Is Osteopontin a Friend or Foe of Cell Apoptosis in Inflammatory Gastrointestinal and Liver Diseases?



Abstract
:1. Introduction
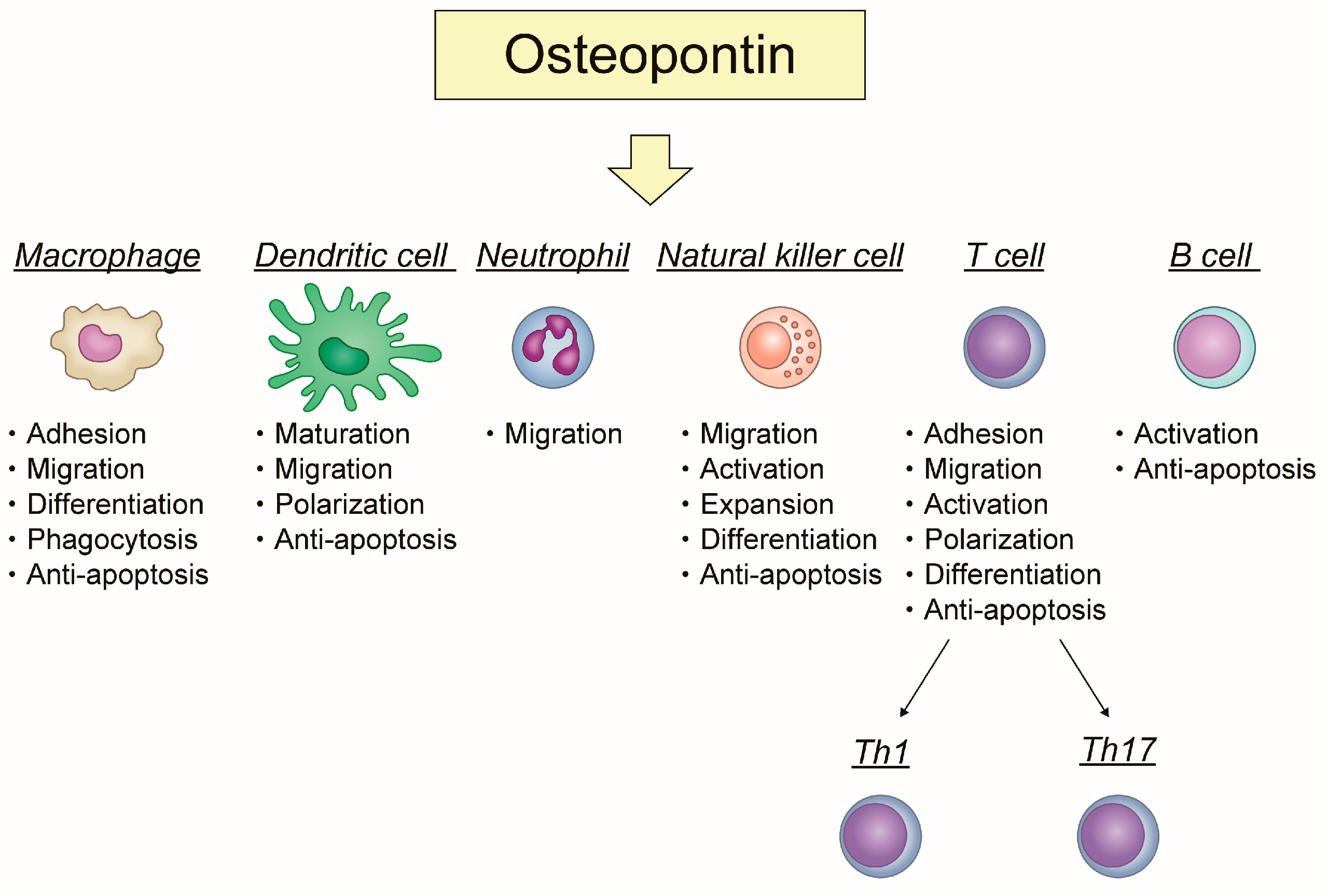
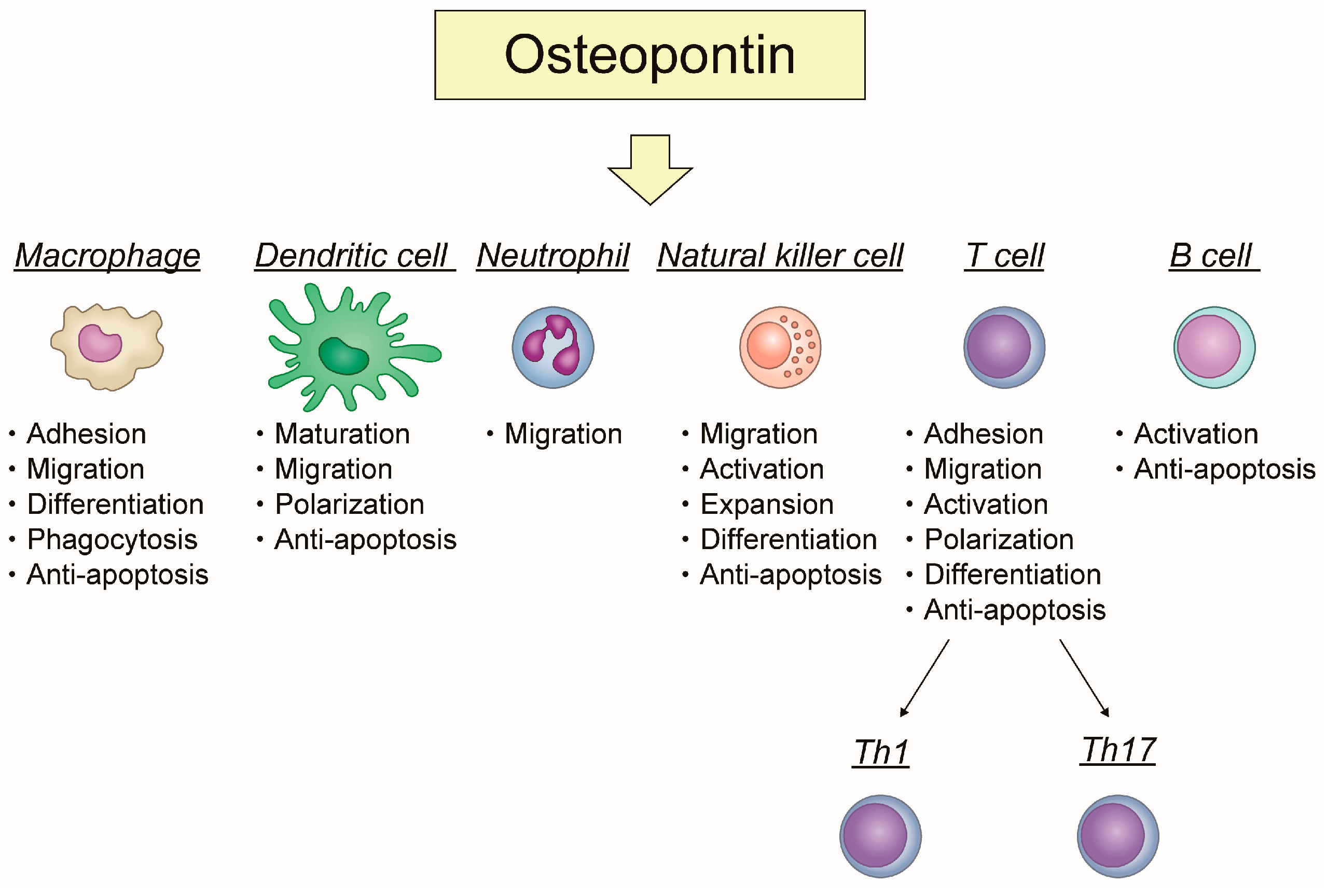
2. The Role of OPN in Immune Cells, Focusing on Its Association with Inflammation and Apoptosis
2.1. Macrophage
2.2. Dendritic Cell (DC)
2.3. Neutrophil
2.4. Natural Killer (NK) Cell
2.5. T Cell
2.6. B Cell
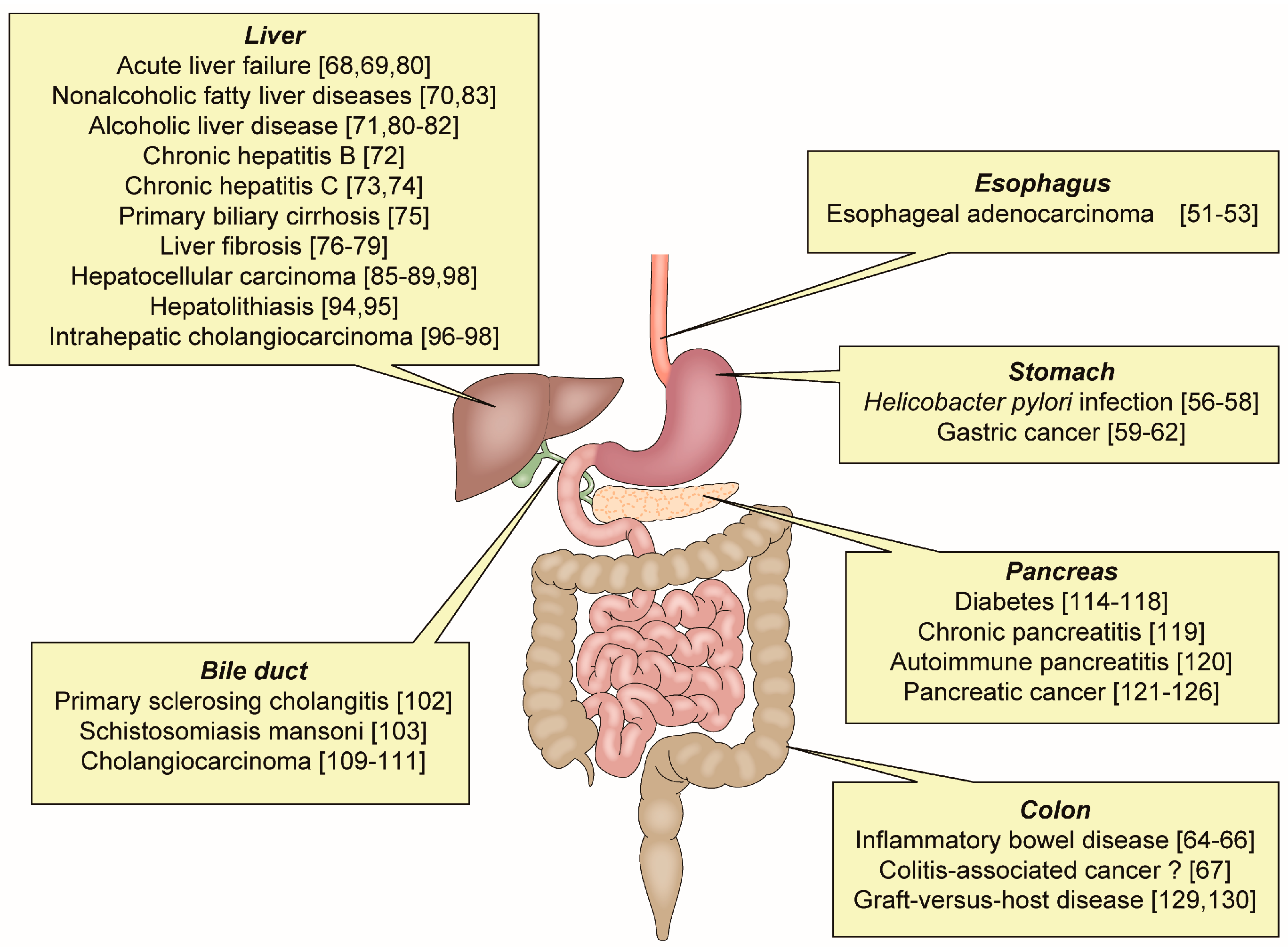
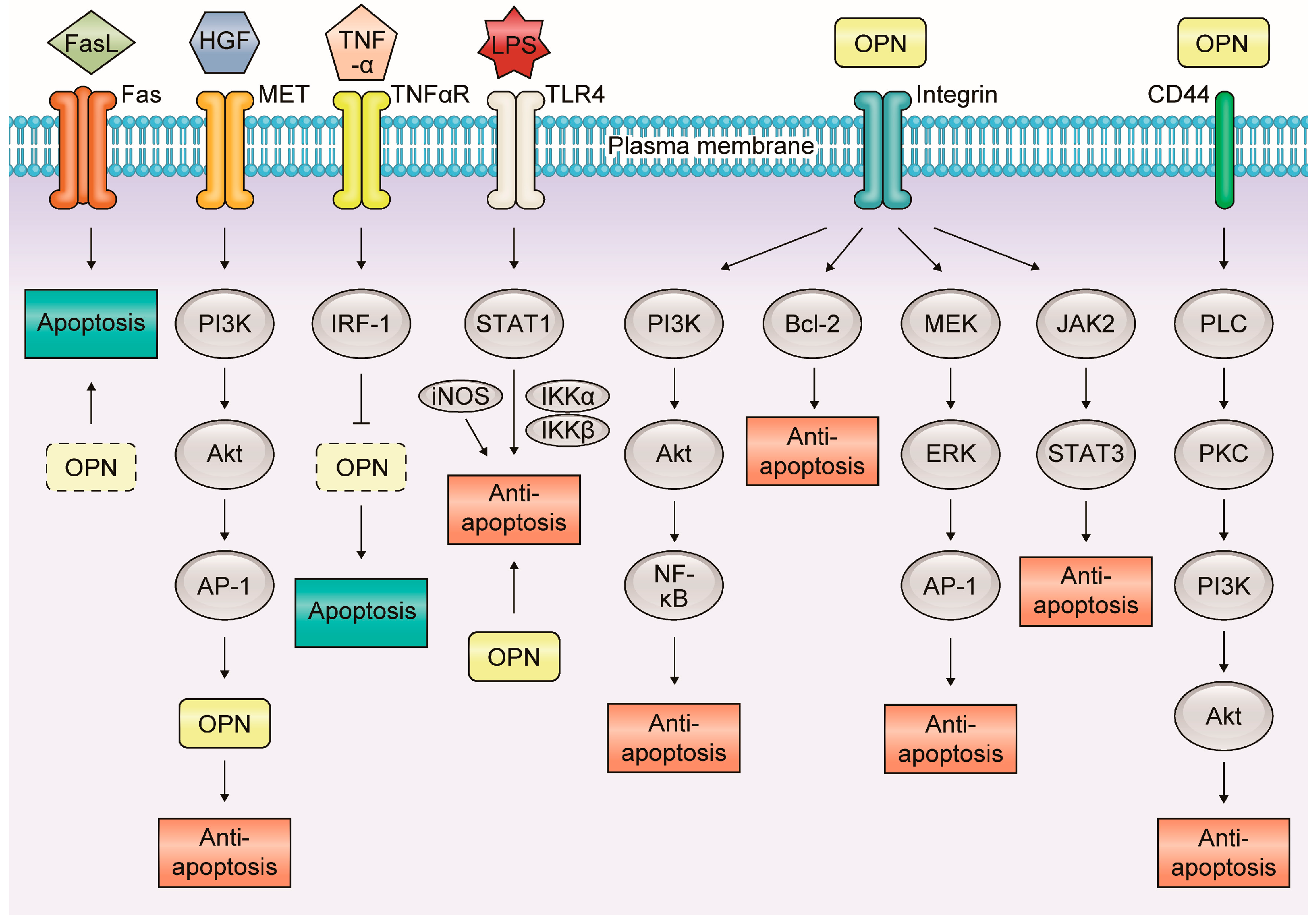
3. The Association of OPN with Apoptosis in Inflammatory GI and Liver Diseases
3.1. Esophageal Adenocarcinoma (EAC)
3.2. Helicobacter pylori Infection and Gastric Cancer (GC)
3.3. Inflammatory Bowel Disease (IBD) and Colitis-Associated Cancer (CAC)
3.4. Liver Diseases
3.5. Bile Duct Diseases
3.6. Pancreatic Diseases
3.7. Graft-Versus-Host Disease (GVHD)
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Global Action against Cancer-Updated Version; World Health Organization and International Union against Cancer: Geneva, Switzerland, 2005; p. 12.
- Galdiero, M.R.; Bonavita, E.; Barajon, I.; Garlanda, C.; Mantovani, A.; Jaillon, S. Tumor associated macrophages and neutrophils in cancer. Immunobiology 2013, 218, 1402–1410. [Google Scholar] [CrossRef] [PubMed]
- Ruffel, B.; Coussens, L.M. Macrophages and therapeutic resistance in cancer. Cancer Cell 2015, 27, 462–472. [Google Scholar] [CrossRef] [PubMed]
- Shalapour, S.; Karin, M. Immunity, inflammation, and cancer: An eternal fight between good and evil. J. Clin. Investig. 2015, 125, 3347–3355. [Google Scholar] [CrossRef] [PubMed]
- Grivennikov, S.I.; Greten, F.R.; Karin, M. Immunity, inflammation, and cancer. Cell 2010, 140, 883–899. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.Y.; Sneddon, J.B.; Alizadeh, A.A.; Sood, R.; West, R.B.; Montgomery, K.; Chi, J.T.; van de Rijn, M.; Botstein, D.; Brown, P.O. Gene expression signature of fibroblast serum response predicts human cancer progression: Similarities between tumors and wounds. PLoS Biol. 2004, 2, E7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erez, N.; Truitt, M.; Olson, P.; Arron, S.T.; Hanahan, D. Cancer-associated fibroblasts are activated in incipient neoplasia to orchestrate tumor-promoting inflammation in an NF-κB-dependent manner. Cancer Cell 2010, 17, 135–147. [Google Scholar] [CrossRef] [PubMed]
- Franzen, A.; Heinegard, D. Isolation and characterization of two sialoproteins present only in bone calcified matrix. Biochem. J. 1985, 232, 715–724. [Google Scholar] [CrossRef] [PubMed]
- Vaschetto, R.; Nicola, S.; Olivieri, C.; Boggio, E.; Piccolella, F.; Mesturini, R.; Damnotti, F.; Colombo, D.; Navalesi, P.; Della Corte, F.; et al. Serum levels of osteopontin are increased in SIRS and sepsis. Intensive Care Med. 2008, 34, 2176–2184. [Google Scholar] [CrossRef] [PubMed]
- Castello, L.M.; Raineri, D.; Salmi, L.; Clemente, N.; Vaschetto, R.; Quaglia, M.; Garzaro, M.; Gentilli, S.; Navalesi, P.; Cantaluppi, V.; et al. Osteopontin at the Crossroads of Inflammation and Tumor Progression. Mediat. Inflamm. 2017, 2017, 4049098. [Google Scholar] [CrossRef] [PubMed]
- Fisher, L.W.; Torchia, D.A.; Fohr, B.; Young, M.F.; Fedarko, N.S. Flexible structures of SIBLING proteins, bone sialoprotein, and osteopontin. Biochem. Biophys. Res. Commun. 2007, 280, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Kazanecki, C.C.; Uzwiak, D.J.; Denhardt, D.T. Control of osteopontin signaling and function by post-translational phosphorylation and protein folding. J. Cell. Biochem. 2007, 102, 912–924. [Google Scholar] [CrossRef] [PubMed]
- Shevde, L.A.; Samant, R.S. Role of osteopontin in the pathophysiology of cancer. Matrix Biol. 2014, 37, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Denhardt, D.T.; Noda, M.; O’Regan, A.W.; Pavlin, D.; Berman, J.S. Osteopontin as a means to cope with environmental insults: Regulation of inflammation, tissue remodeling, and cell survival. J. Clin. Investig. 2001, 107, 1055–1061. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, M.L.; Kim, H.J.; Kim, J.H.; Garcia, V.A.; Cantor, H. Alternative translation of osteopontin generates intracellular and secreted isoforms that mediate distinct biological activities in dendritic cells. Proc. Natl. Acad. Sci. USA 2008, 105, 7235–7239. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.X.; Denhardt, D.T. Osteopontin: Role in immuneregulation and stress responses. Cytokine Growth Factor Rev. 2008, 19, 333–345. [Google Scholar] [CrossRef] [PubMed]
- Ramaiah, S.K.; Rittling, S. Pathophysiological role of osteopontin in hepatic inflammation, toxicity, and cancer. Toxicol. Sci. 2008, 103, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Rittling, S.R.; Chambers, A.F. Role of osteopontin in tumour progression. Br. J. Cancer 2004, 90, 1877–1881. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.F.; Berse, B.; Van de Water, L.; Papadopoulos-Sergiou, A.; Perruzzi, C.A.; Manseau, E.J.; Dvorak, H.F.; Senger, D.R. Expression and distribution of osteopontin in human tissues: Widespread association with luminal epithelial surfaces. Mol. Biol. Cell 1992, 3, 1169–1180. [Google Scholar] [CrossRef] [PubMed]
- Ashkar, S.; Weber, G.F.; Panoutsakopoulou, V.; Sanchirico, M.E.; Jansson, M.; Zawaideh, S.; Rittling, S.R.; Denhardt, D.T.; Glimcher, M.J.; Cantor, H. Eta-1 (osteopontin): An early component of type-1 (cell-mediated) immunity. Science 2000, 287, 860–864. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Guo, M.; Chen, J.H.; Wang, Z.; Du, X.F.; Liu, P.X.; Li, W.H. Osteopontin knockdown inhibits αv, β3 integrin-induced cell migration and invasion and promotes apoptosis of breast cancer cells by inducing autophagy and inactivating the PI3K/Akt/mTOR pathway. Cell Physiol. Biochem. 2014, 33, 991–1002. [Google Scholar] [CrossRef] [PubMed]
- Liaw, L.; Skinner, M.P.; Raines, E.W.; Ross, R.; Cheresh, D.A.; Schwartz, S.M.; Giachelli, C.M. The adhesive and migratory effects of osteopontin are mediated via distinct cell surface integrins. Role of alpha v beta 3 in smooth muscle cell migration to osteopontin in vitro. J. Clin. Investig. 1995, 95, 713–724. [Google Scholar] [CrossRef] [PubMed]
- Yokosaki, Y.; Tanaka, K.; Higashikawa, F.; Yamashita, K.; Eboshida, A. Distinct structural requirements for binding of the integrins αvβ6, αvβ3, αvβ5, α5β1 and α9β1 to osteopontin. Matrix Biol. 2005, 24, 418–427. [Google Scholar] [CrossRef] [PubMed]
- Green, P.M.; Ludbrook, S.B.; Miller, D.D.; Horgan, C.M.; Barry, S.T. Structural elements of the osteopontin SVVYGLR motif important for the interaction with α(4) integrins. FEBS Lett. 2001, 503, 75–79. [Google Scholar] [CrossRef]
- Marcondes, M.C.; Poling, M.; Watry, D.D.; Hall, D.; Fox, H.S. In vivo osteopontin-induced macrophage accumulation is dependent on CD44 expression. Cell. Immunol. 2008, 254, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Lund, S.A.; Wilson, C.L.; Raines, E.W.; Tang, J.; Giachelli, C.M.; Scatena, M. Osteopontin mediates macrophage chemotaxis via α4 and α9 integrins and survival via the α4 integrin. J. Cell. Biochem. 2013, 114, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Das, R.; Philip, S.; Mahabeleshwar, G.H.; Bulbule, A.; Kundu, G.C. Osteopontin: It’s role in regulation of cell motility and nuclear factor κB-mediated urokinase type plasminogen activator expression. IUBMB Life 2005, 57, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Viatour, P.; Merville, M.P.; Bours, V.; Chariot, A. Phosphorylation of NF-κB and IκB proteins: Implications in cancer and inflammation. Trends Biochem. Sci. 2005, 30, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Renkl, A.C.; Wussler, J.; Ahrens, T.; Thoma, K.; Kon, S.; Uede, T.; Martin, S.F.; Simon, J.C.; Weiss, J.M. Osteopontin functionally activates dendritic cells and induces their differentiation toward a Th1-polarizing phenotype. Blood 2005, 106, 946–955. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, K.; Iyonaga, K.; Ichiyasu, H.; Nagano, J.; Suga, M.; Sasaki, Y. Differentiation, maturation, and survival of dendritic cells by osteopontin regulation. Clin. Diagn. Lab. Immunol. 2005, 12, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Weiss, J.M.; Renkl, A.C.; Maier, C.S.; Kimmig, M.; Liaw, L.; Ahrens, T.; Kon, S.; Maeda, M.; Hotta, H.; Uede, T.; et al. Osteopontin is involved in the initiation of cutaneous contact hypersensitivity by inducing Langerhans and dendritic cell migration to lymph nodes. J. Exp. Med. 2001, 194, 1219–1229. [Google Scholar] [CrossRef] [PubMed]
- Hirano, Y.; Azi, M.; Yang, W.L.; Wang, Z.; Zhou, M.; Ochani, M.; Khader, A.; Wang, P. Neutralization of osteopontin attenuates neutrophil migration in sepsis-induced acute lung injury. Crit. Care 2015, 19, 53. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.; da Silva, A.P.; Bansal, A.K.; Bansal, M.; Sun, C.; Lee, H.; Glogauer, M.; Sodek, J.; Zohar, R. Role of osteopontin in neutrophil function. Immunology 2007, 122, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Hui, T.; Matsui, A.; Allahem, Z.; Johnston, C.D.; Ruiz-Torruella, M.; Rittling, S.R. Modulation of infection-mediated migration of neutrophils and CXCR2 trafficking by osteopontin. Immunology 2017, 150, 74–86. [Google Scholar] [CrossRef] [PubMed]
- Leavenworth, J.W.; Verbinnen, B.; Wang, Q.; Shen, E.; Cantor, H. Intracellular osteopontin regulates homeostasis and function of natural killer cells. Proc. Natl. Acad. Sci. USA 2015, 112, 494–499. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.X.; Shek, K.; Wang, S.; Huang, X.; Lau, A.; Yin, Z.; Sun, H.; Liu, W.; Garcia, B.; Rittling, S.; et al. Osteopontin expressed in tubular epithelial cells regulates NK cell-mediated kidney ischemia reperfusion injury. J. Immunol. 2010, 185, 967–973. [Google Scholar] [CrossRef] [PubMed]
- Cen, C.; Aziz, M.; Yang, W.L.; Nicastro, J.M.; Coppa, G.F.; Wang, P. Osteopontin Blockade Attenuates Renal Injury After Ischemia Reperfusion by Inhibiting NK Cell Infiltration. Shock 2017, 47, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; He, C.; Jing, W.; Zhou, X.; Chen, R.; Cao, L.; Zhu, M.; Jia, R.; Wang, H.; Guo, Y.; et al. Intracellular osteopontin inhibits toll-like receptor signaling and impedes liver carcinogenesis. Cancer Res. 2015, 75, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Leavenworth, J.W.; Verbinnen, B.; Yin, J.; Huang, H.; Cantor, H. A p85α-osteopontin axis couples the receptor ICOS to sustained Bcl-6 expression by follicular helper and regulatory T cells. Nat. Immunol. 2015, 16, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Fan, K.; Dai, J.; Wang, H.; Wei, H.; Cao, Z.; Hou, S.; Qian, W.; Li, B.; Zhao, J.; Xu, H.; et al. Treatment of collagen-induced arthritis with an anti-osteopontin monoclonal antibody through promotion of apoptosis of both murine and human activated T cells. Arthritis Rheum. 2008, 58, 2041–2052. [Google Scholar] [CrossRef] [PubMed]
- Lampe, M.A.; Patarca, R.; Iregui, M.V.; Cantor, H. Polyclonal B cell activation by the Eta-1 cytokine and the development of systemic autoimmune disease. J. Immunol. 1991, 147, 2902–2906. [Google Scholar] [PubMed]
- Iizuka, J.; Katagiri, Y.; Tada, N.; Murakami, M.; Ikeda, T.; Sato, M.; Hirokawa, K.; Okada, S.; Hatano, M.; Tokuhisa, T.; et al. Introduction of an osteopontin gene confers the increase in B1 cell population and the production of anti-DNA autoantibodies. Lab. Investig. 1998, 78, 1523–1533. [Google Scholar] [PubMed]
- Kitagori, K.; Yoshifuji, H.; Oku, T.; Sasaki, C.; Miyata, H.; Mori, K.P.; Nakajima, T.; Ohmura, K.; Kawabata, D.; Yukawa, N.; et al. Cleaved Form of Osteopontin in Urine as a Clinical Marker of Lupus Nephritis. PLoS ONE 2016, 11, e0167141. [Google Scholar] [CrossRef] [PubMed]
- Clemente, N.; Raineri, D.; Cappellano, G.; Boggio, E.; Favero, F.; Soluri, M.F.; Dianzani, C.; Comi, C.; Dianzani, U.; Chiocchetti, A. Osteopontin Bridging Innate and Adaptive Immunity in Autoimmune Diseases. J. Immunol. Res. 2016, 2016, 7675437. [Google Scholar] [CrossRef] [PubMed]
- Clemente, N.; Comi, C.; Raineri, D.; Cappellano, G.; Vecchio, D.; Orilieri, E.; Gigliotti, C.L.; Boggio, E.; Dianzani, C.; Sorosina, M.; et al. Role of Anti-Osteopontin Antibodies in Multiple Sclerosis and Experimental Autoimmune Encephalomyelitis. Front. Immunol. 2017, 8, 321. [Google Scholar] [CrossRef] [PubMed]
- Dianzani, C.; Bellavista, E.; Liepe, J.; Verderio, C.; Martucci, M.; Santoro, A.; Chiocchetti, A.; Gigliotti, C.L.; Boggio, E.; Ferrara, B.; et al. Extracellular proteasome-osteopontin circuit regulates cell migration with implications in multiple sclerosis. Sci. Rep. 2017, 7, 43718. [Google Scholar] [CrossRef] [PubMed]
- Ma, N.; He, Y.; Xiao, H.; Han, G.; Chen, G.; Wang, Y.; Wang, K.; Hou, C.; Yang, X.; Marrero, B.; et al. BAFF maintains T-cell survival by inducing OPN expression in B cells. Mol. Immunol. 2014, 57, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Chiche, J.; Rouleau, M.; Gounon, P.; Brahimi-Horn, M.C.; Pouyssegur, J.; Mazure, N.M. Hypoxic enlarged mitochondria protect cancer cells from apoptotic stimuli. J. Cell. Physiol. 2010, 222, 648–657. [Google Scholar] [CrossRef] [PubMed]
- Lagergren, J.; Bergstrom, R.; Lindgren, A.; Nyren, O. Symptomatic gastroesophageal reflux as a risk factor for esophageal adenocarcinoma. N. Engl. J. Med. 1999, 340, 825–831. [Google Scholar] [CrossRef] [PubMed]
- Shaheen, N.J.; Richter, J.E. Barrett’s oesophagus. Lancet 2009, 373, 850–861. [Google Scholar] [CrossRef]
- Kim, S.M.; Park, Y.Y.; Park, E.S.; Cho, J.Y.; Izzo, J.G.; Zhang, D.; Kim, S.B.; Lee, J.H.; Bhutani, M.S.; Swisher, S.G.; et al. Prognostic biomarkers for esophageal adenocarcinoma identified by analysis of tumor transcriptome. PLoS ONE 2010, 5, e15074. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Myers, A.L.; Wang, Z.; Nancarrow, D.J.; Ferrer-Torres, D.; Handlogten, A.; Leverenz, K.; Bao, J.; Thomas, D.G.; Wang, T.D.; et al. Osteopontin (OPN/SPP1) isoforms collectively enhance tumor cell invasion and dissemination in esophageal adenocarcinoma. Oncotarget 2015, 6, 22239–22257. [Google Scholar] [CrossRef] [PubMed]
- Miller, C.T.; Lin, L.; Casper, A.M.; Lim, J.; Thomas, D.G.; Orringer, M.B.; Chang, A.C.; Chambers, A.F.; Giordano, T.J.; Glover, T.W. Genomic amplification of MET with boundaries within fragile site FRA7G and upregulation of MET pathways in esophageal adenocarcinoma. Oncogene 2006, 25, 409418. [Google Scholar] [CrossRef] [PubMed]
- Correa, P.; Haenszel, W.; Cuello, C.; Zavala, D.; Fontham, E.; Zarama, G.; Tannenbaum, S.; Collazos, T.; Ruiz, B. Gastric precancerous process in a high risk population: Cohort follow-up. Cancer Res. 1990, 50, 4737–4740. [Google Scholar] [PubMed]
- Kinoshita, H.; Hayakawa, Y.; Koike, K. Metaplasia in the Stomach-Precursor of Gastric Cancer? Int. J. Mol. Sci. 2017, 18, 2063. [Google Scholar] [CrossRef] [PubMed]
- Rollo, E.E.; Laskin, D.L.; Denhardt, D.T. Osteopontin inhibits nitric oxide production and cytotoxicity by activated RAW264.7 macrophages. J. Leukoc. Biol. 1996, 60, 397–404. [Google Scholar] [PubMed]
- Chang, W.L.; Yang, H.B.; Cheng, H.C.; Chuang, C.H.; Lu, P.J.; Sheu, B.S. Increased gastric osteopontin expression by Helicobacter pylori Infection can correlate with more severe gastric inflammation and intestinal metaplasia. Helicobacter 2011, 16, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.L.; Lin, M.Y.; Kuo, H.Y.; Yang, H.B.; Cheng, H.C.; Lu, C.C.; Sheu, B.S. Osteopontin polymorphism increases gastric precancerous intestinal metaplasia susceptibility in Helicobacter pylori infected male. Future Oncol. 2017, 13, 1415–1425. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Ming, Y.; Mao, Y.; Bao, S.; Ouyang, G. Osteopontin prevents curcumin-induced apoptosis and promotes survival through Akt activation via αvβ3 integrins in human gastric cancer cells. Exp. Biol. Med. (Maywood) 2008, 233, 1537–1545. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.L.; Wang, M.J.; Sudhir, P.R.; Chen, G.D.; Chi, C.W.; Chen, J.Y. Osteopontin promotes integrin activation through outsidein and inside-out mechanisms: OPN-CD44V interaction enhances survival in gastrointestinal cancer cells. Cancer Res. 2007, 67, 2089–2097. [Google Scholar] [CrossRef] [PubMed]
- Gong, M.; Lu, Z.; Fang, G.; Bi, J.; Xue, X. A small interfering RNA targeting osteopontin as gastric cancer therapeutics. Cancer Lett. 2008, 272, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Park, J.W.; Go, D.M.; Kim, H.K.; Kwon, H.J.; Han, S.U.; Kim, D.Y. Ablation of osteopontin suppresses N-methyl-N-nitrosourea and Helicobacter pylori-induced gastric cancer development in mice. Carcinogenesis 2015, 36, 1550–1560. [Google Scholar] [PubMed]
- Tang, R.; Yang, G.; Zhang, S.; Wu, C.; Chen, M. Opposite effects of interferon regulatory factor 1 and osteopontin on the apoptosis of epithelial cells induced by TNF-α in inflammatory bowel disease. Inflamm. Bowel Dis. 2014, 20, 1950–1961. [Google Scholar] [CrossRef] [PubMed]
- Toyonaga, T.; Nakase, H.; Ueno, S.; Matsuura, M.; Yoshino, T.; Honzawa, Y.; Itou, A.; Namba, K.; Minami, N.; Yamada, S.; et al. Osteopontin Deficiency Accelerates Spontaneous Colitis in Mice with Disrupted Gut Microbiota and Macrophage Phagocytic Activity. PLoS ONE 2015, 10, e0135552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heilmann, K.; Hoffmann, U.; Witte, E.; Loddenkemper, C.; Sina, C.; Schreiber, S.; Hayford, C.; Holzlöhner, P.; Wolk, K.; Tchatchou, E.; et al. Osteopontin as two-sided mediator of intestinal inflammation. J. Cell. Mol. Med. 2009, 13, 1162–1174. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Eckhardt, E.R.; Oz, H.S.; Bruemmer, D.; de Villiers, W.J. Osteopontin deficiency protects mice from Dextran sodium sulfate-induced colitis. Inflamm. Bowel Dis. 2006, 12, 790–796. [Google Scholar] [CrossRef] [PubMed]
- Mannick, E.E.; Cote, R.L.; Schurr, J.R.; Krowicka, H.S.; Sloop, G.D.; Zapata-Velandia, A.; Correa, H.; Ruiz, B.; Horswell, R.; Lentz, J.J. Altered phenotype of dextran sulfate sodium colitis in interferon regulatory factor-1 knock-out mice. J. Gastroenterol. Hepatol. 2005, 20, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Arai, M.; Yokosuka, O.; Kanda, T.; Fukai, K.; Imazeki, F.; Muramatsu, M.; Seki, N.; Miyazaki, M.; Ochiai, T.; Hirasawa, H.; et al. Serum osteopontin levels in patients with acute liver dysfunction. Scand. J. Gastroenterol. 2006, 41, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Srungaram, P.; Rule, J.A.; Yuan, H.J.; Reimold, A.; Dahl, B.; Sanders, C.; Lee, W.M.; Acute Liver Failure Study Group. Plasma osteopontin in acute liver failure. Cytokine 2015, 73, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, Y.; Ozturk, O.; Alahdab, Y.O.; Senates, E.; Colak, Y.; Doganay, H.L.; Coskunpinar, E.; Oltulu, Y.M.; Eren, F.; Atug, O.; et al. Serum osteopontin levels as a predictor of portal inflammation in patients with nonalcoholic fatty liver disease. Dig. Liver Dis. 2013, 45, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Seth, D.; Leo, M.A.; McGuinness, P.H.; Lieber, C.S.; Brennan, Y.; Williams, R.; Wang, X.M.; McCaughan, G.W.; Gorrell, M.D.; Haber, P.S. Gene expression profiling of alcoholic liver disease in the baboon (Papio hamadryas) and human liver. Am. J. Pathol. 2003, 163, 2303–2317. [Google Scholar] [CrossRef]
- Zhao, L.; Li, T.; Wang, Y.; Pan, Y.; Ning, H.; Hui, X.; Xie, H.; Wang, J.; Han, Y.; Liu, Z.; et al. Elevated plasma osteopontin level is predictive of cirrhosis in patients with hepatitis B infection. Int. J. Clin. Pract. 2008, 62, 1056–1062. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Zhu, G.; Huang, M.; Lou, G.; Liu, Y.; Wang, S. Plasma osteopontin concentration correlates with the severity of hepatic fibrosis and inflammation in HCV-infected subjects. Clin. Chim. Acta 2010, 411, 675–678. [Google Scholar] [CrossRef] [PubMed]
- Matsue, Y.; Tsutsumi, M.; Hayashi, N.; Saito, T.; Tsuchishima, M.; Toshikuni, N.; Arisawa, T.; George, J. Serum osteopontin predicts degree of hepatic fibrosis and serves as a biomarker in patients with hepatitis C virus infection. PLoS ONE 2015, 10, e0118744. [Google Scholar] [CrossRef] [PubMed]
- Harada, K.; Ozaki, S.; Sudo, Y.; Tsuneyama, K.; Ohta, H.; Nakanuma, Y. Osteopontin is involved in the formation of epithelioid granuloma and bile duct injury in primary biliary cirrhosis. Pathol. Int. 2003, 53, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Seth, D.; Duly, A.; Kuo, P.C.; McCaughan, G.W.; Haber, P.S. Osteopontin is an important mediator of alcoholic liver disease via hepatic stellate cell activation. World J. Gastroenterol. 2014, 20, 13088–13104. [Google Scholar] [CrossRef] [PubMed]
- Geisler, F.; Strazzabosco, M. Emerging roles of Notch signaling in liver disease. Hepatology 2015, 61, 382–392. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.J.; Clouston, A.D.; Forbes, S.J. Links between hepatic fibrosis, ductular reaction, and progenitor cell expansion. Gastroenterology 2014, 146, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lopategi, A.; Ge, X.; Lu, Y.; Kitamura, N.; Urtasun, R.; Leung, T.M.; Fiel, M.I.; Nieto, N. Osteopontin induces ductular reaction contributing to liver fibrosis. Gut 2014, 63, 1805–1818. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Burghardt, R.C.; Johnson, G.A.; White, F.J.; Ramaiah, S.K. The temporal expression of osteopontin (SPP-1) in the rodent model of alcoholic steatohepatitis: A potential biomarker. Toxicol. Pathol. 2006, 34, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Lazaro, R.; Wu, R.; Lee, S.; Zhu, N.L.; Chen, C.L.; French, S.W.; Xu, J.; Machida, K.; Tsukamoto, H. Osteopontin deficiency does not prevent but promotes alcoholic neutrophilic hepatitis in mice. Hepatology 2015, 61, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Leung, T.M.; Arriazu, E.; Lu, Y.; Urtasun, R.; Christensen, B.; Fiel, M.I.; Mochida, S.; Sørensen, E.S.; Nieto, N. Osteopontin binding to lipopolysaccharide lowers tumor necrosis factor-α and prevents early alcohol-induced liver injury in mice. Hepatology 2014, 59, 1600–1616. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Chen, S.; Zhao, C.; Li, X.; Zhao, W.; Yang, J.; Chang, C.; Xu, C. Comparative analysis of gene expression profiles of OPN signaling pathway in four kinds of liver diseases. J. Genet. 2016, 95, 741–750. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhao, C.; Chen, S.; Li, X.; Zhang, L.; Chang, C.; Xu, C. A preliminary in vivo study of the effects of OPN on rat liver regeneration induced by partial hepatectomy. Mol. Biol. Rep. 2016, 43, 1371–1382. [Google Scholar] [CrossRef] [PubMed]
- Nagoshi, S. Osteopontin: Versatile modulator of liver disease. Hepatol. Res. 2014, 44, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.W.; Ou, Y.H.; Peng, S.Y.; Liu, S.H.; Lai, P.L.; Lee, P.H.; Sheu, J.C.; Chen, C.L.; Hsu, H.C. Overexpression of osteopontin is associated with intrahepatic metastasis, early recurrence, and poorer prognosis of surgically resected hepatocellular carcinoma. Cancer 2003, 98, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.C.; Lee, Y.S.; Lin, S.E.; Wu, H.Y.; Chen, T.C.; Lee, W.C.; Chen, M.F.; Tsai, C.N. Recurrence and poor prognosis following resection of small hepatitis B-related hepatocellular carcinoma lesions are associated with aberrant tumor expression profiles of glypican 3 and osteopontin. Ann. Surg. Oncol. 2012, 19, S455–S463. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Dong, L.; Lu, B.; Wu, G.; Xu, D.; Chen, J.; Li, K.; Tong, X.; Dai, J.; Yao, S.; et al. Down-regulation of osteopontin suppresses growth and metastasis of hepatocellular carcinoma via induction of apoptosis. Gastroenterology 2008, 135, 956–968. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Park, J.W.; Woo, S.H.; Go, D.M.; Kwon, H.J.; Jang, J.J.; Kim, D.Y. Suppression of osteopontin inhibits chemically induced hepatic carcinogenesis by induction of apoptosis in mice. Oncotarget 2016, 7, 87219–87231. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.T.; Liu, T.S. Altered mucin gene expression in stone-containing intrahepatic bile ducts and cholangiocarcinomas. Dig. Dis. Sci. 2001, 46, 2166–2172. [Google Scholar] [CrossRef] [PubMed]
- Nakanuma, Y.; Yamaguchi, K.; Ohta, G.; Terada, T. Pathologic features of hepatolithiasis in Japan. Hum. Pathol. 1998, 19, 1181–1186. [Google Scholar] [CrossRef]
- Nakanuma, Y.; Sasaki, M.; Terada, T.; Harada, K. Intrahepatic peribiliary glands of humans. II. Pathological spectrum. J. Gastroenterol. Hepatol. 1994, 9, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K. Intrahepatic periductal glands and their significance in primary intrahepatic lithiasis. Jpn. J. Surg. 1982, 12, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.S.; Joo, S.H.; Lim, S.J.; Joo, K.R. Osteopontin Expression in Patients with Hepatolith. Indian J. Surg. 2015, 77, 551–556. [Google Scholar] [CrossRef] [PubMed]
- Nakai, A.; Imano, M.; Takeyama, Y.; Shiozaki, H.; Ohyanagi, H. An immunohistochemical study of osteopontin in hepatolithiasis. J. Hepatobiliary Pancreat. Surg. 2008, 15, 615–621. [Google Scholar] [CrossRef] [PubMed]
- Terashi, T.; Aishima, S.; Taguchi, K.; Asayama, Y.; Sugimachi, K.; Matsuura, S.; Shimada, M.; Maehara, S.; Maehara, Y.; Tsuneyoshi, M. Decreased expression of osteopontin is related to tumor aggressiveness and clinical outcome of intrahepatic cholangiocarcinoma. Liver Int. 2004, 24, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Sulpice, L.; Rayar, M.; Desille, M.; Turlin, B.; Fautrel, A.; Boucher, E.; Llamas-Gutierrez, F.; Meunier, B.; Boudjema, K.; Clément, B.; et al. Molecular profiling of stroma identifies osteopontin as an independent predictor of poor prognosis in intrahepatic cholangiocarcinoma. Hepatology 2013, 58, 1992–2000. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Jiao, J.; Cermelli, S.; Muir, K.; Jung, K.H.; Zou, R.; Rashid, A.; Gagea, M.; Zabludoff, S.; Kalluri, R.; et al. miR-21 Inhibition Reduces Liver Fibrosis and Prevents Tumor Development by Inducing Apoptosis of CD24+ Progenitor Cells. Cancer Res. 2015, 75, 1859–1867. [Google Scholar] [CrossRef] [PubMed]
- Razumilava, N.; Gores, G.J. Cholangiocarcinoma. Lancet 2014, 383, 2168–2179. [Google Scholar] [CrossRef]
- Rizvi, S.; Gores, G.J. Pathogenesis, diagnosis, and management of cholangiocarcinoma. Gastroenterology 2013, 145, 1215–1229. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Tanaka, H.; Tsukuma, H.; Ioka, A.; Oshima, A.; Nakahara, T. Risk factors for intrahepatic cholangiocarcinoma: A possible role of hepatitis B virus. J. Viral Hepat. 2010, 17, 742–748. [Google Scholar] [CrossRef] [PubMed]
- Fickert, P.; Thueringer, A.; Moustafa, T.; Silbert, D.; Gumhold, J.; Tsybrovskyy, O.; Lebofsky, M.; Jaeschke, H.; Denk, H.; Trauner, M. The role of osteopontin and tumor necrosis factor α receptor-1 in xenobiotic-induced cholangitis and biliary fibrosis in mice. Lab. Investig. 2010, 90, 844–852. [Google Scholar] [CrossRef] [PubMed]
- Pereira, T.A.; Syn, W.K.; Machado, M.V.; Vidigal, P.V.; Resende, V.; Voieta, I.; Xie, G.; Otoni, A.; Souza, M.M.; Santos, E.T.; et al. Schistosome-induced cholangiocyte proliferation and osteopontin secretion correlate with fibrosis and portal hypertension in human and murine schistosomiasis mansoni. Clin. Sci. 2015, 129, 875–883. [Google Scholar] [CrossRef] [PubMed]
- Tannapfel, A.; Sommerer, F.; Benicke, M.; Katalinic, A.; Uhlmann, D.; Witzigmann, H.; Hauss, J.; Wittekind, C. Mutations of the BRAF gene in cholangiocarcinoma but not in hepatocellular carcinoma. Gut 2003, 52, 706–712. [Google Scholar] [CrossRef] [PubMed]
- Zheng, T.; Hong, X.; Wang, J.; Pie, T.; Liang, Y.; Yin, D.; Song, R.; Song, X.; Lu, Z.; Qi, S.; et al. Gankyrin promotes tumor growth and metastasis through activation of IL-6/STAT3 signaling in human cholangiocarcinoma. Hepatology 2014, 59, 935–946. [Google Scholar] [CrossRef] [PubMed]
- Isomoto, H.; Mott, J.L.; Kobayashi, S.; Warneburg, N.W.; Bronk, S.F.; Haan, S.; Gores, G.J. Sustained IL-6/STAT-3 signaling in cholangiocarcinoma cells due to SOCS-3 epigenetic silencing. Gastroenterology 2007, 132, 384–396. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Han, C.; Wu, T. 15-hydroxyprostaglandin dehydrogenase-derived 15-keto-prostaglandin E2 inhibits cholangiocarcinoma cell growth through interaction with peroxisome proliferator-activated receptor-γ, SMAD2/3, and TAP63 proteins. J. Biol. Chem. 2013, 288, 19484–19502. [Google Scholar] [CrossRef] [PubMed]
- Techasen, A.; Namwat, N.; Loilome, W.; Bungkanjana, P.; Khuntikeo, N.; Puapairoj, A.; Jearanaikoon, P.; Saya, H.; Yongvanit, P. Tumor necrosis factor-α (TNF-α) stimulates the epithelial-mesenchymal transition regulator Snail in cholangiocarcinoma. Med. Oncol. 2012, 29, 3083–3091. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.X.; Cui, J.Y.; Ma, H.; Jia, X.M.; Huang, F.L.; Jiang, L.X. Screening of potential biomarkers for cholangiocarcinoma by integrated analysis of microarray data sets. Cancer Gene Ther. 2016, 23, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Laohaviroj, M.; Chamgramol, Y.; Pairojkul, C.; Mulvenna, J.; Sripa, B. Clinicopathological Significance of Osteopontin in Cholangiocarcinoma Cases. Asian Pac. J. Cancer Prev. 2016, 17, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Loosen, S.H.; Roderburg, C.; Kauertz, K.L.; Pombeiro, I.; Leyh, C.; Benz, F.; Vucur, M.; Longerich, T.; Koch, A.; Braunschweig, T.; et al. Elevated levels of circulating osteopontin are associated with a poor survival after resection of cholangiocarcinoma. J. Hepatol. 2017, 67, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Lustri, A.M.; Di Matteo, S.; Fraveto, A.; Costantini, D.; Cantafora, A.; Napoletano, C.; Bragazzi, M.C.; Giuliante, F.; De Rose, A.M.; Berloco, P.B.; et al. TGF-β signaling is an effective target to impair survival and induce apoptosis of human cholangiocarcinoma cells: A study on human primary cell cultures. PLoS ONE 2017, 12, e0183932. [Google Scholar] [CrossRef] [PubMed]
- Isomoto, H.; Kobayashi, S.; Werneburg, N.W.; Bronk, S.F.; Guicciardi, M.E.; Frank, D.A.; Gores, G.J. Interleukin 6 upregulates myeloid cell leukemia-1 expression through a STAT3 pathway in cholangiocarcinoma cells. Hepatology 2005, 42, 1329–1338. [Google Scholar] [CrossRef] [PubMed]
- Takemoto, M.; Yokote, K.; Yamazaki, M.; Ridall, A.L.; Butler, W.T.; Matsumoto, T.; Tamura, K.; Saito, Y.; Mori, S. Enhanced expression of osteopontin by high glucose in cultured rat aortic smooth muscle cells. Biochem. Biophys. Res. Commun. 1999, 258, 722–726. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, M.S.; Zhong, W.B.; Yu, S.C.; Lin, J.Y.; Chi, W.M.; Lee, H.M. Dipyridamole suppresses high glucose-induced osteopontin secretion and mRNA expression in rat aortic smooth muscle cells. Circ. J. 2010, 74, 1242–1250. [Google Scholar] [CrossRef] [PubMed]
- Katakam, A.K.; Chipitsyna, G.; Gong, Q.; Vancha, A.R.; Gabβ, J.; Arafat, H.A. Streptozotocin (STZ) mediates acute upregulation of serum and pancreatic osteopontin (OPN): A novel islet-protective effect of OPN through inhibition of STZ-induced nitric oxide production. J. Endocrinol. 2005, 187, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Arafat, H.A.; Katakam, A.K.; Chipitsyna, G.; Gong, Q.; Vancha, A.R.; Gabβ, J.; Dafoe, D.C. Osteopontin protects the islets and β-cells from interleukin-1 β-mediated cytotoxicity through negative feedback regulation of nitric oxide. Endocrinology 2007, 148, 575–584. [Google Scholar] [CrossRef] [PubMed]
- Lyssenko, V.; Eliasson, L.; Kotova, O.; Pilgaard, K.; Wierup, N.; Salehi, A.; Wendt, A.; Jonsson, A.; De Marinis, Y.Z.; Berglund, L.M.; et al. Pleiotropic effects of GIP on islet function involve osteopontin. Diabetes 2011, 60, 2424–2433. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Oka, M.; Iizuka, N.; Kawauchi, S.; Gondo, T.; Ueno, T.; Tangoku, A. Osteopontin expression in chronic pancreatitis. Pancreas 2002, 25, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Takada, H.; Nakazawa, T.; Ohara, H.; Ando, T.; Hayashi, K.; Naito, I.; Okumura, F.; Tanaka, H.; Yamada, T.; Takahashi, S.; et al. Role of osteopontin in calcification in autoimmune pancreatitis. Dig. Dis. Sci. 2009, 54, 793–801. [Google Scholar] [CrossRef] [PubMed]
- Rychlíková, J.; Vecka, M.; Jáchymová, M.; Macášek, J.; Hrabák, P.; Zeman, M.; Vávrová, L.; Řoupal, J.; Krechler, T.; Ák, A. Osteopontin as a discriminating marker for pancreatic cancer and chronic pancreatitis. Cancer Biomark. 2016, 17, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Kolb, A.; Kleeff, J.; Guweidhi, A.; Esposito, I.; Giese, N.A.; Adwan, H.; Giese, T.; Büchler, M.W.; Berger, M.R. Friess, H. Osteopontin influences the invasiveness of pancreatic cancer cells and is increased in neoplastic and inflammatory conditions. Cancer Biol. Ther. 2005, 4, 740–746. [Google Scholar] [CrossRef] [PubMed]
- Li, J.J.; Li, H.Y.; Gu, F. Diagnostic significance of serum osteopontin level for pancreatic cancer: A meta-analysis. Genet. Test. Mol. Biomark. 2014, 18, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Makino, N.; Matsuda, A.; Ikeda, Y.; Kakizaki, Y.; Saito, Y.; Ueno, Y.; Kawata, S. High Glucose Accelerates Cell Proliferation and Increases the Secretion and mRNA Expression of Osteopontin in Human Pancreatic Duct Epithelial Cells. Int. J. Mol. Sci. 2017, 18, 807. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.L.; Rock, J.; Malhotra, L.; Frankel, W.L.; Bloomston, M. Osteopontin expression is associated with improved survival in patients with pancreatic adenocarcinoma. Ann. Surg. Oncol. 2012, 19, 2673–2678. [Google Scholar] [CrossRef] [PubMed]
- Weber, C.E.; Erşahin, Ç.H.; Kuo, P.C.; Mi, Z. Pancreatic Cancer and Osteopontin: The Relationship Remains Unclear. Pancreas 2016, 45, e35–e36. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, J.L.; Levine, J.E.; Reddy, P.; Holler, E. Graft-versus-host disease. Lancet 2009, 373, 1550–1561. [Google Scholar] [CrossRef]
- Markey, K.A.; MacDonald, K.P.; Hill, G.R. The biology of graft-versus-host disease: Experimental systems instructing clinical practice. Blood 2014, 124, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Zhang, Y.; Wang, H.; Jin, M.; He, S.; Shi, Y.; Guo, Y.; Zhang, Y. Blockade of osteopontin reduces alloreactive CD8+T cell-mediated graft-versus-host disease. Blood 2011, 117, 1723–1733. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, K.; Minami, N.; Matsuura, M.; Iida, T.; Toyonaga, T.; Nagaishi, K.; Arimura, Y.; Fujimiya, M.; Uede, T.; Nakase, H. Osteopontin attenuates acute gastrointestinal graft-versus-host disease by preventing apoptosis of intestinal epithelial cells. Biochem. Biophys. Res. Commun. 2017, 485, 468475. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Organ | Disease | Osteopontin-Induced Apoptosis | Reference |
|---|---|---|---|
| Esophagus | Esophageal adenocarcinoma | ↓ | [52,53] |
| Stomach | Gastric caner | ↓ | [59,60,61,62] |
| Colon | Inflammatory bowel disease | ↓ | [63] |
| Colitis-associated cancer | ↓? | [63,67] | |
| Graft-versus-host disease | ↑/↓ | [129]/[130] | |
| Liver | Acute liver failure | ↑ | [83] |
| Nonalcoholic fatty liver disease | ↑ | [83] | |
| Hepatocellular carcinoma | ↓ | [88,89] | |
| Bile duct | Cholangiocarcinoma | ↓? | [113] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iida, T.; Wagatsuma, K.; Hirayama, D.; Nakase, H. Is Osteopontin a Friend or Foe of Cell Apoptosis in Inflammatory Gastrointestinal and Liver Diseases? Int. J. Mol. Sci. 2018, 19, 7. https://doi.org/10.3390/ijms19010007
Iida T, Wagatsuma K, Hirayama D, Nakase H. Is Osteopontin a Friend or Foe of Cell Apoptosis in Inflammatory Gastrointestinal and Liver Diseases? International Journal of Molecular Sciences. 2018; 19(1):7. https://doi.org/10.3390/ijms19010007
Chicago/Turabian StyleIida, Tomoya, Kohei Wagatsuma, Daisuke Hirayama, and Hiroshi Nakase. 2018. "Is Osteopontin a Friend or Foe of Cell Apoptosis in Inflammatory Gastrointestinal and Liver Diseases?" International Journal of Molecular Sciences 19, no. 1: 7. https://doi.org/10.3390/ijms19010007