1. Introduction
Oral mucositis is a significant complication of chemotherapy and/or radiation therapy, occurring in 80–100% of patients undergoing “high-risk” regimens and 40% of patients receiving standard dose chemotherapy [
1]. Initially, oral mucositis manifests as erythematous lesions 4–5 days following chemotherapy, progresses to erosive and ulcerative lesions within 7–10 days and persists for 2–4 weeks [
2]. Lesions associated with mucositis result in severe pain, difficulty in speech and swallowing, and a decreased quality of life for the patient. Furthermore, it predisposes the patient to a higher risk of local and systemic infection and interferes with the ability to deliver the intended dose of cancer therapy, thereby posing a threat to patient life, as well as causing an increased length of hospital stay for the patient [
3,
4]. 5-fluorouracil (5-FU) is a common chemotherapeutic agent which is widely used in the treatment of solid tumors [
5,
6]. 5-FU is an antimetabolite drug that acts as a phase-specific anti-pyrimidine by inhibiting DNA synthesis via inhibition of the enzyme thymidylate synthase [
7,
8]. Additional molecular mechanisms underlying the anticancer activities of 5-FU have been reported, such as via the inhibition of the mammalian target of rapamycin (mTOR)/ribosomal protein S6 kinase pathway activation [
9]. 5-FU was also found to inhibit cancer cell migration by upregulating Sestrin-2 [
10].
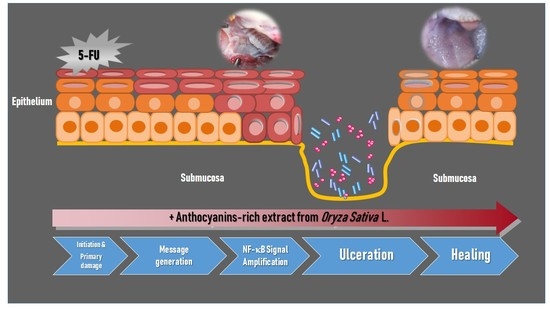
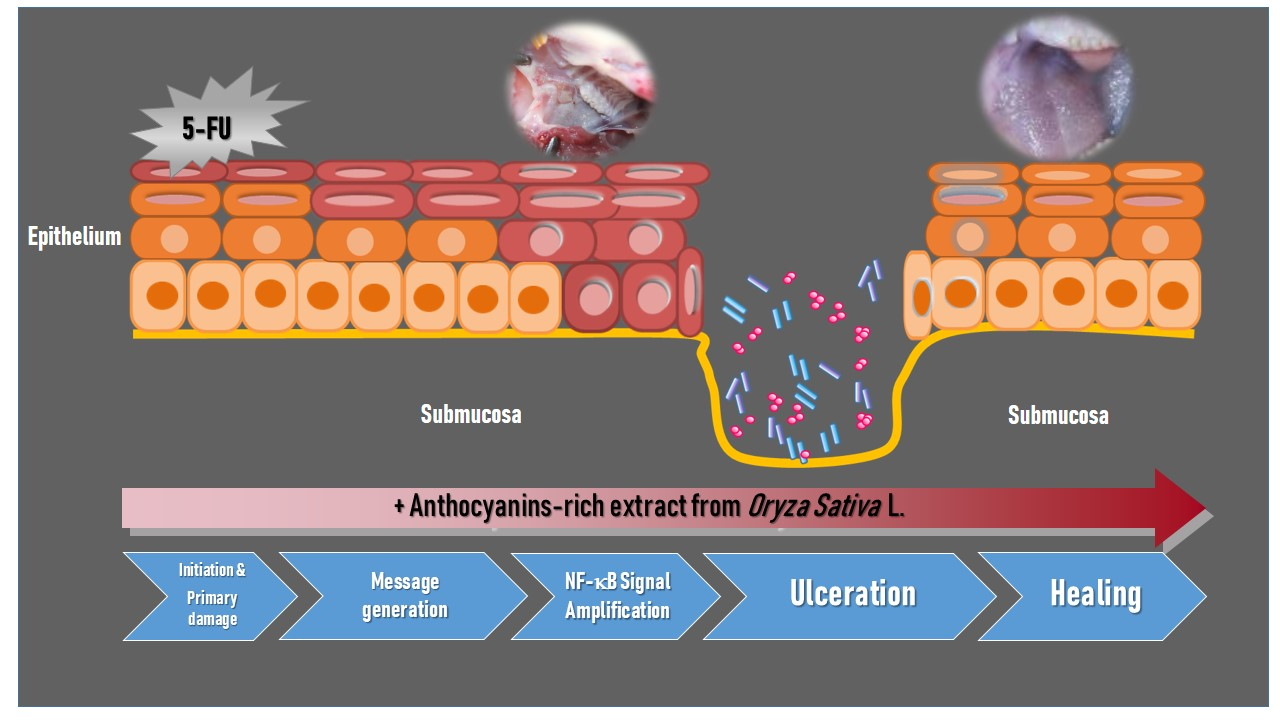
Systemic chemotherapy with 5-FU causes toxicity to the entire gastrointestinal tract. These cytotoxic effects are particularly evident in the oral and the oropharyngeal mucosa due to its rapid rate of epithelial cell turnover. Oral mucositis is not only a consequence of direct injury to the oral basal cells, but is also caused by a series of complex biological events based on five overlapping biological phases: initiation, message generation, signal amplification, ulceration, and healing [
11]. The mechanism of 5-FU-induced oral mucositis has been well identified. Likewise, the activation and translocation of transcription factor, nuclear factor-κB (NF-κB), from the cytosol to the nucleus is one of the key elements in the initiation of 5-FU-induced mucositis that leads to the upregulation of genes and the subsequent production of pro-inflammatory cytokines such as tumor necrosis factor-α (TNF-α), interleukin (IL)-1β, and IL-6 [
2,
11], cyclooxygenase (COX)-2 [
12], and high-mobility group box 1 protein (HMGB1) [
13].
The consumption of anthocyanins (ANTs) has a wide range of health-promoting properties [
14]. ANTs are members of the flavonoid group of phytochemicals that have been shown to have antioxidative properties and potential in therapeutic immunomodulation, leading to anti-cancer [
15,
16] and anti-inflammatory effects [
17,
18]. In vitro, ANTs from black soybean seed coats prevent excessive inflammation by blocking the translocation of NF-κB p65 into the nucleus in human dermal fibroblasts and keratinocytes [
17]. In an in vivo rat model of acute lung inflammation, ANT from blackberries was shown to reduce inflammatory parameters in a dose-dependent manner [
19]. Similarly, human models have revealed that ANT could suppress NF-κB activation in monocytes and reduce the plasma concentration of pro-inflammatory mediators [
18]. Recently, Tao, et al. reported that the administration of ANT from red cabbage (
Brassica oleracea L. var.
capitata L.) could prevent irinotecan-induced intestinal mucositis [
20]. Black rice (
Oryza sativa L.) has been widely cultivated and consumed in Southeast Asian countries for a long time. ANT in black rice was able to restore liver enzyme activity in rats with liver damage through its antioxidant properties [
21]. In addition, ANTs were also shown to inhibit breast cancer cell proliferation [
22]. Despite these findings, there have been no investigations into the effects of black rice-derived ANT in oral mucositis.
Although a variety of new approaches to oral mucositis have been employed, an efficacious intervention for prophylaxis or management of oral mucositis has not yet been identified. Based on our ANT study, and the critical role of NF-κB activation in the initiation of oral mucositis, the aim of this study was to evaluate the protective effects of ANT-rich extract found in Oryza sativa L. on oral wound healing, using an experimental rat model of 5-FU-induced mucositis, as well as attempting to elucidate possible transcription factor mechanisms.
3. Discussion
Studies on mucositis using animal models focus on continuing the search for new therapies for oral mucositis. The results from this study suggest that ANT-rich extract demonstrates significant anti-inflammatory properties and oral mucosa wound healing through the inhibition of NF-κB p50 and p65 signaling in 5-FU induced oral mucositis rat model and in the oral keratinocyte culture. To the best of our knowledge, this study demonstrates for the first time that ANT-rich extract from black rice (
Oryza sativa L.) is able to protect rats from post-chemotherapy oral mucositis and suppress the cell growth inhibitory effect of 5-FU. The results demonstrate a protective effect of ANT-rich extract on 5-FU induced oral mucositis, which was consistent with previous studies stating that ANT significantly lowered chemotherapy-induced intestinal mucositis [
20] and that ANT-rich extract from bilberry exhibited a protective effect against the myelotoxicity caused by 5-FU [
27].
Microorganisms inhabiting the oral cavity can enter the system through the ulcerative lesions seen in oral mucositis, consequently causing systemic infection in cancer patients receiving chemotherapy [
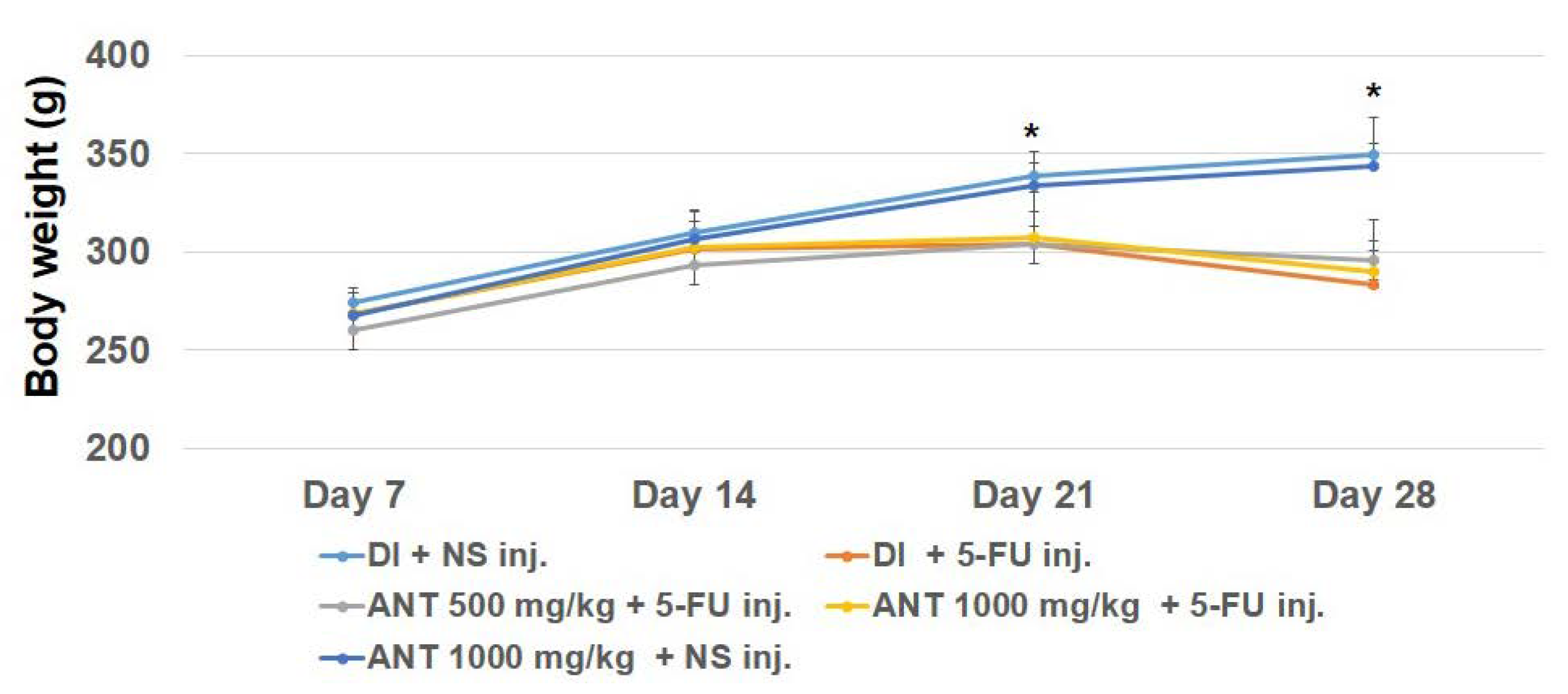
28]. Additionally, our findings revealed a significant loss of body weight in rats treated with 5-FU as early as day 14, suggesting a systemic repercussion of experimental oral mucositis, consistent with previous studies in a hamster model [
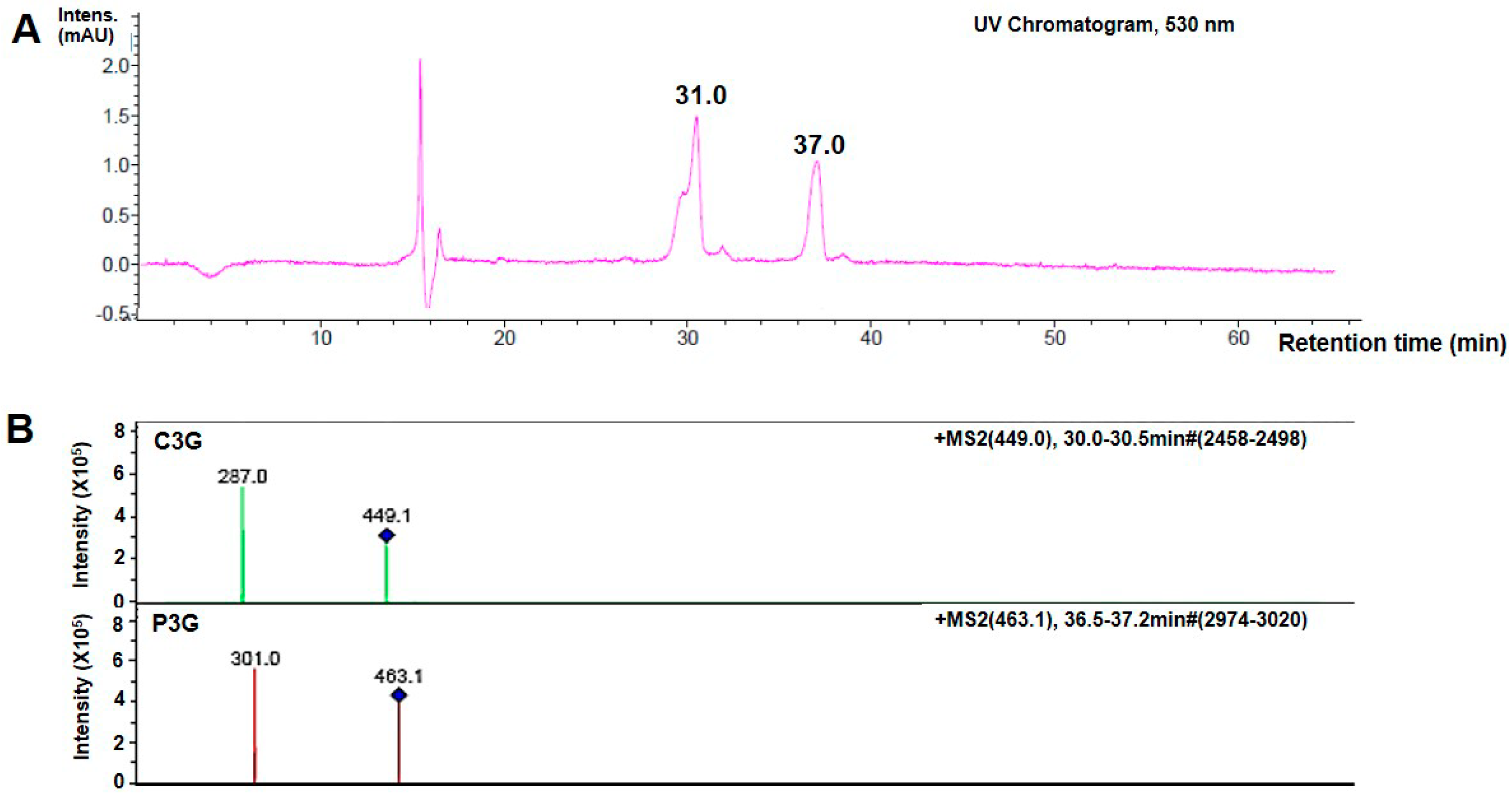
29]. However, gavage feeding with ANT did not cause any alterations in the average body weights of rats. In the present study, the chromatogram of purified products following ANT detection showed that C3G was the predominant ANT present in Thai
Oryza sativa L. The minor ANT detection was P3G. The ANT composition identified in our black rice extract was consistent with previous reporting [
30]. In our study, ANT was extracted in ethanol and contained C3G ≈ 0.85 mg/g and P3G ≈ 0.076 mg/g of the dry extract. The FRAP value and total phenolic content of the ANT-rich extract is 27.23 mM Fe (II) equivalents/gFW and 181.73 mM GAE/gFW, respectively. However, Sompong, et al. demonstrated tthe C3G (5.69 mg/g) and peonidin3-glucoside (11.46 mg/g) contents in black rice using HCl solvent in methanol (85:15, (
v/
v) [
31]. Their total phenolic contents values (300–600 mM GAE/gFW) were also higher than those of our extracts. The previous study on the ANT content of different rice varieties demonstrated that Thai red rice, Niaw Dam Pleuak Khao and Niaw Dam Pleuak Dam, followed by China black rice, has a superior quality with respect to its ANT compositions [
31]. The higher amount of ANT content and antioxidant capacities from the previous study compared to our findings are probably due to differences in rice cultivars, as well as varying methods of extraction, separation, purification, and analysis between the studies.
Previous study showed that rats receiving ANT-rich extract from
Oryza sativa L. Japonica at 500 mg/kg had an effect against liver damage by alcohol [
21]. Before beginning the current experiments, different concentrations (250, 500, 750, and 1000 mg/kg) of the same ANT extracts were tested. C3G is considered as a powerful antioxidant with pharmacological benefits including antioxidant properties [
21] and anti-inflammatory effects [
17,
18,
19]. We report for the first time that the highest ANT dose (1000 mg/kg body weight) from Thai black rice, which was gavage-fed once daily, contains approximately 0.85 mg/kg; C3G shows maximum protective and healing effects on oral mucositis induced by 5-FU. Apart from total phenolic and anthocyanin content, black rice contains high levels of flavonoids; more phytochemical analysis and the pharmaceutical activity of this plant should be further investigated.
5-FU is a common chemotherapeutic drug that causes oral mucositis [
4]. 5-FU injected rats have been demonstrated to be at a high risk of oral mucositis [
5,
32]. Recent studies have pinpointed NF-κB as having a key role in the pathogenesis of mucositis [
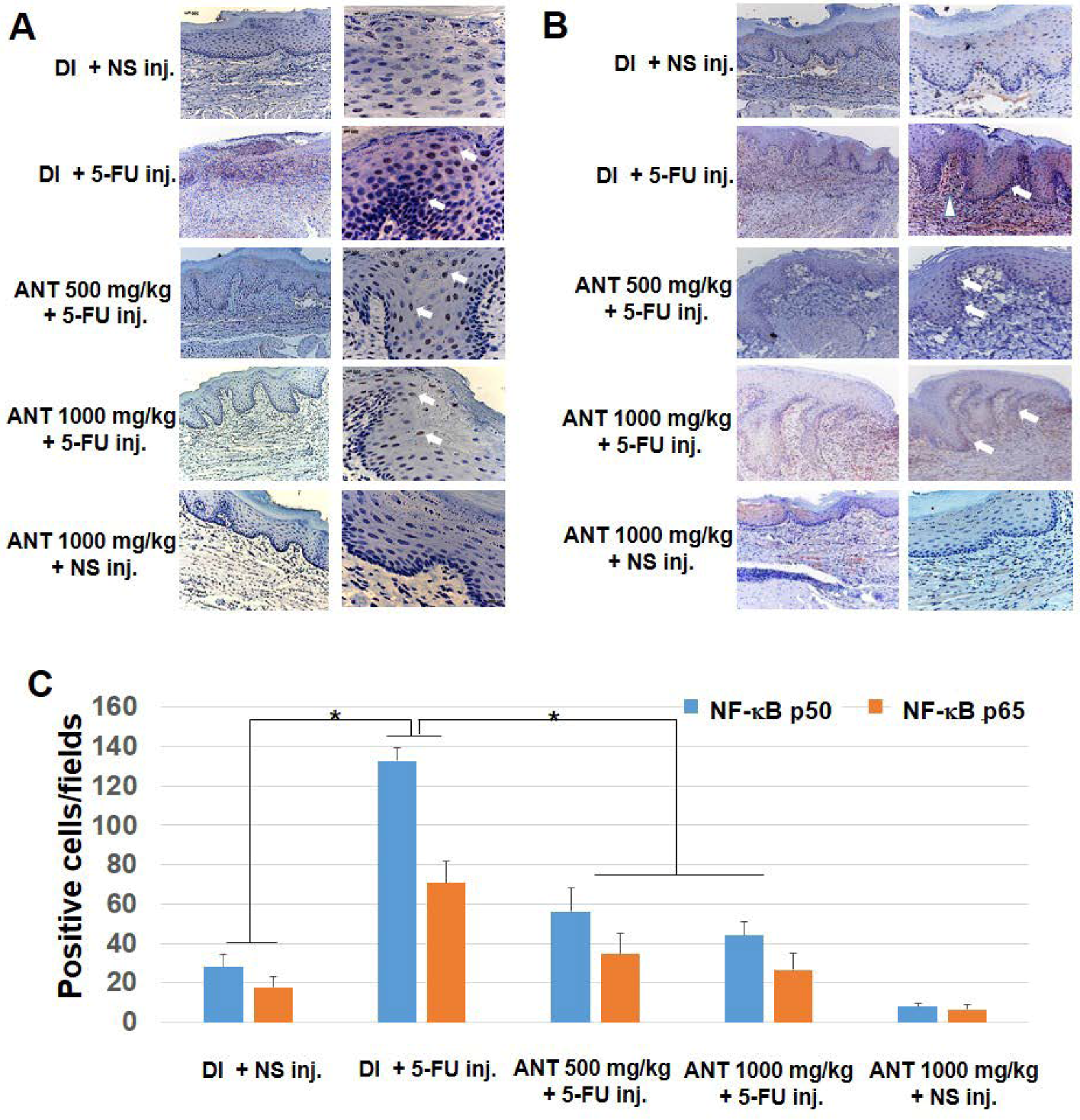
3,
33,
34]. In our study, we detected an abundance of activated NF-κB p50 and p65 in nuclear-stained oral keratinocytes obtained from rats injected with 5-FU. The exact mechanism(s) by which 5-FU causes oral mucositis are still unknown; however, 5-FU has been reported as an inducer of NF-κB [
24]. Therefore, the abundant amount of activated NF-κB p50 and p65 seen in oral keratinocytes and inflammatory cells in the granulation tissues of rats treated with 5-FU alone suggests the importance of NF-κB in the pathogenesis of 5-FU-induced oral mucositis. In agreement with our investigation, previous studies have shown strong associations between NF-κB-mediated biological manipulation of the oral mucosa and mucositis progression [
35].
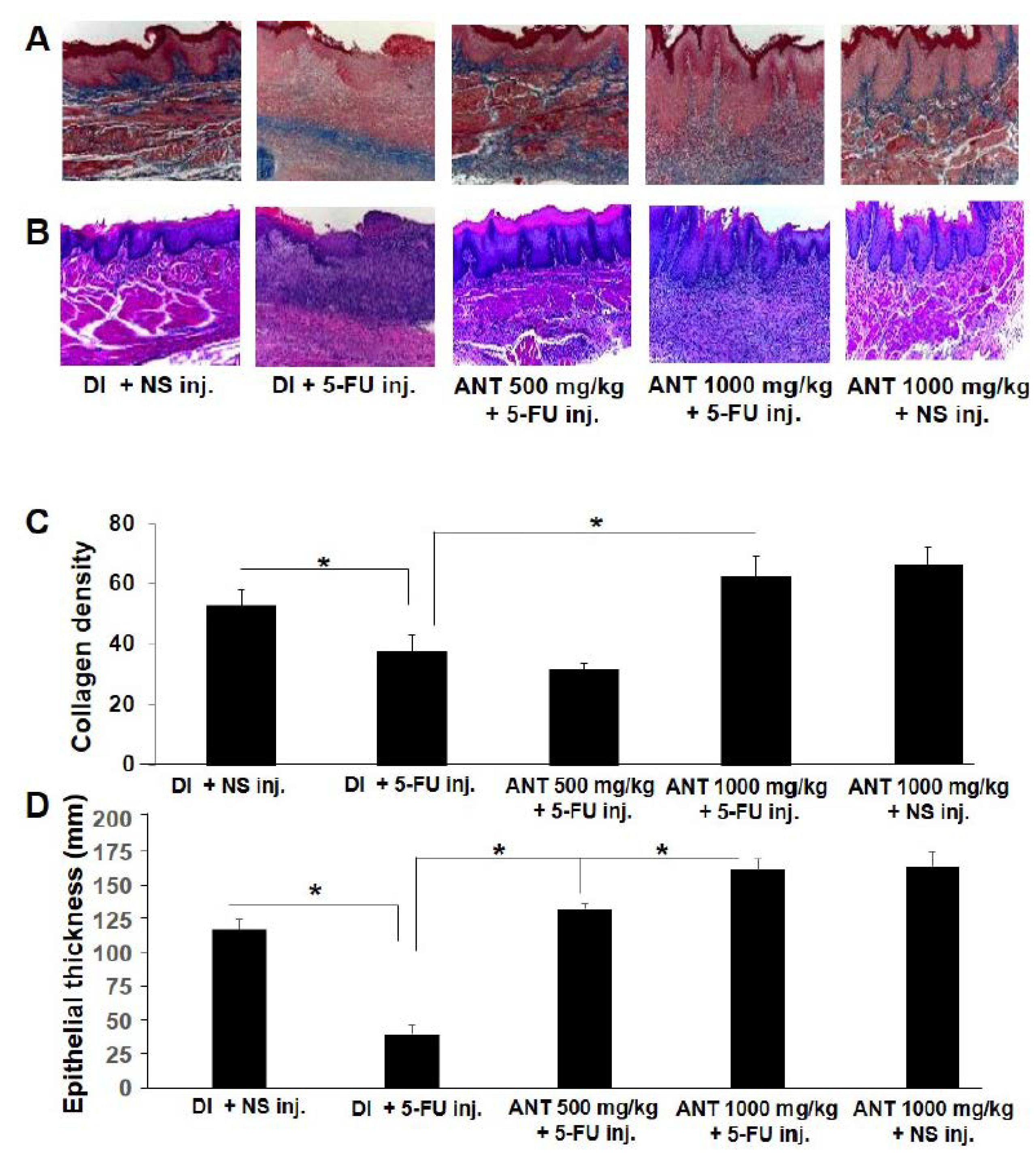
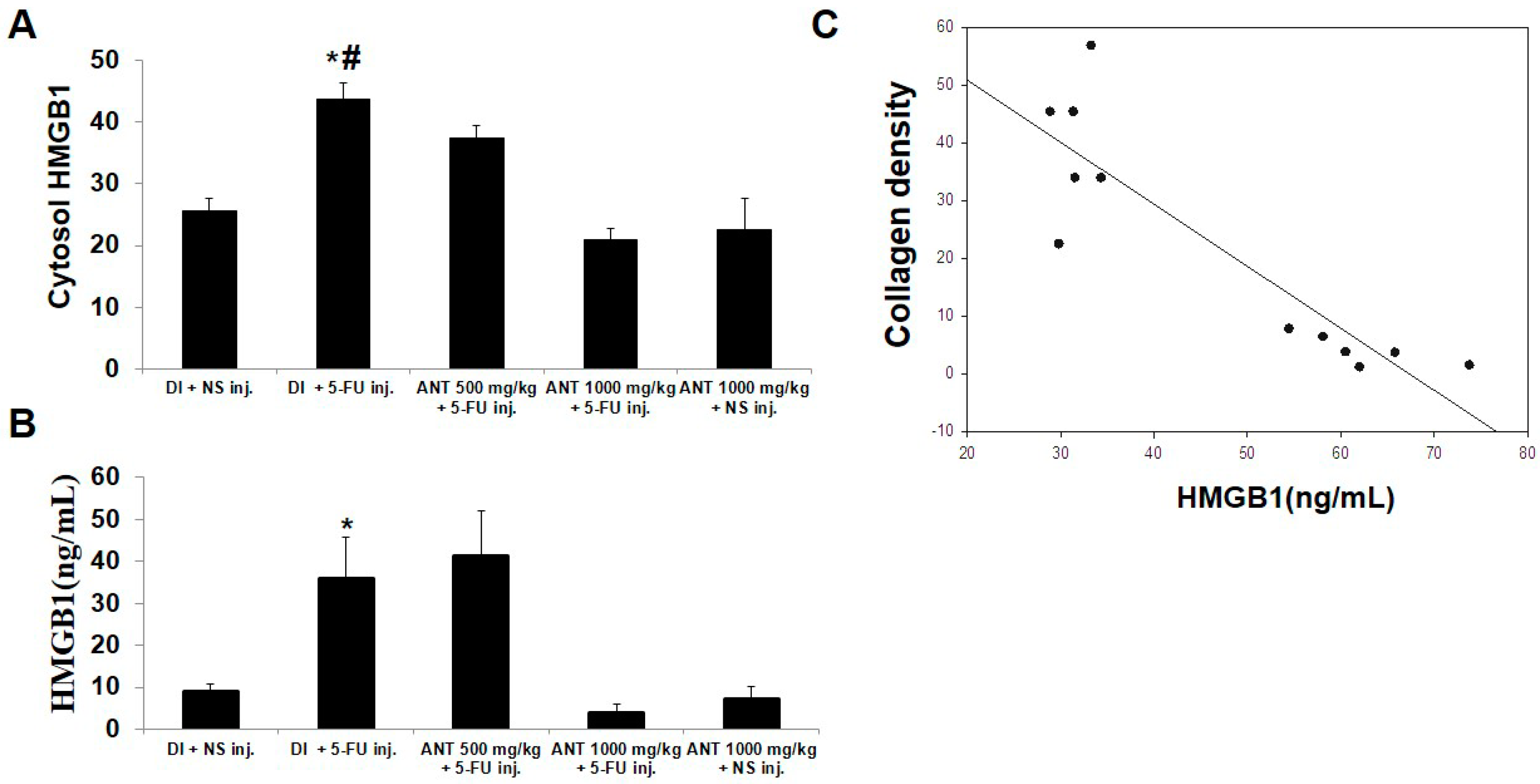
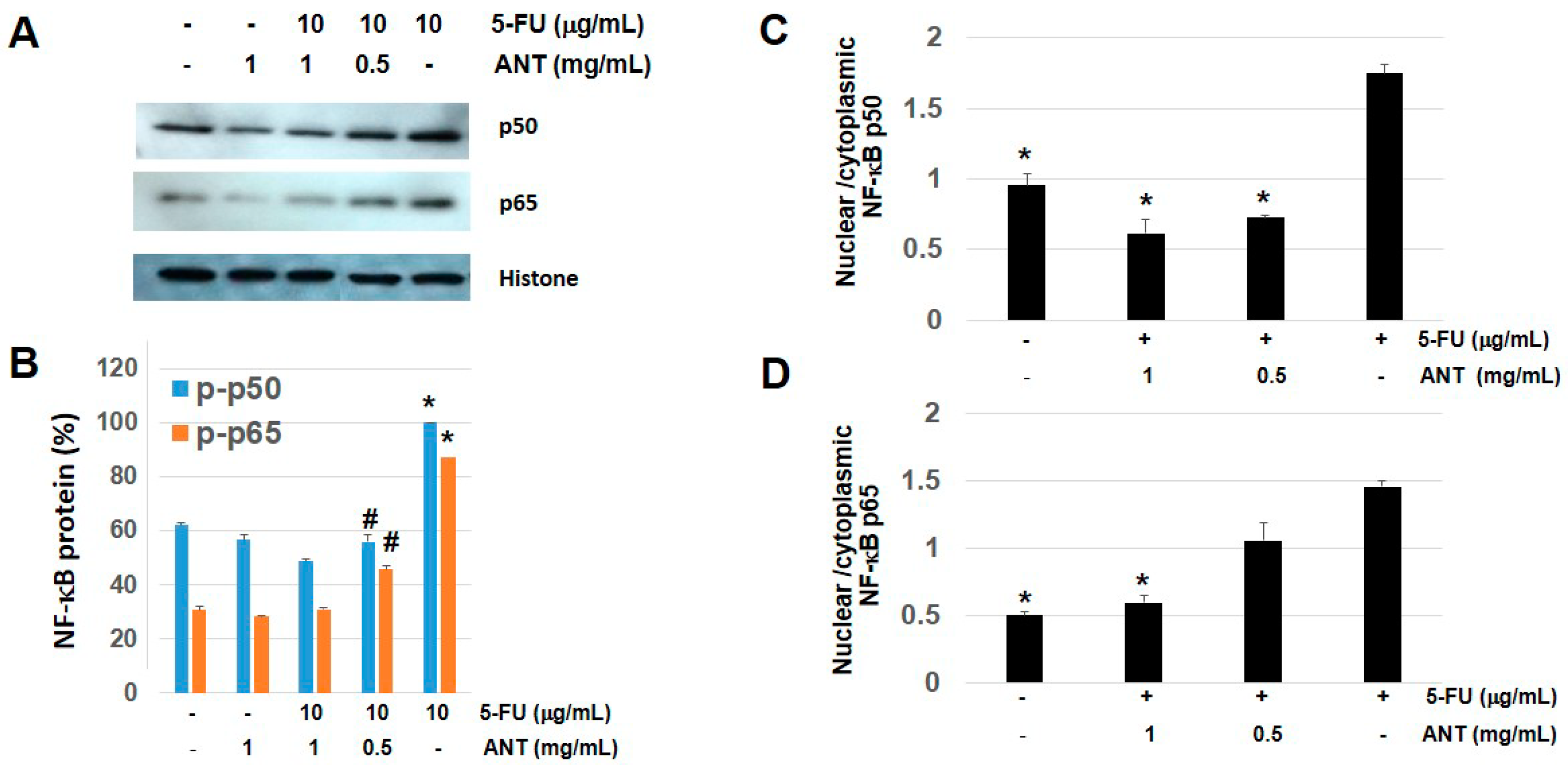
In the present investigation, we demonstrated that ANT-rich extract significantly reduced the lesions induced by 5-FU in the oral mucosa. The data provided in this study strongly suggest that ANT-rich extract given prior to chemotherapy favorably affects the severity and duration of ulcerative mucositis induced by 5-FU. The macroscopic and microscopic effects were associated with reduced inflammatory infiltration and epithelialization. Accordingly, these histopathologic findings were presented in a previous study [
27], although there were variations in the parameters used for biochemical analysis, plant species, and ANT content in the study mentioned. The results herein further verify the anti-inflammatory role of ANT on NF-κB and HMGB1 expression following 5-FU-treatment by immunohistochemical analysis and ELISA. Thus, the protective role of ANT found in the present study may be explained by its capacity to inhibit NF-κB p50 and p65 activation, thereby inhibiting the release of inflammatory cytokines that mediate 5-FU-induced oral mucositis. These findings are consistent with previous observations in animal and clinical studies, where ANT demonstrated anti-inflammatory effects by the inhibition of NF-κB transactivation [
17] and decreased plasma concentrations of pro-inflammatory mediators in human subjects [
18].
The primary damage of oral mucositis response describes an activation of transcription factors such as NF-κB leading to the production of pro-inflammatory cytokines such as TNF-α, IL-1β, and IL-6, which cause damage to both epithelium and connective tissue [
2,
11]. Our results demonstrate that the ANT-rich extract partially protects against 5-FU-induced cellular stasis in oral keratinocytes, and restores oral mucosal surfaces in vivo via immunological modulatory effects. Future studies should assess the predictive potential of ANT in determining associated cytokine production in vitro using peripheral blood mononuclear cells.
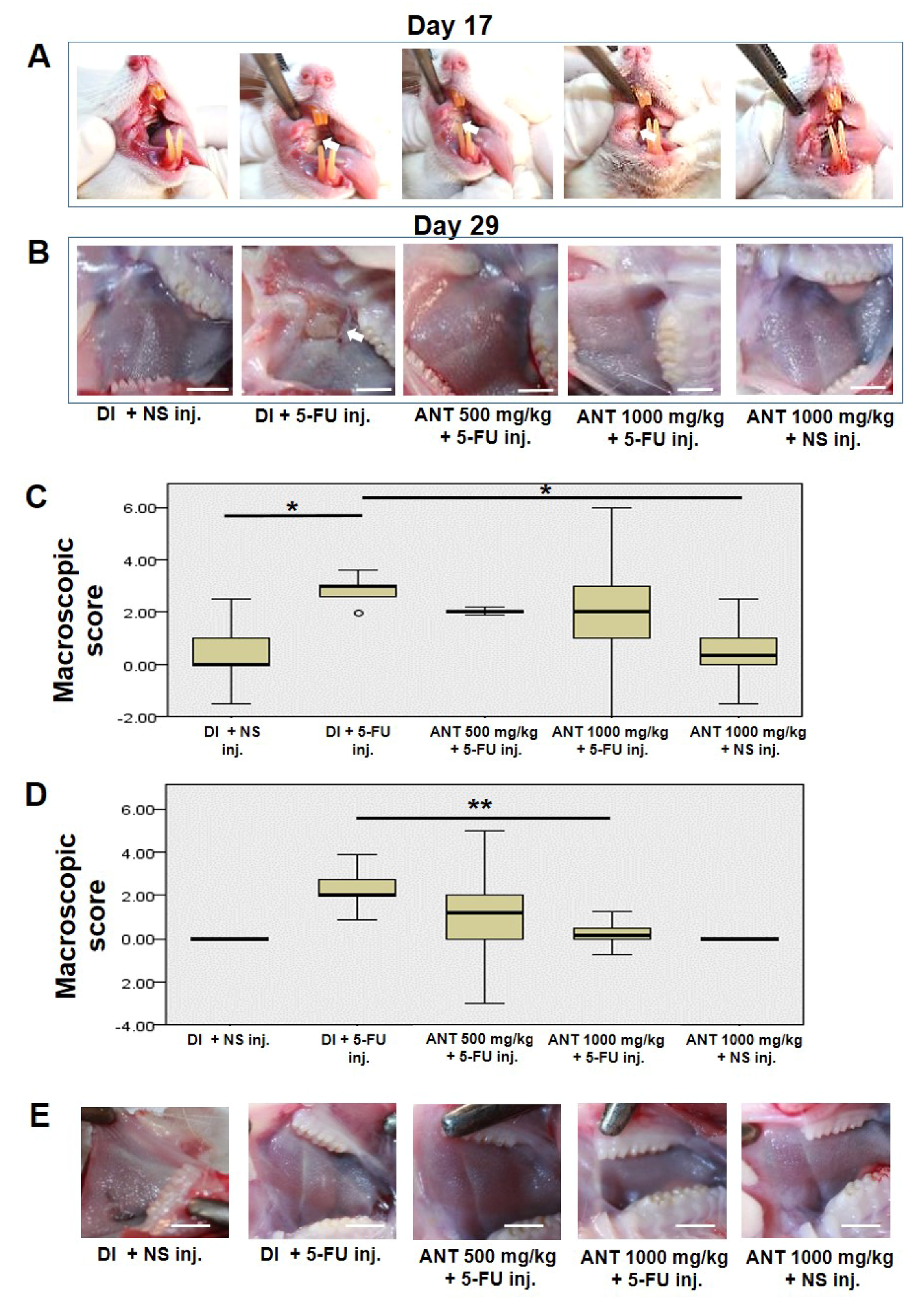
In the present study, erythema, hyperemia, hemorrhagic areas, and extensive ulcer and abscesses appeared in the cheek pouches of rats which were subjected to 5-FU injection by day 14, and their degree was maximal by day 17. Wound healing involves complex cellular and molecular interactions. There are five phases involved in the pathogenesis of mucositis, including initiation, primary damage and message generation, signal amplification, ulceration, and healing phase [
11]. Following the initiation of the healing cascade by direct injury to DNA, a series of transcription factors were prompted to respond to the primary damage, which triggered a consequence of cytokine release. In the final stage of the healing process, collagen deposition and remodeling occurs. Our findings demonstrated a slight decrease in the macroscopic scores of mucositis in rats which were treated with ANT-rich extract (groups C and D) compared to group B on day 17 of the experiment. However, a significant decrease in these scores (
p < 0.01) was observed on day 29 in rats which were treated with ANT-rich extract for 1000 mg/kg (group D) compared to group B. In addition, ANT-rich extract treatments attenuated NF-κB activation, suppressed the elevation of HMGB1 expression, and induced collagen formation. Thus, from our findings, ANT-rich extract treatments may not result in direct repair during the initiation stage of the wound healing but promote a permeability of collagen to the lesion, epithelial cell proliferation and differentiation, and repair the destructive mucosa during the late stage. Nevertheless, specific growth factors such as nerve growth factor [
36] or drugs such as corticosteroids [
37] which take part in the effective wound healing in the oral cavity should be employed to compare the efficacy with ANT-rich extract.
The present study is, as far as we know, the first investigation into the protective effects of ANT in 5-FU–induced oral mucositis and its involvement in protecting oral keratinocytes from the growth inhibitory effect of 5-FU. Mucositis can affect the entire mucosal lining of the gastrointestinal tract, with oral and oropharyngeal mucosae being common sites of disease occurrence [
38]. Our findings in vitro are consistent with animal studies demonstrating that the nuclear portions of NF-κB p50 and NF-κB p65 were activated by 5-FU, then suppressed following ANT-rich extract administration. However, the molecular mechanisms in other cells involved in oral mucositis such as oral fibroblasts should be further investigated [
39]. With these outcomes, further clinical studies on the anticancer activities of black rice extract should be performed.
In conclusion, our results highly suggest that ANT found in Thai Oryza sativa L. extracts possess anti-inflammatory and protective effects against experimental models of mucositis induced by 5-FU. We speculate that these anti-inflammatory effects are related to NF-κB suppression. The possibility of using this extract in the treatment of human oral mucositis merits further investigation.
4. Materials and Methods
4.1. Materials and Chemicals
NF-κB p50 and p65 Abs were purchased from Santa Cruz (Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA). HMGB1 Ab and cell fractionation kit were obstained from Abcam (Cambridge, MA, USA). Unless otherwise stated, all other reagents were supplied by Sigma-Aldrich Inc. (St. Louis, MO, USA).
4.2. Plant Material and Extraction
Black rice was obtained from Chachoengsao Province, Thailand. ANTs in Thai black rice were extracted in ethanol (60/40,
v/
v%) and concentrated at 40 °C (extraction yield 3.5% of dry weight) using a Buchi B-490 rotary evaporator (BÜCHI Labortechnik AG, Flawil, Switzerland), lyophilized with a freeze-dryer (Labconco Corp., Kansas City, MO, USA), and the crude extract was stored at 4 °C. Purified extracts were prepared following previous reporting [
40]. Briefly, a C18 Sep-Pak cartridge (Waters Corp., Milford, MO, US) was activated with DI and methanol (Merck KGaA, Darmstadt, Germany). The crude ANT extract was then loaded onto the column. Following successive washes, the ANTs were eluted with methanol containing 0.01% HCl. The ANT solution was then collected and condensed at 40 °C using a Büchi B-490 rotary evaporator under vacuum.
For HPLC-electrospray ionization (ESI) mass spectrometry (MS), ANT-rich extracts were separated and quantified by reverse-phase HPLC (Agilent model Model:1100 (Binary pump, Degasser, Autosample, DAD) using a Hypersil Gold C18 column (inner diameter, 3 μm; 4.6 × 150 mm; Thermo Fisher Scientific Inc., Waltham, MA, USA) following previous reporting [
40], with slight modifications. The column was eluted with a mobile phase consisting of formic acid (VWR International, Ltd., Leicestershire, UK) and methanol at a flow rate of 1 mL/min. The separated ANTs were detected and measured at 530 nm, and were identified based on their retention times and ultraviolet (UV)-visible (Vis) wavelength spectra of pure authentic standards (Sigma, St. Louis, MO, USA). The identity of each peak was verified. The chromatographic system was coupled to an ion trap mass spectrometer (Esqurire 3000+, Bruker Daltanic Model, Billerica, MA, USA) equipped with an ESI working in positive ion mode. The LC-MS was eluted with formic acid and methanol.
4.3. Quantification of ANT by UV-Vis Spectroscopy and Antioxidant Activities
The ANTs were quantified by UV-Vis spectroscopy as previously described [
14]. The model reaction solution was diluted with 0.01% HCl in distilled water. Absorbance at 510 nm was compared with that of known standard solutions using a Genesys 10 UV spectrophotometer (Thermo Scientific, Grand Island, NY, USA). The determination of total phenolic content was determined using Folin–Ciocalteu reagent (FRC) as previously described [
40], with minor modifications. The absorbance of the mixture was measured at 765 nm using a UV-Vis Genesys 10 UV spectrophotometer. A standard curve was plotted using gallic acid (0.07–10 mg/mL in methanol; Sigma) as a standard. The total phenolic content was expressed as gallic acid equivalents (mM GAE/gFW). Ferric-reducing antioxidant power (FRAP) was measured as per previous reporting [
40]. The absorption was measured at 595 nm using a spectrophotometer (Epoch; Biotek, Winooski, VT, USA) with the Gen 5 Data analysis software interface (
https://www.biotek.com/). Aqueous or methanol solutions containing known Fe (II) concentrations were used to calibrate the FRAP assay. FRAP values, expressed as mM of Fe(II) equivalents/gFW), were determined by comparing the change in absorption of the test mixture with those of the Fe(II) standards. Total antioxidant activity was determined using the OxiSelect TAC Assay kit (Cell Biolabs, San Diego, CA, USA) following manufacturer’s instructions.
4.4. Animal Model
Healthy male Wistar rats (7 weeks, 280–300 g) from the National Laboratory Animal Center, Mahidol University, Nakhon Pathom, Thailand were used in this study. Animals were maintained in an animal room with a standard environmental condition of 22 ± 2 °C, 50 ± 5% relative humidity, a 12 h/12 h light/dark cycle with water and food ad libitum. All animal experiments were performed according to the guidelines for animal care and use of Mahidol University Animal Care and Use Committee. All animal experiments were approved by the Ethical Clearance Institutional Animal Care and Use Committee (Faculty of Pharmacy, Mahidol University, Bangkok, Thailand) in 8 May 2014 (certificate No: PYT011/2557).
4.5. Induction of Experimental Oral Mucositis
Oral mucositis was induced following the protocol proposed by Sonis et al [
32]. Three intraperitoneal (i.p.) injections of 5-FU 60 mg/kg were administered once a day at a five-day interval (days 7, 12, 17) for 29 days. On day 11, the right cheek pouch of all animals was irritated by superficial scratching with a tip of an 18-gauge needle under isoflurane anesthesia to mimic the clinical effect of chronic irritation. The needle was dragged twice in a linear fashion across the everted right cheek pouch. This technique has been used repeatedly to induce ulcerative mucositis, which is similar to human oral mucositis.
4.6. Animals and Study Design
Wistar rats were randomly divided into five groups, as follows: group A: gavage-fed 1 mL deionized water (DI) and injected normal saline (NS) by i.p. injection, serving as a negative control group; group B: gavage-fed 1 mL (DI) and injected with 60 mg/kg 5-FU by i.p. injection, serving as a positive control group; group C: gavage-fed 500 mg/kg ANT-rich extracts once daily and injected with 60 mg/kg 5-FU i.p.; group D: gavage-fed 1000 mg/kg ANT-rich extracts once daily and injected with 60 mg/kg 5-FU i.p.; and group E: gavage-fed 1000 mg/kg ANT-rich extracts once daily and injected with NS i.p. The beginning of the gavage was considered to be day 1 of the study. ANT-rich extracts were fed 1 week before the first dose of 5-FU injection. The animals were observed daily and weighed once every two days. For the purpose of observation, rats were anesthetized with isoflurane and the buccal pouches were everted and photographed. Subsequently, buccal mucosa was biopsied and tissue samples were collected for histochemical analysis and immunohistochemistry for p50 and p65 subunits of NF-κB, and HMGB1. Blood was collected for the assessment of serum HMGB1 level.
4.7. Macroscopic Analysis of Buccal Mucosae
On day 29, the animals were anesthetized with isoflurane inhalation, and the buccal pouches were photographed for macroscopic scoring based on a valid macroscopic scoring system described by Lima et al. [
41]. For macroscopic analysis, inflammatory aspects such as erythema, hyperemia, hemorrhagic areas, epithelial ulcerations, and abscesses were evaluated by three examiners and graded as follows: score 0: normal cheek pouch with erythema and hyperemia absent or discrete, no hemorrhagic areas, ulcerations, or abscess; score 1: moderate erythema and hyperemia, no hemorrhagic areas, ulceration, or abscess; score 2: severe erythema and hyperemia, presence of hemorrhagic areas, small ulceration, or scarred tissue, but no abscess; and score 3: severe erythema and hyperemia, presence of hemorrhagic areas, extensive ulcerations, and abscess of the buccal mucosa.
4.8. Histopathological Analysis
In each experimental group, the samples of buccal pouches were removed by incisional biopsy for histopathological analysis. The specimens were fixed in 10% neutral buffered formalin, dehydrated, and embedded in paraffin. Sections of 6 μm thickness were obtained. Mallory’s azan staining was used to visualize collagen fibers, and Mayer’s hematoxylin stain was used to examine the parameters of inflammatory cell infiltration, vasodilation, presence of hemorrhagic areas, edema, ulcerations, and abscesses [
42]. Histopathological sections were analyzed under light microscopy. Density of collagen bundles and epithelial thickness were quantified using Image J software (ImageJ, US National Institutes of Health, Bethesda, MD, USA) from six randomly selected areas of the same section under high power fields (×400). The average density of collagen was calculated as Integrated Density (the product of Area and Mean Gray Value)/Area [
43]. Calculations of the average epithelial thickness in the Mayer’s hematoxylin staining samples were based on 20 measurements for each time point.
4.9. Immunohistochemistry for p50 and p65 NF-κB and HMGB1
Sections were deparaffinized by xylene and rehydrated. Immunohistochemistry was performed as described previously for NF-κB p50, p65 and HMGB1 [
20]. Sections were incubated with their respective primary Abs diluted in PBS containing 0.1% bovine serum albumin overnight at 4 °C. Monocolonal NF-κB p50, p65 and HMGB1 Abs were used at a dilution of 1:500. Following washing with PBS, the sections were incubated with corresponding HRP-conjugated secondary Abs for 1 h at RT, followed by staining with diaminobenzidine (DAB) (Histofine MAX-PO kit, Nicherei, Tokyo, Japan). Control staining was performed with non-immune IgGs and slides stained with DAB (DakoCytomation; Glostrup, Denmark) and counterstained for 2 min with hematoxylin (DakoCytomation). Sample was visualized by light microscopy (LEICA Microsystem GmbH, Wetzlar, Germany). A total of four digital images per group/time point under high-power fields (×400) were assessed by a calibrated examiner blinded to the protocol. A modified semi-quantitative method was used to count the NF-κB p50, p65 and HMGB1 positive cells as previously described [
12].
4.10. Cell Culture Conditions
Oral keratinocytes were obtained from ScienCell Research Laboratories (San Diego, CA, USA). Cells were maintained in complete oral keratinocyte medium supplemented with keratinocyte growth supplement and penicillin/streptomycin solution. Cells were maintained at 37 °C in a humidified atmosphere containing 5% CO2 and 95% air. All cells were cultured in serum-free Opti-MEM-I medium (Gibco, Grand Island, NY, USA) for at least 15 h before treatment to eliminate the possible side effect of growth factors.
4.11. Cell Viability Test
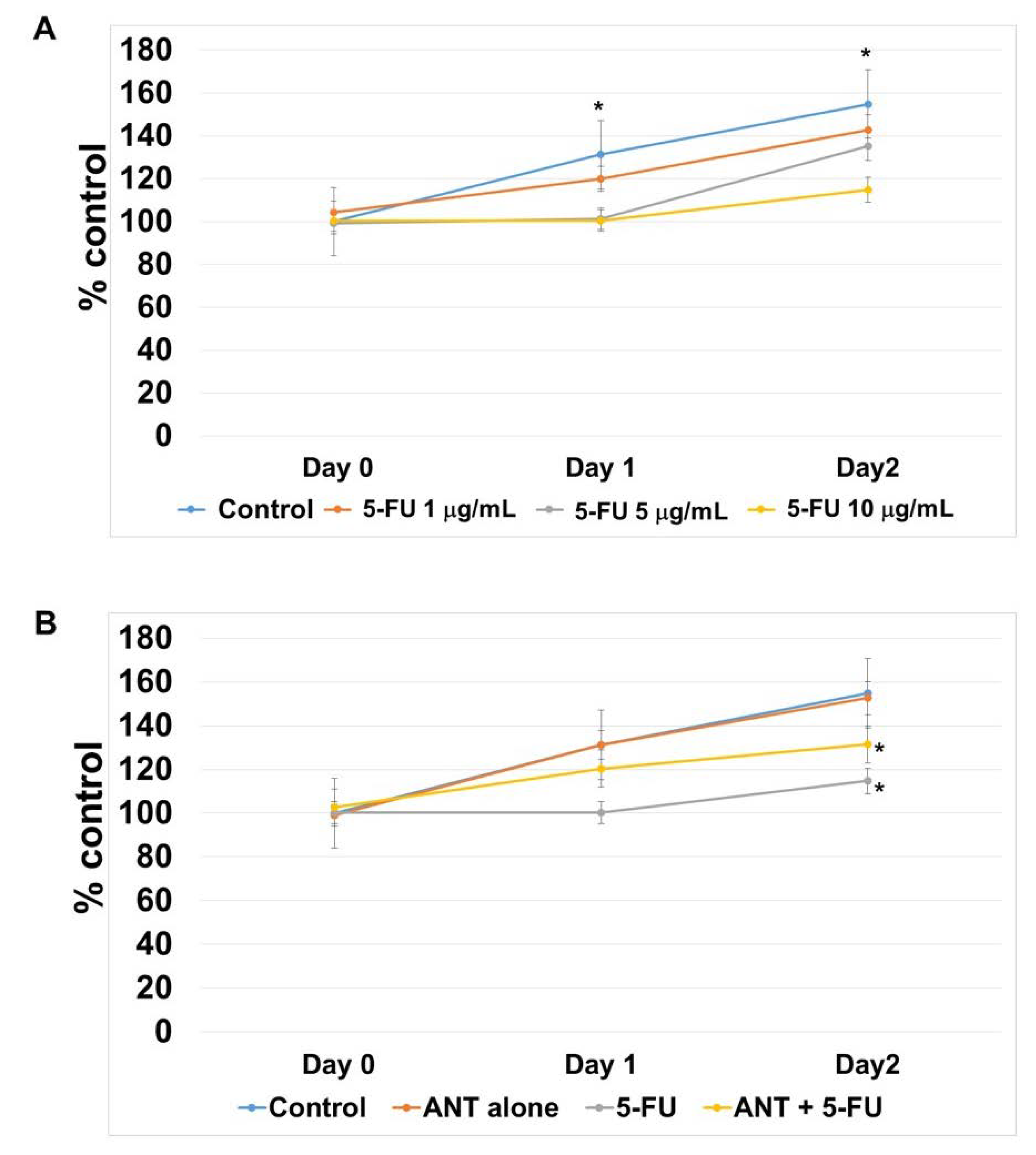
Cell viability was performed by MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) assay according to a method previously described [
42]. Briefly, cells were treated with ANT-rich extracts for 1 mg/mL and 5-FU for 1, 5, and 10 μg/mL for 2 days and incubated with MTT (0.5 mg/mL) for 3 h. Formazan crystal was solubilized by adding DMSO for 16 h.
4.12. Preparation of Nucleic/Cytosolic Fractions
Cells (2 × 105 cells) were cultured in 60 mm dish and were pretreated with ANT-rich extracts (0.5 and 1 mg/mL) and exposed to 10 g/mL of 5-FU for 1 h. Whole cell lysates were extracted using cell lysis buffer (Cell Signaling Technology, Inc., Danvers, MA, USA) or nucleic/cytosolic fractions were extracted using cell fractionation kit according to the manufacturer’s instructions and stored at −20 °C until use. Protein concentrations were determined by Bradford protein assay using bovine serum albumin as standard (Bio-Rad, Hercules, CA, USA).
4.13. Western Blot Analysis
Western blot analyses were carried out as described previously [
44]. Briefly, samples were mixed with 2× electrophoresis sample buffer solution with bromophenol blue (Santa Cruz Biotechnology) before being subjected to 12% sodium dodecyl sulfate -polyacrylamide gel electrophoresis (PAGE) and transferred onto nitrocellulose membranes (Schleicher & Schuell, Dassel, Germany). To prevent nonspecific binding, the membrane was blocked with a solution containing 5% (
w/
v) nonfat dry milk with 1% (
v/
v) Tween 20 in PBS for 1 h at RT. Anti-NF-κB p50 or p65 Ab was incubated for 1 h at RT. The membranes were washed and incubated with horseradish peroxidase-conjugated anti-mouse monoclonal IgG (MP Biomedicals Inc., Solon, OH, USA). Labeled bands were visualized using an enhanced chemiluminescence system (GE Healthcare Bio-Science, Pittsburgh, PA, USA) and exposed to high-performance chemiluminescence film (GE Healthcare). The intensity of the protein bands in Western blotting was quantified using National Institutes of Health Image version 1.63 software (National Institutes of Health, Bethesda, MO, USA).
4.14. HMGB1 Measurement by Enzyme-Linked Immunosorbent Assay
HMGB1 levels were quantified using a commercial ELISA kit (Shino-test, Sagamihara, Kanagawa, Japan), according to the manufacturer’s instruction.
4.15. Statistical Analysis
Statistical significances between different groups were determined by one-way analysis of variance (ANOVA) test or Student’s paired t-test. The correlation among three examiners and the macroscopic scores were calculated using Pearson’s correlation coefficient. p values < 0.05 were considered statistically significant. All calculations were performed using SPSS software (version 20.0; SPSS Inc., Chicago, IL, USA).

 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}