) receptor kinases [39], and the well-characterized members—Xa21 and Xa3/Xa26—are representatives of non-RD (non-arginine-aspartate) RKs; both confer broad-spectrum resistance to Xoo strains. Xa21, originating from Oryza longistaminata, is the first cloned BB resistance gene and encodes a plasma membrane-localized receptor with extracellular LRR, transmembrane, juxtamembrane (JM), and cytoplasmic kinase domains (Figure 1) [35]. The function of this receptor has been intensively studied (Figure 2), and it is now clear that the JM domain plays an important role in the XA21 signal transduction pathway. In the absence of an Avr protein, XB24 (XA21 binding protein 24) binds XA21 and promotes phosphorylation of Ser/Thr sites (Ser686, Thr688, and Ser689) in the JM region through its ATPase activity, keeping the XA21 in an inactive state [40]. However, when rice plants face a challenge from Xoo, XA21 can recognize a sulfated Type-I-secreted protein RaxX, which is the Avr protein highly conserved in many Xanthomonas species [41]. Subsequently, XA21 dissociates from XB24 and excites the downstream defense response, e.g., XA21 transphosphorylates XB3, a ring figure ubiquitin ligase, to activate MAPK cascade. Furthermore, once the signal has been replayed, XA21 binds the XB15 phosphates, which attenuates the immune response, likely by dephosphorylation of amino acids required for XA21 function to negatively regulate the Xa21-mediated immune responses (Figure 2). RaxX is a 60-amino acid peptide similar to a PSY (plant peptide containing sulfated tyrosine) family peptide PSY1 in structure, suggesting that RaxX might have evolved to mimic plant hormone to regulate host biological process, thereby facilitating pathogen infection in the struggle [42]. Compared to Xoo, the RaxX homolog in Xoc is closer to another PSY peptide in rice OsPSY2 based on the three residues required for XA21 recognition.
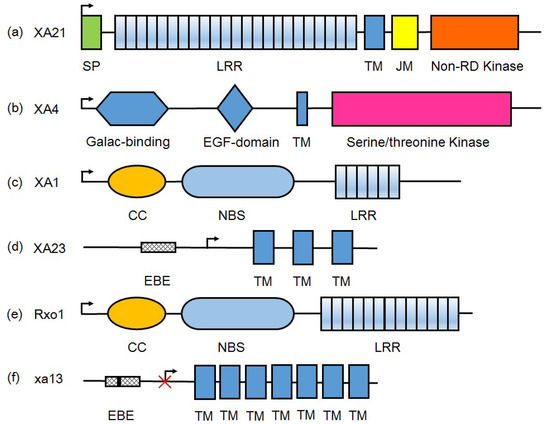
Figure 1.
The protein domain structures of representative R-gene products for Xoo resistance. (a) Receptor kinase, including XA21 and XA3/XA26. XA21 is exampled for illustrations here; (b) Wall-associated kinase, XA4; (c) Nucleotide-binding leucine-rich repeat (NLR)-type, XA1; (d) Executor R proteins, including Xa10, XA23, and XA27. Xa23 is exampled; (e) Nonhost R protein from maize, Rxo1; (f) SWEET protein encoded by recessive type R genes such as xa13, xa25, and xa41. These recessive type R genes are evolved by mutation (black vertical line in the EBE box) in promoter to block transcription activator-like effectors (TALE)-induced susceptibility. xa13 is exampled here. SP: signal peptide; LRR: leucine-rich repeat domain; TM: transmembrane domain; JM: juxtamembrane domain; non-RD: non-arginine-aspartate domain; Galac-binding: galacturonan-binding domain; EGF: calcium-binding epidermal growth factor domain; CC: coiled-coil domain; NBS: nucleotide binding site; EBE: effector binding element. Arrow indicates the direction of transcription.
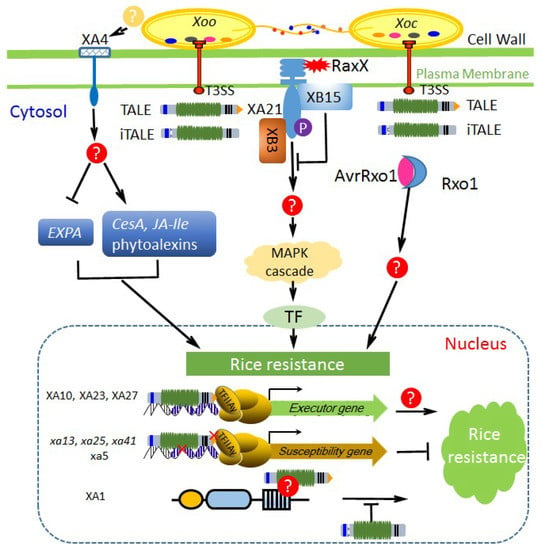
Figure 2.
The known pathways of rice major resistance against X. oryzae. X. oryzae secretes virulence factors (multicolor oval) via protein secretion systems (e.g., T3SS) to benefit the pathogens, but some were recognized by R gene products in rice to trigger innate immunity. XA21 localized at the apoplast recognizes tyrosine-sulfated protein RaxX (red sharp shape) from X. oryzae and activates defense response by phosphorylating downstream targets. XB3, a ring figure ubiquitin ligase, is required for effective XA21-mediated resistance and may participate in activating the MAPK (mitogen-activated protein kinase) cascade. XB15, a PP2C phosphatase, dephosphorylates autophosphorylated XA21 and negatively regulates the resistance. XA4 is a cell wall-associated kinase and can be activated by ligands from Xoo, leading to induction of CesAs expression, inhibition of EXPA expression, and accumulation of JA-Ile and phytoalexins. AvrRxo1 (pink oval) is a Xoc-specific effector and can interact with a maize NLR protein Rxo1 (light blue crescent), resulting in hypersensitive response (HR) induction in rice. Executor R genes, recessive R genes, and Xa1-like NLR genes evolved in rice represent different strategies of countering TALE attacks. iTALEs from X. oryzae are suppressors of Xa1-mediated resistance. Question marks (?) in red and yellow backgrounds indicate the unknown interactors from rice and X. oryzae, respectively.
Xa3/Xa26, another RK-type R gene, encodes a protein similar to XA21 in structure, mediating a different spectrum but also broad and durable resistance to Xoo strains [33,43,44,45]. A gene-dosage effect exists in Xa3/Xa26-mediated resistance—the higher expression level of Xa3/Xa26 confers a stronger, wider spectrum, and whole-growth-stage resistance. The Xa3/Xa26-mediated resistance is also influenced by rice genetic backgrounds: Oryza japonica background and WRKY45-1 allele are more appropriate for Xa3/Xa26 expression compared to Oryza indica background and WRKY45-2 allele [46].
The BB resistance gene Xa4 has been widely utilized in developing resistant varieties since 1970s [34]. It locates closely to the Xa3/X26 locus, encoding a cell wall-associated kinase (WAK) (Figure 1) and confers durable resistance to multiple Xoo races (e.g., Philippine race 1, 4, 5, 7, 8). Expression of Xa4 is rapidly and strongly induced in incompatible rice–Xoo interactions. In Xa4-carrying rice, expressions of cellulose synthase (CESA) family genes are enhanced, while expression of the α-expansin (EXPA) genes is suppressed. This regulation of multiple gene expression patterns enhances the mechanical strength of rice cell wall, as well as resistance against different biotic stress, and reduces plant height. Furthermore, expression induction of Xa4 in incompatible interactions may strengthen CesAs induction to reinforce the cell wall, exhibiting race-specific resistance (Figure 2). The Xa4-mediated resistance has been reported to be associated with increased accumulation of JA-Ile, sakuranetin, and momilactone A (Figure 2) [34].
3. Broad-Spectrum Resistance Mediated by the Xa1 Family Genes
The nucleotide-binding leucine-rich repeat (NLR) gene family is another big group of R genes in plants and the first cloned gene of this type genes conferring resistance against Xoo is Xa1, which is cloned from rice line IRBB1 [47]. Xa1 confers high levels of specific resistance to Japanese Xoo strain race 1 (T7174), but the mechanism of Xa1-mediated resistance is still unclear. A recent study on effects of deleting TALEs in the Xoo strain PXO99A revealed that Xa1 is actually a broad-spectrum resistance gene that can recognize different typical TALEs (tTALEs, identical with TALEs here) and initiate hypersensitive response (HR) cell death [10]. The activation of Xa1 resistance is independent of the number of TALE’s central repeats (if >3.5 repeats) and the RVDs composition of each repeat. In the natural habitat, such Xa1 resistance is suppressed by two groups of truncated TALEs (interfering TALEs, iTALEs), e.g., Tal3a and Tal3b in PXO99A. The Xa1-mediated resistance is activated by tTALE but suppressed by iTALE independently; both cases are prevalent in Xoo and Xoc isolates [10,48]. Compared with tTALEs, iTALEs lack 58 amino acids at the N-terminus and the transcriptional activation domain but retain one or two NLSs at the C-terminus, which are conserved in iTALEs and essential for interfering the Xa1-mediated resistance (Figure 2 and Figure 3). XA1 possesses a unique nucleotide binding site (NBS) region and its expression is induced by wounding and pathogen infection. The LRR domain located at the carboxy termini is the major diversified region among XA1 family members, and the LRR versions determine the output of resistance. Xa1 harbors six nearly identical LRR repeats (each repeat with 92 amino acids), but the first two LRR repeats of its susceptible allele (Os04g53120) still seem in the course of evolution (Figure 1).
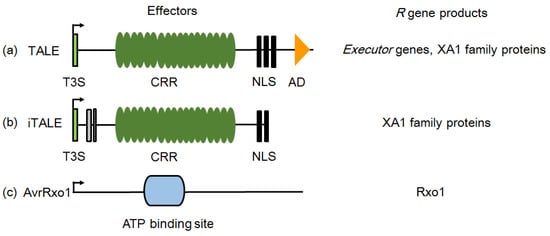
Figure 3.
Structural features of identified effector proteins participating in induction or suppression of major R gene resistances. (a) The typical structure of TALEs. TALEs contain an N-terminal type III secretion and translocation signal (T3S), a specific central repeat region (CRR), three nuclear localization signals (NLS), and a C-terminal transcription activation domain (AD). TALEs play multiple roles in rice–X. oryzae interaction and can be recognized by executor and Xa1 family genes. (b) The structure of type-A iTALEs. iTALEs are truncated variants of TALEs. Boxes with stripes represent deletions. In X. oryzae, iTALEs are prevalent inhibitors of resistance of Xa1 family genes. The different structures endow TALE and iTALE with opposite functions. (c) AvrRxo1 is similar to the zeta toxin protein, and an ATP-binding site exists in the central region. The targeted R gene products involved in susceptibility or resistance induced by the effectors are shown on the right.
What is more interesting is that T7174, the representative strain of Japanese Xoo race 1, shows incompatible interaction with IRBB1 (Xa1/Xa1), even though it harbors an iTALE (Tal3b type). Furthermore, expression of avrXa7 and avrXa10 (two tTALEs) or the AD (activation domian)-deleted tTALE pthXo1CRR (ΔAD) in rice variety Carolina Gold Select (Xo1/Xo1) can drastically decrease the virulence of Xoo strain X11-5A [31]. These findings suggest that the resistance function of the NLR proteins (here Xa1 and Xo1) may be inhibited by iTALEs. Similar observation has been recently described in a study on the Phytophthora sojae–soybean interaction [49]. It appears that iTALEs have been evolved as a paralogous decoy to neutralize the ability of NLR proteins. iTALEs themselves have no function in the development of disease, other than interfering NLR proteins in order to protect tTALE from recognition. Xa1 is not the sole NLR-type R protein that can recognize TALEs or its homologues because Bs4, a major R gene cloned from tomato, encodes a protein that also recognize the TALE member AvrBs4 from bacterial spot pathogen Xanthomonas campestris pv. vesicatoria in pepper [50]. The difference between Bs4 and Xa1 is that AD domain of elicitor is not required for Bs4 resistance elicitation as C-terminal-truncated avrBs4 can also trigger resistance. However, the mechanism of Bs4 resistance needs to be revealed as well.
4. Race-Specific Resistance Mediated by Recessive R Genes
TALE proteins were found primarily in plant pathogenic bacteria Xanthomonas species, Ralstonia solanacearum and Burkholderia rhizoxinica. They are special and significant weapons of pathogens in the rice–X. oryzae interaction [1]. PthXo1, AvrXa7, PthXo2, PthXo3, TalC, and Tal5 from Xoo and Tal2g from Xoc are all important virulence determinants, and except Tal2g, all of them target the SWEET family genes [13,14,51,52,53,54]. The first identified SWEET gene of TALE target is Os8N3 (also named as OsWEET11 and Xa13), which encodes a sucrose efflux transporter and can be bound and transcriptionally activated by PthXo1. Deletion of pthXo1 or silencing of Os8N3 leads to nearly 80% lesion length reduced upon PXO99A challenge [13]. The induction of SWEETs expression might facilitate sucrose movement through intracellular regions of plants, which provides nutrition to support bacterial multiplication and disease development [55]. Some other pathogens, such as Botrytis cinerea, also benefit from induction of SWEET gene expression during the infection process [56,57,58]. The expression of Os8N3 is essential for the pathogen but can be exchanged by the other clade III homologs [54]. It is astonishing that no native SWEET-targeting TALEs have been identified in Xoc, although SWEET gene expression induction can benefit Xoc infection [59]. Xoc genomic research indicates that Tal2gBLS256, together with four other TALEs, are highly conserved in Xoc. Experiment has shown that Tal2gBLS256 can activate a sulfate transporter-encoding gene expression, which is a major susceptible target for BLS. However, there is no explanation on the reason why a TALE contributes greatly for Xoc virulence by targeting the OsSULTR3;6 [52].
To counter the pathogen’s attack, plants mutated the effector binding elements (EBEs) in promoters of the S genes to disrupt the TALE-induced S target expression. For example, the recessive BB-resistant gene xa13 evolved from the susceptibility allele Xa13 by various nucleotide substitution, deletion, and insertion in the EBE region [60]. These nucleotide alterations in the promoter confer race-specific resistance to Xoo strains, which apply pthXo1 as the only major virulence factor, e.g., PXO99A. Similarly, xa25 is another recessive R gene and allelic to the S gene OsSWEET13, which is implicated as a target of major TALEs from Xoo strains PXO339 and PXO163 [61]. In addition, xa41 is an 18bp deletion promoter variant of OsSWEET14 present only in African rice wild species O. barthii and O. glaberrima [32].
An alternative route of rice to counter the action of pathogenic TALEs is to reduce the affinity of TALEs with plant transcription factor. The recessive xa5 is a natural allele of Xa5 (TFIIAγ5). TFIIAγ5 is the small (γ) subunit of the conserved general transcription factor TFIIA, which is important for polymerase II(Pol-II)-dependent transcription. The protein encoded by xa5 contains a mutation in the 39th residue, where the valine (V) residue is replaced with glutamine (E) (V39E). This missense mutation in xa5 attenuates the transcription of downstream TALE-targeted S genes, therefore improving rice resistance by abolishing the interaction between virulence factor TALEs and the preinitiation complex [62]. OsTFIIAγ1 and OsTFX1 are two other S genes associated with gene transcription; unfortunately, naturally resistant allele has still not been found. Although TFIIAγ1 do not directly interact with TALEs as TFIIAγ5, a recent study displayed that TFIIAγ1 can compensate for the absence of Xa5 when Xoo induces the expression of OsTFIIAγ1 by PthXo7 [63].
5. Race-Specific Resistance Mediated by Executor R Genes
In contrast to avoiding damage from pathogen by adopting recessive genes, rice employs another vigorous tactic to defend the fierce attacks from TALEs, which is more aggressive and effective (30). Executor R (E) genes encode a type of small proteins, conferring dominant resistance to X. oryzae and showing no similarity to any other known R gene products [30]. The primal TALE’s targets for bacteria are supposed to be some susceptibility (S) genes, because induction of S genes will benefit bacterial virulence and disease development. E genes possess specific EBEs in their promoter regions that can trap the cognate TALE effectors, e.g., AvrBs3 and AvrXa23. E genes are silent in normal conditions, but their expression occurs upon recognizing the Avr proteins from pathogen, which triggers immunity response in plants.
Thus far, five E genes have been cloned and three of them are from rice, e.g., Xa10, Xa23, and Xa27 [64,65,66], whose corresponding avr genes are the TALE-encoding genes avrXa10, avrXa23, and avrXa27, respectively. Zhang et al. grouped the E proteins into two groups: Group 1 is likely associated with plant development or physiology and group 2 are small proteins with multiple hydrophobic potential membrane-spanning domain [30]. Bs3, isolated from pepper, is the sole member of group 1, which is structurally related to flavin-dependent monooxygenases of the Arabidopsis YUCCA family, and plays roles in auxin biosynthesis and pathogen defense [67]. The E genes identified from rice are all classified into group 2. Actually, the E gene-related or similar coding sequences, which harbor polymorphism in promoters, are present in different rice cultivars and are mostly similar in function if induced by artificial TALEs. Xa27 is the first reported E gene in rice, encoding a 113-amino-acid protein localized to cytoplasmic membrane and apoplast. Compared to the promoter of susceptible allele xa27 in rice variety IR24, one GAA trinucleotide repeat insertion and a nucleotide substitution downstream the TATA box endows the resistant allele Xa27 in IRBB27 with an ability to recognize the natural TALE AvrXa27 [66]. Protein sequence analysis and function studies have indicated that the signal anchor-like sequence at amino (N)-terminal region of XA27 determine the protein localization and is essential for resistance to Xoo [68]. The Xa27 resistance is affected by growth stages and sometimes genetic backgrounds. Xa10 and Xa23 reside at two adjacent loci on chromosome 11 and are closely related members, sharing 64% similarity in DNA sequences and 50% identity at protein level [64,69]. Both XA10 and XA23 are endoplasmic reticulum-localized proteins and can induce HR in nonhost plants such as Nicotiana benthamiana. XA10 and XA10-like R proteins (e.g., XA10-Ni) have four predicted transmembrane helices, whereas XA23 or XA23-like R proteins have only three predicted transmembrane helices in short N-terminal region [70]. An ED or ED-like motif, comprising acidic amino acids residues (D and E), appears at the C-termini of XA10, XA23, and XA27. This specific domain in XA10 and XA23 has been proven essential for HR induction in tobacco and resistance in rice. All the E proteins appear to be toxic to plant cells, and expression of E genes can elicit hypersensitive response (HR) to restrict pathogen growth and diffusion. Further experimentation has revealed that XA10 forms hexamers, then induces ER Ca2+ depletion, and triggers cell death in rice, N. benthamiana, and human HeLa cells; however, details about how an E protein switches on the HR in host plants is still unclear [65].
Intriguingly, no cognate S genes of the TALEs AvrXa10, AvrXa23, and AvrXa27 have been identified. AvrXa27 and AvrXa23 are conserved in various Xoo strains; therefore, Xa27 and Xa23 have been widely applied in rice breeding programs, and the resulting varieties show broad-spectrum resistance against Xoo strains. However, none of the three E genes has been utilized to control BLS because natural Xoc strains do not carry the cognate TALEs. Additionally, it has been shown that XA10-mediated resistance is sometimes not strong enough to revolt attacks from Xoc, even if the avr gene AvrXa10 was introduced into Xoc by electrotransformation [71].
6. Durable and Broad-Spectrum Resistance Mediated by Nonhost R Genes
In contrast to the host disease resistance genes, nonhost resistance genes usually target conserved effectors and show durable and broad-spectrum resistance [72]. Rxo1 (the same gene as Rba1) is a maize NLR-type R gene conferring resistance against the maize/sorghum bacterial stripe pathogen Burkholderia andropogonis [73]. Rxo1 conditions a typical HR phenotype in maize and rice upon presence of AvrRxo1 (also termed as XopAJ) or its homolog from Burkholderia andropogonis and nonhost bacterial pathogen Xoc or X. euvesicatoria [74]. However, AvrRxo1, a conserved bacterial toxin in Asian Xoc strains and some African Xoc strains, can enhance Xoo/Xoc early proliferation in rice leaves and suppress HR induced by Xoo/Xoc in nonhost N. benthamiana (Figure 3) [75,76]. The toxicity and virulence contribution of AvrRxo1 might be associated with its nucleotide kinase activity, where AvrRxo1 phosphorylates NAD at the 3′ hydroxyl position to produce 3′-NADP [77]. NAD is a coenzyme and redox carrier that is critically required for organism metabolism function and is an important signal in early events of plant defense responses. AvrRxo1 consumes NAD through direct phosphorylation to manipulate host metabolism and immunity, and the product 3′-NADP could be an inhibitor of defense-related oxidative burst in plants.
The way in which Rxo1 interacts with AvrRxo1 and activates resistance is still a question that needs to be elucidated. A microarray experiment revealed that Rxo1 functions in the early stage of Xoc infection and possibly activates multiple pathways associated with basal defense and HR, such as SA and ET pathways [78]. Besides Rxo1, six Xop genes from Xoc were transiently expressed and caused HR in N. benthamiana, indicating that candidate R genes existing in tobacco could be exploited as nonhost R genes to improve rice resistance [28].
7. Conclusions and Perspectives
The rice–X. oryzae pathosystem provides a model system for studying plant–pathogen interactions at a molecular level. Intensive investigations in the past decade have revealed that the resistance and susceptibility of rice to BB and BLS are generally controlled by the molecular interactions between X. oryzae effectors and its target genes in rice. Among the 11 rice R genes against X. oryzae characterized so far, eight (Xa1, xa5, Xa10, xa13, Xa23, xa25, Xa27, xa41) mediate resistance associated with TALEs from the pathogen; only three (Xa3/Xa26, Xa4, and Xa21) encode kinase proteins and mediate rice resistance. This is very special because NLR-type R genes are the major players in other plant–pathogen pathosystems, e.g., rice–M. oryzae [79]. Moreover, the TALE-associated rice R genes against X. oryzae play different roles. Based on these advanced information, this review summed up five routes that rice has evolved or has adopted to defend attacks from X. oryzae.
Although enormous progress in characterizing rice–X. oryzae interaction and breeding for resistance has been achieved since the molecular cloning of the first R gene Xa21 against Xoo, many questions and challenges still remain. For example, nearly a dozen R genes against Xoo have been cloned, but no effective major R gene against Xoc has been discovered in rice. In addition, the BB R genes cannot be used for BLS resistance in rice breeding, although Xoo and Xoc are close species and even possess similar biological weapons, e.g., Xop proteins and TALEs. The lack of endogenous BLS R gene can be partially explained for immense and expansionary TALomes of Xoc. The Xoc strains may target diverse S genes to exploit nutrients, which is different from other Xanthomonas species. Another factor is that Xoc TALEs function to suppress multiple resistance, for example, both AvrXa7 and Tal2aBLS256 with Xoc background failed to initiate expected HR in the corresponding resistant rice [16,80]. Elucidation of the function mechanism of TALE-like proteins (Tal7RS105 and Tal3aPXO99A) exerting immunity suppressors is the basis to engineer effective R genes against Xoc and also Xoo. Up to now, the genetic working model of executor R genes has been well addressed. However, many biochemical details about how an E protein switches on the HR in host cells is still to be elucidated, even though there is a clue associated with induced ER Ca2+ depletion [65]. Elucidation of the detailed biochemical mechanism of the executor R genes is a great challenge because the cell death is too fast to track the biochemical process easily.
The Xoo and Xoc populations exhibit high genetic diversity and present rapid evolution in natural environments. New aggressive strains/races have often emerged, eroded the R gene resistance, and expanded in different regions, probably due to strong host selection, changing ecological conditions, and farm managements [81]. In this scenario, natural genetic resources may not be enough to continuously breed new, resistant rice cultivars in the future. TALENs (transcription activator-like effector nucleases) and CRISPR (clustered regularly interspaced short palindromic repeats)/Cas9 (CRISPR-associated 9) system-based genome editing technologies have become essential to develop new crop varieties with durable and broad-spectrum resistance. To this aim, Os11N3 (OsSWEET14), the S gene targeted by AvrXa7 and PthXo3, has been edited by TALENs to create BB-resistant rice through disrupting the EBE site in the promoter region [82,83]. Moreover, CRISPR/Cas9 is a new, simple, and more effective tool for genome editing system, which even enables multiplex gene editing in many organisms, including rice [84,85]. Two recent reports have provided successful examples of enhancing rice blast resistance via CRISPR/Cas9-mediated knock-out of OsERF922 and Bsr-d1 [86,87]. It appears that the CRISPR/Cas9 will become an efficient and frequently used technology, accelerating both basic and applied research on the rice–X. oryzae pathosystem. Except for genome editing, artificial E genes with designed EBE could become another effective approach to achieve plant resistance because a designed EBE can trap multiple or widely conserved TALEs that enable the artificial E gene to resist X. oryzae strains broadly [88,89].
Author Contributions
The manuscript was written by Z.J. and C.W. under the supervision of K.Z. The manuscript was critically revised by K.Z. All the authors have read and approved the final manuscript.
Funding
This work was supported by grants from the National Priority Program—Breeding New Rice Varieties for Southwest and Southern China Area (2017YFD0100202; 2017YFD0100100), the Young Elite Scientist Sponsorship of China Association for Science and Technology (2017QNRC001) and the Innovation Program of Chinese Academy of Agricultural Sciences to Kaijun Zhao and Chunlian Wang.
Acknowledgments
We thank Rezwan Traiq (NFCRI) for critical reading of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Boch, J.; Bonas, U. Xanthomonas AvrBs3 family-type III effectors: Discovery and function. Annu. Rev. Phytopathol. 2010, 48, 419–436. [Google Scholar] [CrossRef] [PubMed]
- Buttner, D.; Bonas, U. Regulation and secretion of Xanthomonas virulence factors. FEMS Microbiol. Rev. 2010, 34, 107–133. [Google Scholar] [CrossRef] [PubMed]
- Nino-Liu, D.O.; Ronald, P.C.; Bogdanove, A.J. Xanthomonas oryzae pathovars: Model pathogens of a model crop. Mol. Plant Pathol. 2006, 7, 303–324. [Google Scholar] [CrossRef] [PubMed]
- White, F.F.; Yang, B. Host and pathogen factors controlling the rice-Xanthomonas oryzae interaction. Plant Physiol. 2009, 150, 1677–1686. [Google Scholar] [CrossRef] [PubMed]
- Moscou, M.J.; Bogdanove, A.J. A simple cipher governs DNA recognition by TAL effectors. Science 2009, 326, 1501. [Google Scholar] [CrossRef] [PubMed]
- Deng, D.; Yan, C.; Pan, X.; Mahfouz, M.; Wang, J.; Zhu, J.K.; Shi, Y.; Yan, N. Structural basis for sequence-specific recognition of DNA by TAL effectors. Science 2012, 335, 720–723. [Google Scholar] [CrossRef] [PubMed]
- Boch, J.; Scholze, H.; Schornack, S.; Landgraf, A.; Hahn, S.; Kay, S.; Lahaye, T.; Nickstadt, A.; Bonas, U. Breaking the code of DNA binding specificity of TAL-type III effectors. Science 2009, 326, 1509–1512. [Google Scholar] [CrossRef] [PubMed]
- Boch, J.; Bonas, U.; Lahaye, T. TAL effectors—Pathogen strategies and plant resistance engineering. New Phytol. 2014, 204, 823–832. [Google Scholar] [CrossRef] [PubMed]
- Cox, K.L.; Meng, F.; Wilkins, K.E.; Li, F.; Wang, P.; Booher, N.J.; Carpenter, S.C.D.; Chen, L.Q.; Zheng, H.; Gao, X.; et al. TAL effector driven induction of a SWEET gene confers susceptibility to bacterial blight of cotton. Nat. Commun. 2017, 8, 15588. [Google Scholar] [CrossRef] [PubMed]
- Ji, Z.; Ji, C.; Liu, B.; Zou, L.; Chen, G.; Yang, B. Interfering TAL effectors of Xanthomonas oryzae neutralize R-gene-mediated plant disease resistance. Nat. Commun. 2016, 7, 13435. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Zhang, J.; Jia, H.; Sosso, D.; Li, T.; Frommer, W.B.; Yang, B.; White, F.F.; Wang, N.; Jones, J.B. Lateral organ boundaries 1 is a disease susceptibility gene for citrus bacterial canker disease. Proc. Natl Acad. Sci. USA 2014, 111, E521–E529. [Google Scholar] [CrossRef] [PubMed]
- Kay, S.; Hahn, S.; Marois, E.; Hause, G.; Bonas, U. A bacterial effector acts as a plant transcription factor and induces a cell size regulator. Science 2007, 318, 648–651. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Sugio, A.; White, F.F. Os8N3 is a host disease-susceptibility gene for bacterial blight of rice. Proc. Natl. Acad. Sci. USA 2006, 103, 10503–10508. [Google Scholar] [CrossRef] [PubMed]
- Antony, G.; Zhou, J.H.; Huang, S.; Li, T.; Liu, B.; White, F.; Yang, B. Rice xa13 recessive resistance to bacterial blight is defeated by induction of the disease susceptibility gene Os-11N3. Plant Cell 2010, 22, 3864–3876. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Wang, S. Rice MtN3/saliva/SWEET family genes and their homologs in cellular organisms. Mol. Plant 2013, 6, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Cao, Y.; Xu, Z.; Ma, W.; Zakria, M.; Zou, L.; Cheng, Z.; Chen, G. A transcription activator-like effector Tal7 of Xanthomonas oryzae pv. oryzicola activates rice gene Os09g29100 to suppress rice immunity. Sci. Rep. 2017, 7, 5089. [Google Scholar]
- Salzberg, S.L.; Sommer, D.D.; Schatz, M.C.; Phillippy, A.M.; Rabinowicz, P.D.; Tsuge, S.; Furutani, A.; Ochiai, H.; Delcher, A.L.; Kelley, D.; et al. Genome sequence and rapid evolution of the rice pathogen Xanthomonas oryzae pv. oryzae PXO99A. BMC Genom. 2008, 9, 204. [Google Scholar] [CrossRef]
- Bogdanove, A.J.; Koebnik, R.; Lu, H.; Furutani, A.; Angiuoli, S.V.; Patil, P.B.; Van Sluys, M.A.; Ryan, R.P.; Meyer, D.F.; Han, S.W.; et al. Two new complete genome sequences offer insight into host and tissue specificity of plant pathogenic Xanthomonas spp. J. Bacteriol. 2011, 193, 5450–5464. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, K.E.; Booher, N.J.; Wang, L.; Bogdanove, A.J. TAL effectors and activation of predicted host targets distinguish Asian from African strains of the rice pathogen Xanthomonas oryzae pv. oryzicola while strict conservation suggests universal importance of five TAL effectors. Front. Plant Sci. 2015, 6, 536. [Google Scholar] [PubMed]
- Yamaguchi, K.; Yamada, K.; Ishikawa, K.; Yoshimura, S.; Hayashi, N.; Uchihashi, K.; Ishihama, N.; Kishi-Kaboshi, M.; Takahashi, A.; Tsuge, S.; et al. A receptor-like cytoplasmic kinase targeted by a plant pathogen effector is directly phosphorylated by the chitin receptor and mediates rice immunity. Cell Host Microbe 2013, 13, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, K.; Yamaguchi, K.; Sakamoto, K.; Yoshimura, S.; Inoue, K.; Tsuge, S.; Kojima, C.; Kawasaki, T. Bacterial effector modulation of host E3 ligase activity suppresses PAMP-triggered immunity in rice. Nat. Commun. 2014, 5, 5430. [Google Scholar] [CrossRef] [PubMed]
- Cheong, H.; Kim, C.Y.; Jeon, J.S.; Lee, B.M.; Sun Moon, J.; Hwang, I. Xanthomonas oryzae pv. oryzae type III effector XopN targets OsVOZ2 and a putative thiamine synthase as a virulence factor in rice. PLoS ONE 2013, 8, e73346. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Yang, B. Mutagenesis of 18 type III effectors reveals virulence function of XopZPXO99 in Xanthomonas oryzae pv. oryzae. Mol. Plant Microbe Interact. 2010, 23, 893–902. [Google Scholar] [CrossRef] [PubMed]
- Mondal, K.K.; Verma, G.; Manju; Junaid, A.; Mani, C. Rice pathogen Xanthomonas oryzae pv. oryzae employs inducible hrp-dependent XopF type III effector protein for its growth, pathogenicity and for suppression of PTI response to induce blight disease. Eur. J. Plant Pathol. 2016, 144, 311–323. [Google Scholar] [CrossRef]
- Akimoto-Tomiyama, C.; Furutani, A.; Tsuge, S.; Washington, E.J.; Nishizawa, Y.; Minami, E.; Ochiai, H. XopR, a type III effector secreted by Xanthomonas oryzae pv. oryzae, suppresses microbe-associated molecular pattern-triggered immunity in Arabidopsis thaliana. Mol. Plant Microbe Interact. 2012, 25, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.K.; Nathawat, R.; Sinha, D.; Haque, A.S.; Sankaranarayanan, R.; Sonti, R.V. Mutations in the predicted active site of Xanthomonas oryzae pv. oryzae XopQ differentially affect virulence, suppression of host innate immunity, and induction of the HR in a nonhost plant. Mol. Plant Microbe Interact. 2015, 28, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Sinha, D.; Gupta, M.K.; Patel, H.K.; Ranjan, A.; Sonti, R.V. Cell wall degrading enzyme induced rice innate immune responses are suppressed by the type 3 secretion system effectors XopN, XopQ, XopX and XopZ of Xanthomonas oryzae pv. oryzae. PLoS ONE 2013, 8, e75867. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, Y.; Wang, S.; Fang, A.; Wang, J.; Liu, L.; Zhang, K.; Mao, Y.; Sun, W. The Type III effector AvrBs2 in Xanthomonas oryzae pv. oryzicola suppresses rice immunity and promotes disease development. Mol. Plant Microbe Interact. 2015, 28, 869–880. [Google Scholar] [CrossRef] [PubMed]
- Kourelis, J.; van der Hoorn, R.A.L. Defended to the nines: 25 years of resistance gene cloning identifies nine mechanisms for R protein function. Plant Cell 2018, 30, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yin, Z.; White, F. TAL effectors and the executor R genes. Front. Plant Sci. 2015, 6, 641. [Google Scholar] [CrossRef] [PubMed]
- Triplett, L.R.; Cohen, S.P.; Heffelfinger, C.; Schmidt, C.L.; Huerta, A.I.; Tekete, C.; Verdier, V.; Bogdanove, A.J.; Leach, J.E. A resistance locus in the American heirloom rice variety Carolina Gold Select is triggered by TAL effectors with diverse predicted targets and is effective against African strains of Xanthomonas oryzae pv. oryzicola. Plant J. 2016, 87, 472–483. [Google Scholar] [CrossRef] [PubMed]
- Hutin, M.; Sabot, F.; Ghesquiere, A.; Koebnik, R.; Szurek, B. A knowledge-based molecular screen uncovers a broad-spectrum OsSWEET14 resistance allele to bacterial blight from wild rice. Plant J. 2015, 84, 694–703. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Cao, Y.; Yang, Z.; Xu, C.; Li, X.; Wang, S.; Zhang, Q. Xa26, a gene conferring resistance to Xanthomonas oryzae pv. oryzae in rice, encodes an LRR receptor kinase-like protein. Plant J. 2004, 37, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Cao, J.; Zhang, J.; Xia, F.; Ke, Y.; Zhang, H.; Xie, W.; Liu, H.; Cui, Y.; Cao, Y.; et al. Improvement of multiple agronomic traits by a disease resistance gene via cell wall reinforcement. Nat. Plants 2017, 3, 17009. [Google Scholar] [CrossRef] [PubMed]
- Song, W.Y.; Wang, G.L.; Chen, L.L.; Kim, H.S.; Pi, L.Y.; Holsten, T.; Gardner, J.; Wang, B.; Zhai, W.X.; Zhu, L.H.; et al. A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21. Science 1995, 270, 1804–1806. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Akamatsu, A.; Wong, H.L.; Fujiwara, M.; Okuda, J.; Nishide, K.; Uno, K.; Imai, K.; Umemura, K.; Kawasaki, T.; Kawano, Y.; et al. An OsCEBiP/OsCERK1-OsRacGEF1-OsRac1 module is an essential early component of chitin-induced rice immunity. Cell Host Microbe 2013, 13, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.Z.; Wang, G.X.; Zhou, J.M. Receptor kinases in plant-pathogen interactions: More than pattern recognition. Plant Cell 2017, 29, 618–637. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.L.; Xue, H.W. Global analysis of expression profiles of rice receptor-like kinase genes. Mol. Plant 2012, 5, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Park, C.J.; Han, S.W.; Chen, X.; Ronald, P.C. Elucidation of XA21-mediated innate immunity. Cell Microbiol. 2010, 12, 1017–1025. [Google Scholar] [CrossRef] [PubMed]
- Pruitt, R.N.; Schwessinger, B.; Joe, A.; Thomas, N.; Liu, F.; Albert, M.; Robinson, M.R.; Chan, L.J.; Luu, D.D.; Chen, H.; et al. The rice immune receptor XA21 recognizes a tyrosine-sulfated protein from a Gram-negative bacterium. Sci. Adv. 2015, 1, e1500245. [Google Scholar] [CrossRef] [PubMed]
- Pruitt, R.N.; Joe, A.; Zhang, W.G.; Feng, W.; Stewart, V.; Schwessinger, B.; Dinneny, J.R.; Ronald, P.C. A microbially derived tyrosine-sulfated peptide mimics a plant peptide hormone. New Phytol. 2017, 215, 725–736. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Ding, X.; Cai, M.; Zhao, J.; Lin, Y.; Li, X.; Xu, C.; Wang, S. The expression pattern of a rice disease resistance gene Xa3/Xa26 is differentially regulated by the genetic backgrounds and developmental stages that influence its function. Genetics 2007, 177, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Li, H.J.; Li, X.H.; Xiao, J.H.; Wing, R.A.; Wang, S.P. Ortholog alleles at Xa3/Xa26 locus confer conserved race-specific resistance against Xanthomonas oryzae in rice. Mol. Plant 2012, 5, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Cao, Y.L.; Xu, C.G.; Li, X.H.; Wang, S.P. Xa3, conferring resistance for rice bacterial blight and encoding a receptor kinase-like protein, is the same as Xa26. Theor. Appl. Genet. 2006, 113, 1347–1355. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Liu, H.; Zhou, Y.; Zhang, Q.; Li, X.; Wang, S. Exploring the mechanism and efficient use of a durable gene-mediated resistance to bacterial blight disease in rice. Mol. Breed. 2018, 38. [Google Scholar] [CrossRef]
- Yoshimura, S.; Yamanouchi, U.; Katayose, Y.; Toki, S.; Wang, Z.X.; Kono, I.; Kurata, N.; Yano, M.; Iwata, N.; Sasaki, T. Expression of Xa1, a bacterial blight-resistance gene in rice, is induced by bacterial inoculation. Proc. Natl. Acad. Sci. USA 1998, 95, 1663–1668. [Google Scholar] [CrossRef] [PubMed]
- Read, A.C.; Rinaldi, F.C.; Hutin, M.; He, Y.-Q.; Triplett, L.R.; Bogdanove, A.J. Suppression of Xo1-mediated disease resistance in rice by a truncated, non-DNA-binding TAL effector of Xanthomonas oryzae. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Zhu, L.; Song, T.; Wang, Y.; Zhang, Q.; Xia, Y.; Qiu, M.; Lin, Y.; Li, H.; Kong, L.; et al. A paralogous decoy protects Phytophthora sojae apoplastic effector PsXEG1 from a host inhibitor. Science 2017, 355, 710–714. [Google Scholar] [CrossRef] [PubMed]
- Schornack, S.; Ballvora, A.; Gürlebeck, D.; Peart, J.; Ganal, M.; Baker, B.; Bonas, U.; Lahaye, T. The tomato resistance protein Bs4 is a predicted non-nuclear TIR-NB-LRR protein that mediates defense responses to severely truncated derivatives of AvrBs4 and overexpressed AvrBs3. Plant J. 2004, 37, 46–60. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Peng, Z.; Long, J.; Sosso, D.; Liu, B.; Eom, J.S.; Huang, S.; Liu, S.; Vera Cruz, C.; Frommer, W.B.; et al. Gene targeting by the TAL effector PthXo2 reveals cryptic resistance gene for bacterial blight of rice. Plant J. 2015, 82, 632–643. [Google Scholar] [CrossRef] [PubMed]
- Cernadas, R.A.; Doyle, E.L.; Nino-Liu, D.O.; Wilkins, K.E.; Bancroft, T.; Wang, L.; Schmidt, C.L.; Caldo, R.; Yang, B.; White, F.F.; et al. Code-assisted discovery of TAL effector targets in bacterial leaf streak of rice reveals contrast with bacterial blight and a novel susceptibility gene. PLoS Pathog. 2014, 10, e1003972. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Streubel, J.; Balzergue, S.; Champion, A.; Boch, J.; Koebnik, R.; Feng, J.; Verdier, V.; Szurek, B. Colonization of rice leaf blades by an African strain of Xanthomonas oryzae pv. oryzae depends on a new TAL effector that induces the rice nodulin-3 Os11N3 gene. Mol. Plant Microbe Interact. 2011, 24, 1102–1113. [Google Scholar] [CrossRef] [PubMed]
- Streubel, J.; Pesce, C.; Hutin, M.; Koebnik, R.; Boch, J.; Szurek, B. Five phylogenetically close rice SWEET genes confer TAL effector-mediated susceptibility to Xanthomonas oryzae pv. oryzae. New Phytol. 2013, 200, 808–819. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Q.; Hou, B.H.; Lalonde, S.; Takanaga, H.; Hartung, M.L.; Qu, X.Q.; Guo, W.J.; Kim, J.G.; Underwood, W.; Chaudhuri, B.; et al. Sugar transporters for intercellular exchange and nutrition of pathogens. Nature 2010, 468, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Piron, M.C.; Meyer, S.; Merdinoglu, D.; Bertsch, C.; Mestre, P. The SWEET family of sugar transporters in grapevine: VvSWEET4 is involved in the interaction with Botrytis cinerea. J. Exp. Bot. 2014, 65, 6589–6601. [Google Scholar] [CrossRef] [PubMed]
- Asai, Y.; Kobayashi, Y. Increased expression of the tomato SISWEET15 gene during grey mold infection and the possible involvement of the sugar efflux to apoplasm in the disease susceptibility. J. Plant Pathol. Microbiol. 2016, 7. [Google Scholar] [CrossRef]
- Chen, L.Q.; Qu, X.Q.; Hou, B.H.; Sosso, D.; Osorio, S.; Fernie, A.R.; Frommer, W.B. Sucrose efflux mediated by SWEET proteins as a key step for phloem transport. Science 2012, 335, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Verdier, V.; Triplett, L.R.; Hummel, A.W.; Corral, R.; Cernadas, R.A.; Schmidt, C.L.; Bogdanove, A.J.; Leach, J.E. Transcription activator-like (TAL) effectors targeting OsSWEET genes enhance virulence on diverse rice (Oryza sativa) varieties when expressed individually in a TAL effector-deficient strain of Xanthomonas oryzae. New Phytol. 2012, 196, 1197–1207. [Google Scholar] [CrossRef] [PubMed]
- Chu, Z.; Yuan, M.; Yao, J.; Ge, X.; Yuan, B.; Xu, C.; Li, X.; Fu, B.; Li, Z.; Bennetzen, J.L.; et al. Promoter mutations of an essential gene for pollen development result in disease resistance in rice. Genes Dev. 2006, 20, 1250–1255. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Yuan, M.; Zhou, Y.; Li, X.; Xiao, J.; Wang, S. A paralog of the MtN3/saliva family recessively confers race-specific resistance to Xanthomonas oryzae in rice. Plant Cell Environ. 2011, 34, 1958–1969. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Ke, Y.G.; Huang, R.Y.; Ma, L.; Yang, Z.Y.; Chu, Z.H.; Xiao, J.H.; Li, X.H.; Wang, S.P. A host basal transcription factor is a key component for infection of rice by TALE-carrying bacteria. eLife 2016, 5, e19605. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Zou, L.; Ji, Z.; Xu, X.; Xu, Z.; Yang, Y.; Alfano, J.R.; Chen, G. Xanthomonas oryzae pv. oryzae TALE proteins recruit OsTFIIAγ1 to compensate for the absence of OsTFIIAγ5 in bacterial blight in rice. Mol. Plant Pathol. 2018, 19, 2248–2262. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.L.; Zhang, X.P.; Fan, Y.L.; Gao, Y.; Zhu, Q.L.; Zheng, C.K.; Qin, T.F.; Li, Y.Q.; Che, J.Y.; Zhang, M.W.; et al. XA23 is an executor R protein and confers broad-spectrum disease resistance in rice. Mol. Plant 2015, 8, 290–302. [Google Scholar] [CrossRef] [PubMed]
- Tian, D.; Wang, J.; Zeng, X.; Gu, K.; Qiu, C.; Yang, X.; Zhou, Z.; Goh, M.; Luo, Y.; Murata-Hori, M.; et al. The rice TAL effector-dependent resistance protein XA10 triggers cell death and calcium depletion in the endoplasmic reticulum. Plant Cell 2014, 26, 497–515. [Google Scholar] [CrossRef] [PubMed]
- Gu, K.; Yang, B.; Tian, D.; Wu, L.; Wang, D.; Sreekala, C.; Yang, F.; Chu, Z.; Wang, G.L.; White, F.F.; et al. R gene expression induced by a type-III effector triggers disease resistance in rice. Nature 2005, 435, 1122–1125. [Google Scholar] [CrossRef] [PubMed]
- Romer, P.; Hahn, S.; Jordan, T.; Strauβ, T.; Bonas, U.; Lahaye, T. Plant pathogen recognition mediated by promoter activation of the pepper Bs3 resistance gene. Science 2007, 318, 645–648. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Goh, M.L.; Sreekala, C.; Yin, Z. XA27 depends on an amino-terminal signal-anchor-like sequence to localize to the apoplast for resistance to Xanthomonas oryzae pv oryzae. Plant Physiol. 2008, 148, 1497–1509. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.L.; Qin, T.F.; Yu, H.M.; Zhang, X.P.; Che, J.Y.; Gao, Y.; Zheng, C.K.; Yang, B.; Zhao, K.J. The broad bacterial blight resistance of rice line CBB23 is triggered by a novel transcription activator-like (TAL) effector of Xanthomonas oryzae pv. oryzae. Mol. Plant Pathol. 2014, 15, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Tian, D.; Gu, K.; Yang, X.; Wang, L.; Zeng, X.; Yin, Z. Induction of Xa10-like Genes in Rice Cultivar Nipponbare Confers Disease Resistance to Rice Bacterial Blight. Mol. Plant Microbe Interact. 2017, 30, 466–477. [Google Scholar] [CrossRef] [PubMed]
- Makino, S.; Sugio, A.; White, F.; Bogdanove, A.J. Inhibition of resistance gene-mediated defense in rice by Xanthomonas oryzae pv. oryzicola. Mol. Plant Microbe Interact. 2006, 19, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Niks, R.E.; Marcel, T.C. Nonhost and basal resistance: How to explain specificity? New Phytol. 2009, 182, 817–828. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Lin, X.; Poland, J.; Trick, H.; Leach, J.; Hulbert, S. A maize resistance gene functions against bacterial streak disease in rice. Proc. Natl. Acad. Sci. USA 2005, 102, 15383–15388. [Google Scholar] [CrossRef] [PubMed]
- Triplett, L.R.; Shidore, T.; Long, J.; Miao, J.; Wu, S.; Han, Q.; Zhou, C.; Ishihara, H.; Li, J.; Zhao, B.; et al. AvrRxo1 is a bifunctional type III secreted effector and toxin-antitoxin system component with homologs in diverse environmental contexts. PLoS ONE 2016, 11, e0158856. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Zhou, C.; Wu, S.; Liu, Y.; Triplett, L.; Miao, J.; Tokuhisa, J.; Deblais, L.; Robinson, H.; Leach, J.E.; et al. Crystal structure of Xanthomonas AvrRxo1-ORF1, a type III effector with a polynucleotide kinase domain, and its interactor AvrRxo1-ORF2. Structure 2015, 23, 1900–1909. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Chang, Q.; Feng, W.; Zhang, B.; Wu, T.; Li, N.; Yao, F.; Ding, X.; Chu, Z. Domain dissection of AvrRxo1 for suppressor, avirulence and cytotoxicity functions. PLoS ONE 2014, 9, e113875. [Google Scholar] [CrossRef] [PubMed]
- Shidore, T.; Broeckling, C.D.; Kirkwood, J.S.; Long, J.J.; Miao, J.; Zhao, B.; Leach, J.E.; Triplett, L.R. The effector AvrRxo1 phosphorylates NAD in planta. PLoS Pathog. 2017, 13, e1006442. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.L.; Xu, M.R.; Zhao, M.F.; Xie, X.W.; Zhu, L.H.; Fu, B.Y.; Li, Z.K. Genome-wide gene responses in a transgenic rice line carrying the maize resistance gene Rxo1 to the rice bacterial streak pathogen, Xanthomonas oryzae pv. oryzicola. BMC Genom. 2010, 11, 78. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.D.; Wang, G.L. Plant innate immunity in rice: A defense against pathogen infection. Natl. Sci. Rev. 2016, 3, 295–308. [Google Scholar] [CrossRef]
- Hummel, A.W.; Wilkins, K.E.; Wang, L.; Cernadas, R.A.; Bogdanove, A.J. A transcription activator-like effector from Xanthomonas oryzae pv. oryzicola elicits dose-dependent resistance in rice. Mol. Plant Pathol. 2017, 18, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Quibod, I.L.; Perez-Quintero, A.; Booher, N.J.; Dossa, G.S.; Grande, G.; Szurek, B.; Vera Cruz, C.; Bogdanove, A.J.; Oliva, R. Effector diversification contributes to Xanthomonas oryzae pv. oryzae phenotypic adaptation in a semi-isolated environment. Sci. Rep. 2016, 6, 34137. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Liu, B.; Spalding, M.H.; Weeks, D.P.; Yang, B. High-efficiency TALEN-based gene editing produces disease-resistant rice. Nat. Biotechnol. 2012, 30, 390–392. [Google Scholar] [CrossRef] [PubMed]
- Blanvillain-Baufume, S.; Reschke, M.; Sole, M.; Auguy, F.; Doucoure, H.; Szurek, B.; Meynard, D.; Portefaix, M.; Cunnac, S.; Guiderdoni, E.; et al. Targeted promoter editing for rice resistance to Xanthomonas oryzae pv. oryzae reveals differential activities for SWEET14-inducing TAL effectors. Plant Biotechnol. J. 2017, 15, 306–317. [Google Scholar] [CrossRef] [PubMed]
- Xie, K.; Minkenberg, B.; Yang, Y. Boosting CRISPR/Cas9 multiplex editing capability with the endogenous tRNA-processing system. Proc. Natl. Acad. Sci. USA 2015, 112, 3570–3675. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, J.; Wei, P.; Zhang, B.; Gou, F.; Feng, Z.; Mao, Y.; Yang, L.; Zhang, H.; Xu, N.; et al. The CRISPR/Cas9 system produces specific and homozygous targeted gene editing in rice in one generation. Plant Biotechnol. J. 2014, 12, 797–807. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.J.; Wang, C.L.; Liu, P.Q.; Lei, C.L.; Hao, W.; Gao, Y.; Liu, Y.G.; Zhao, K.J. Enhanced rice blast resistance by CRISPR/Cas9-targeted mutagenesis of the ERF transcription factor gene OsERF922. PLoS ONE 2016, 11, e0154027. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhu, Z.; Chern, M.; Yin, J.; Yang, C.; Ran, L.; Cheng, M.; He, M.; Wang, K.; Wang, J.; et al. A natural allele of a transcription factor in rice confers broad-spectrum blast resistance. Cell 2017, 170, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Hummel, A.W.; Doyle, E.L.; Bogdanove, A.J. Addition of transcription activator-like effector binding sites to a pathogen strain-specific rice bacterial blight resistance gene makes it effective against additional strains and against bacterial leaf streak. New Phytol. 2012, 195, 883–893. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Tian, D.; Gu, K.; Zhou, Z.; Yang, X.; Luo, Y.; White, F.F.; Yin, Z. Genetic engineering of the Xa10 promoter for broad-spectrum and durable resistance to Xanthomonas oryzae pv. oryzae. Plant Biotechnol. J. 2015, 13, 993–1001. [Google Scholar] [CrossRef] [PubMed]
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).