Excessive Endoplasmic Reticulum Stress Correlates with Impaired Mitochondrial Dynamics, Mitophagy and Apoptosis, in Liver and Adipose Tissue, but Not in Muscles in EMS Horses

 ,
,
Abstract
:
1. Introduction
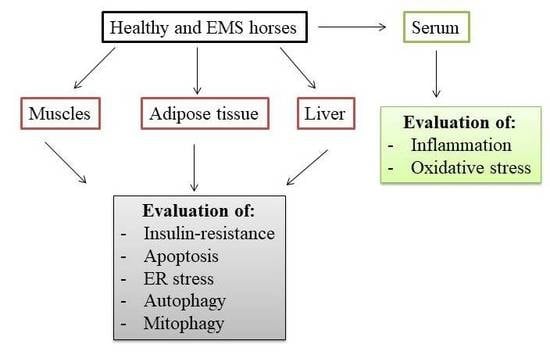
2. Results
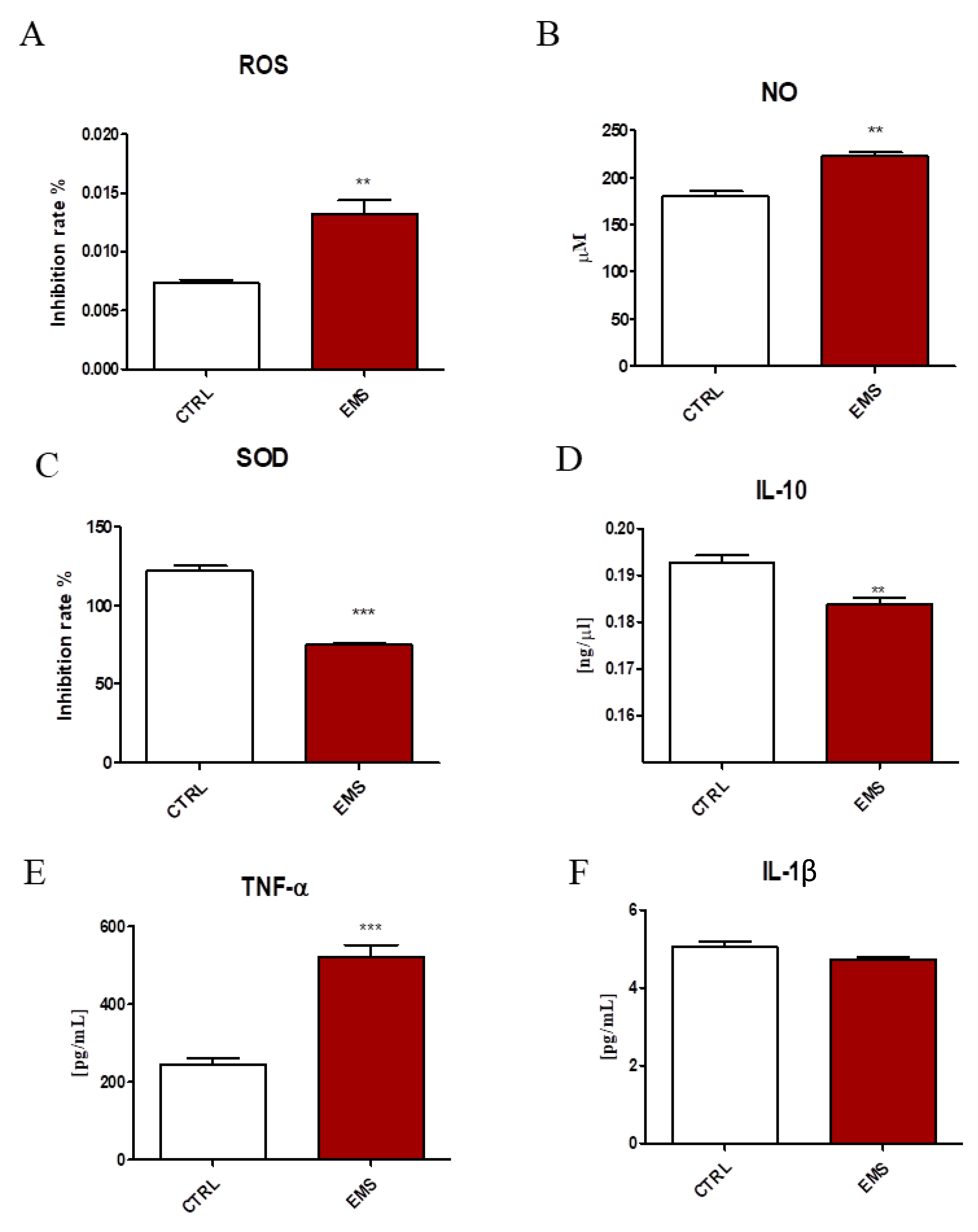
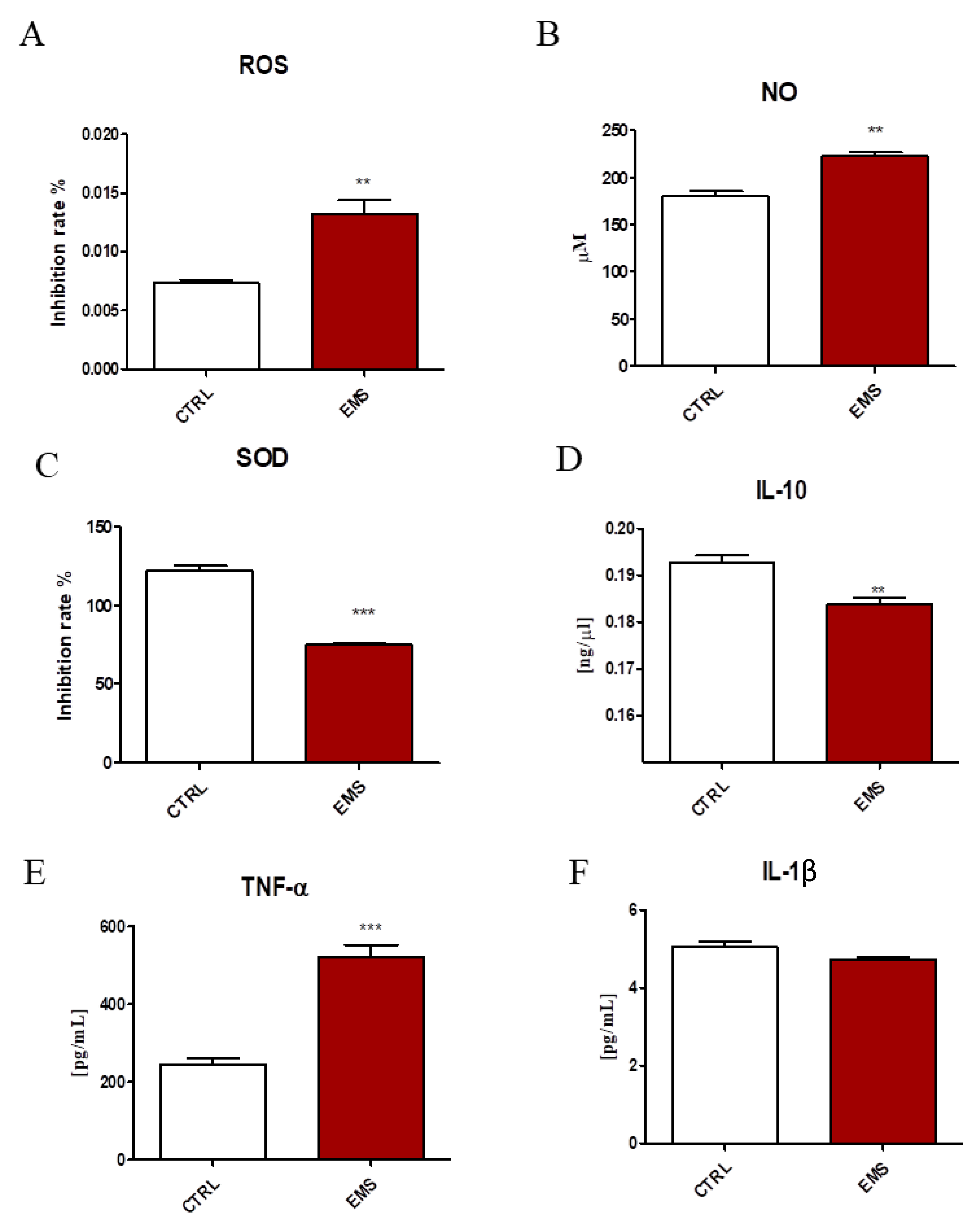
2.1. Oxidative Stress and Inflammation
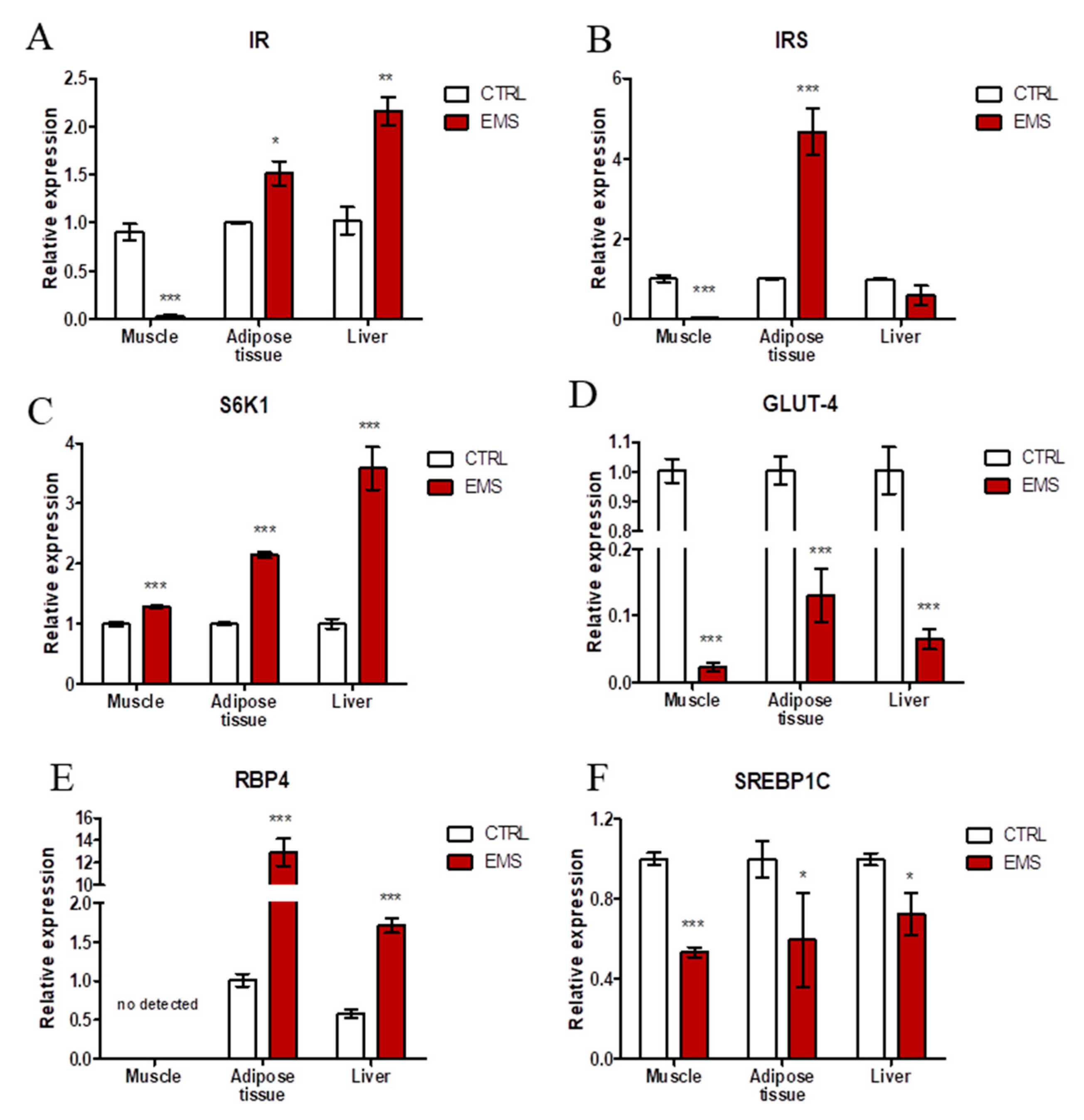
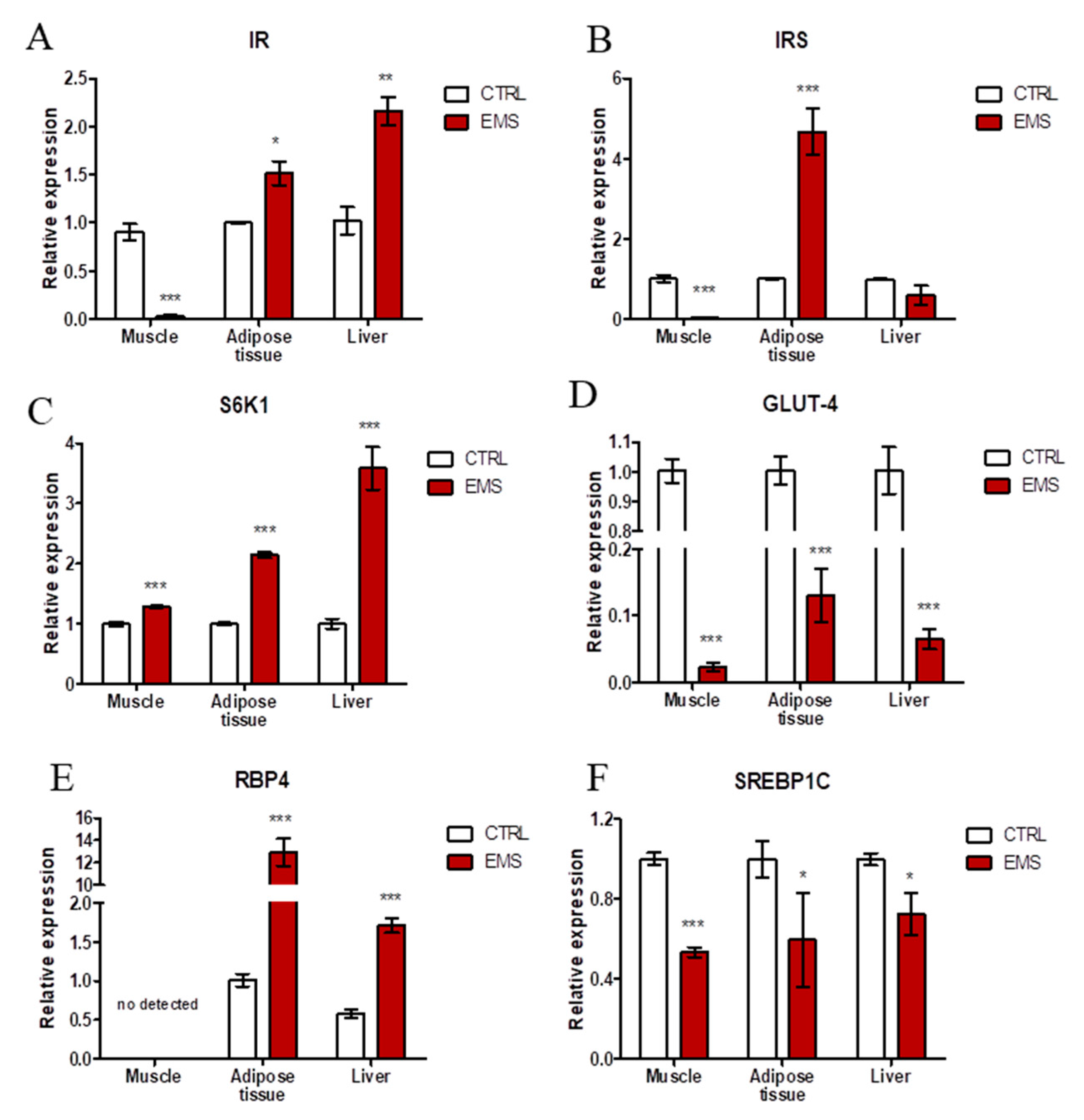
2.2. Insulin Resistance
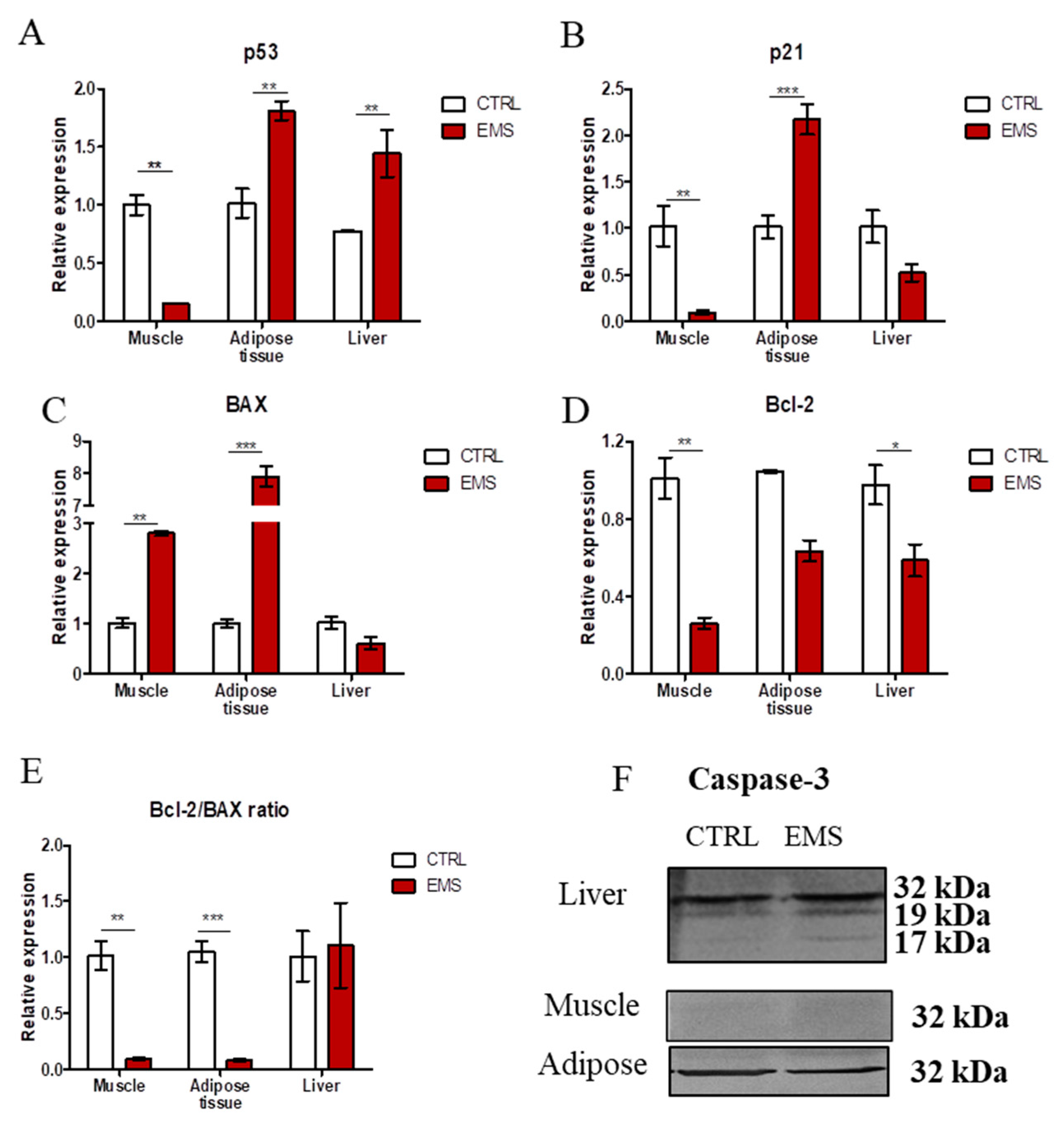
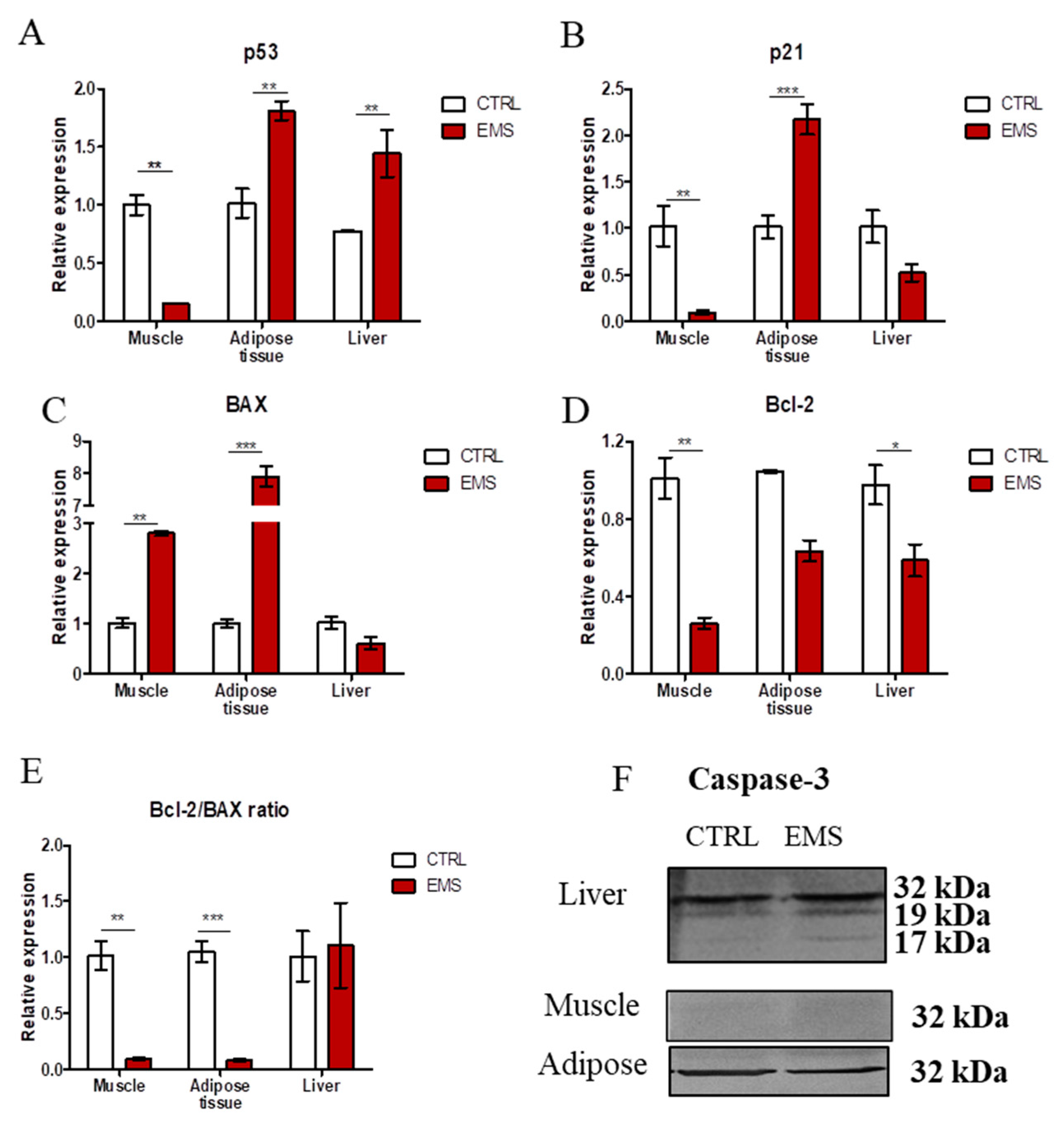
2.3. Apoptosis
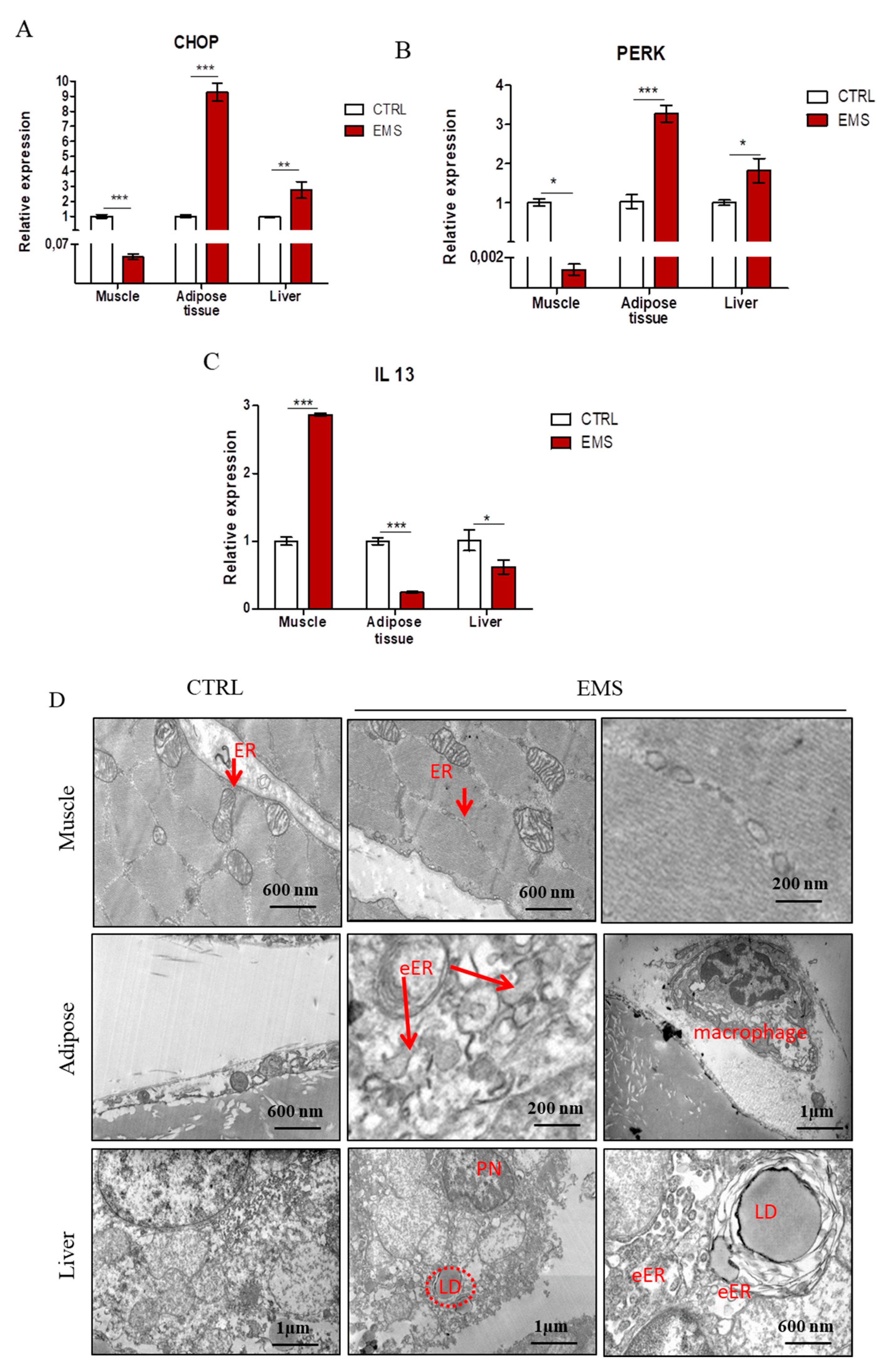
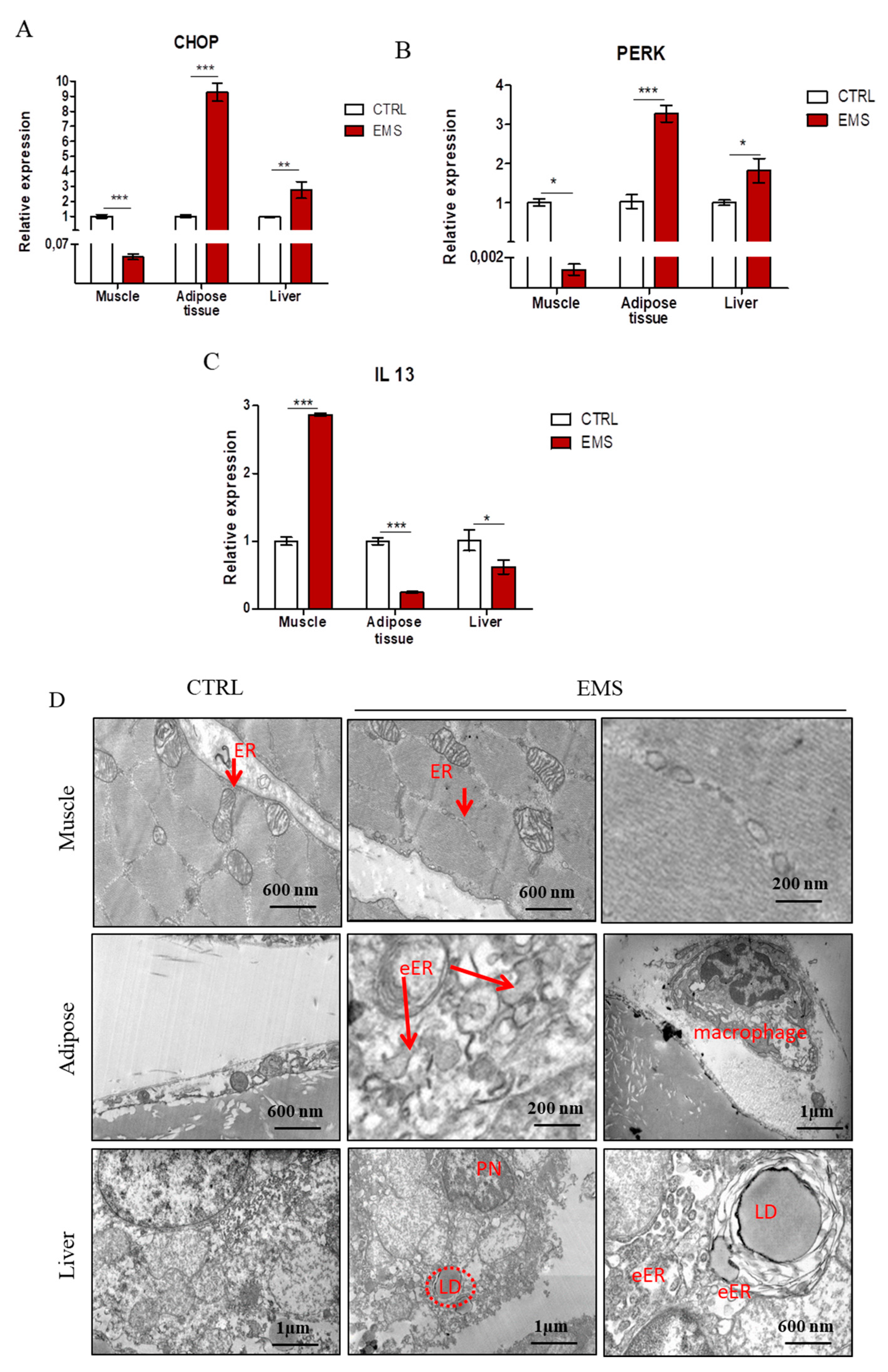
2.4. Endoplasmic Reticulum (ER) Stress
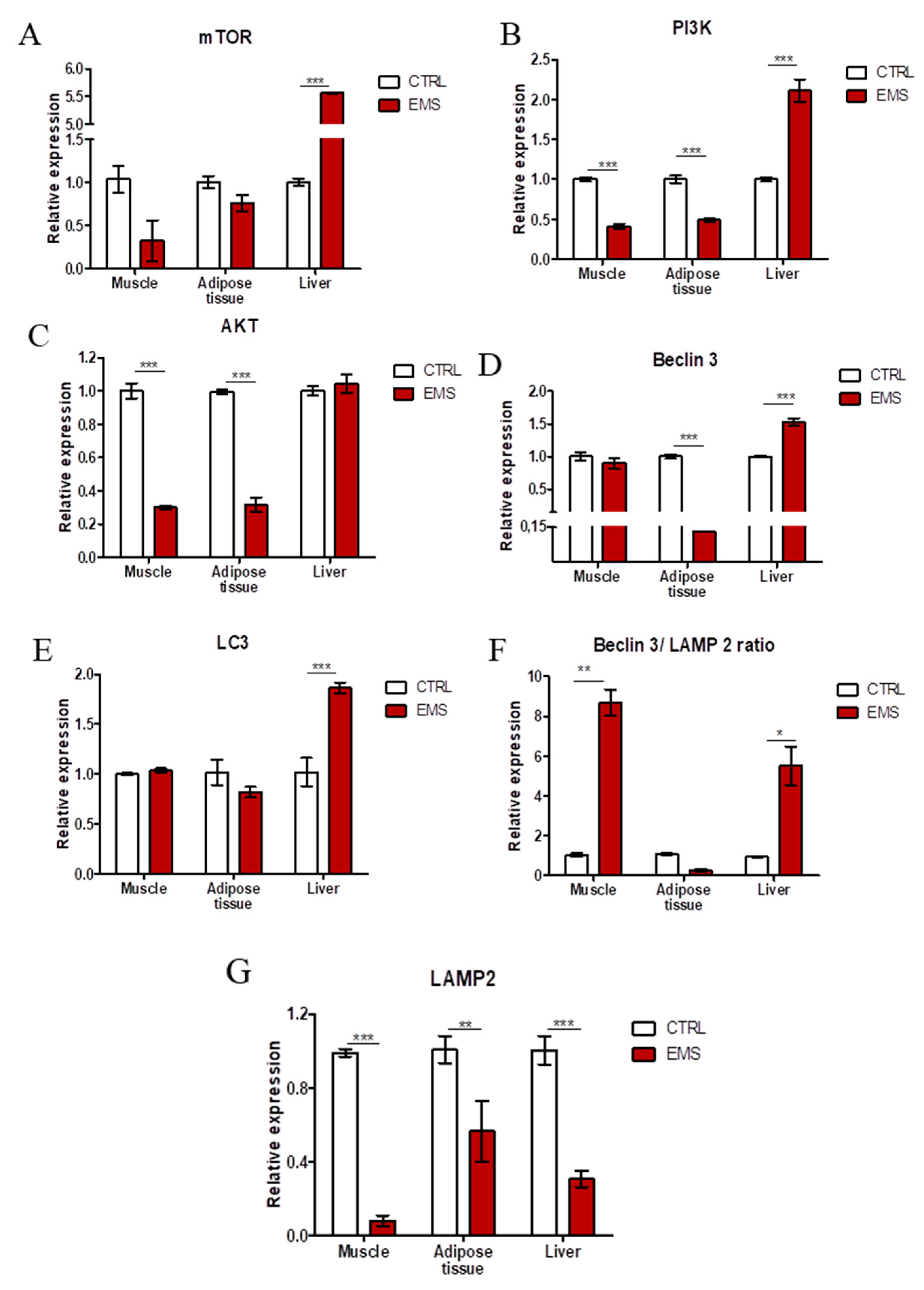
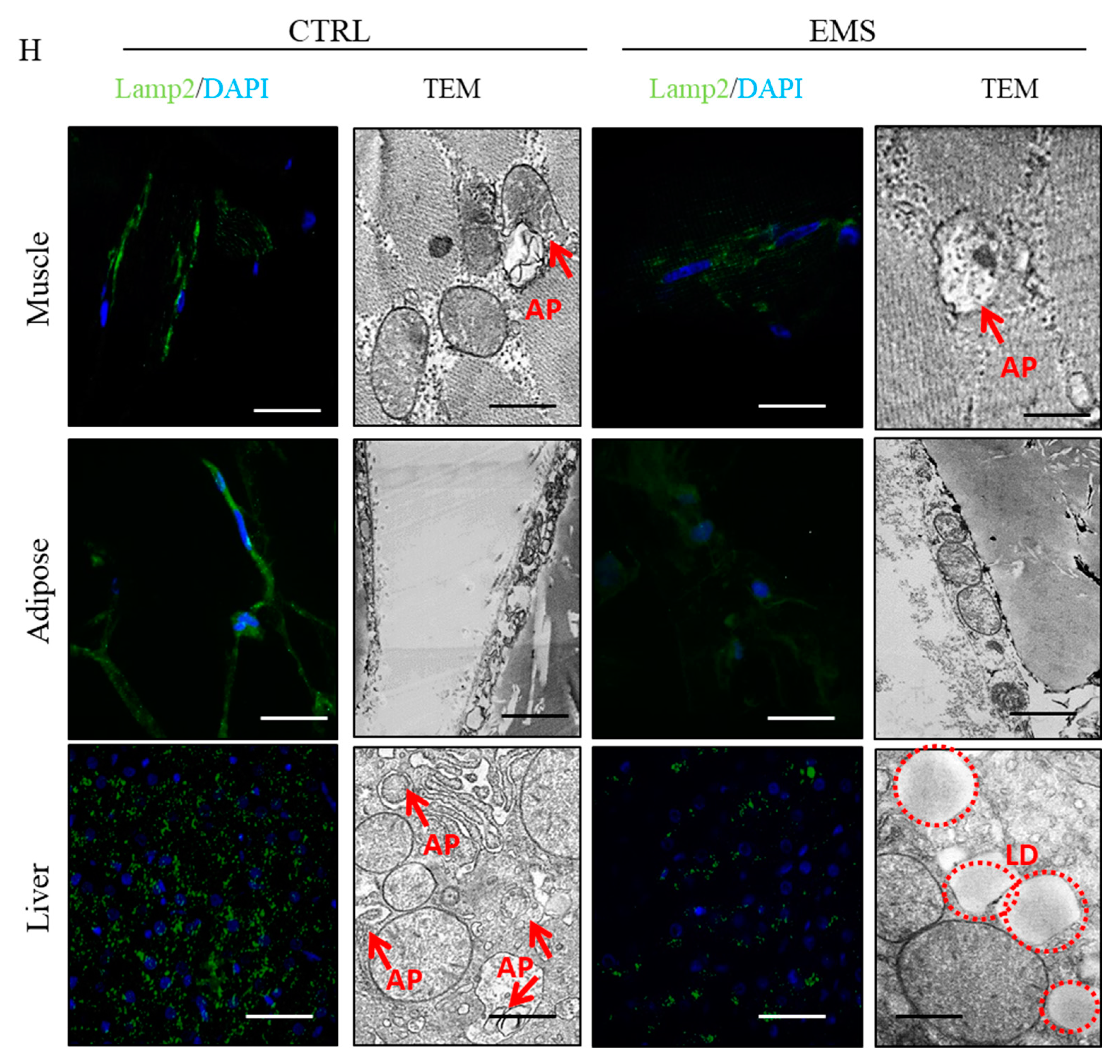
2.5. Autophagy
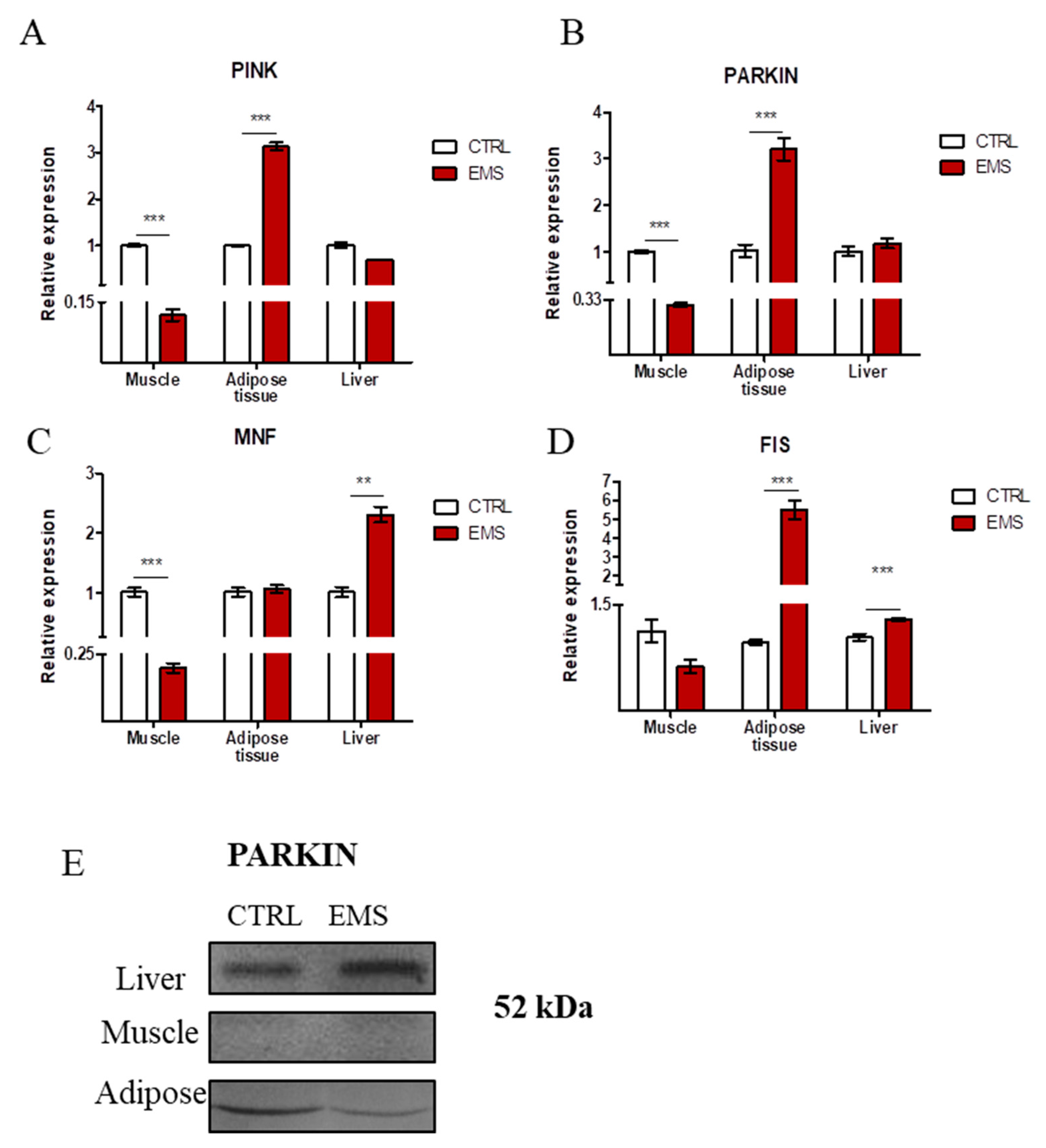
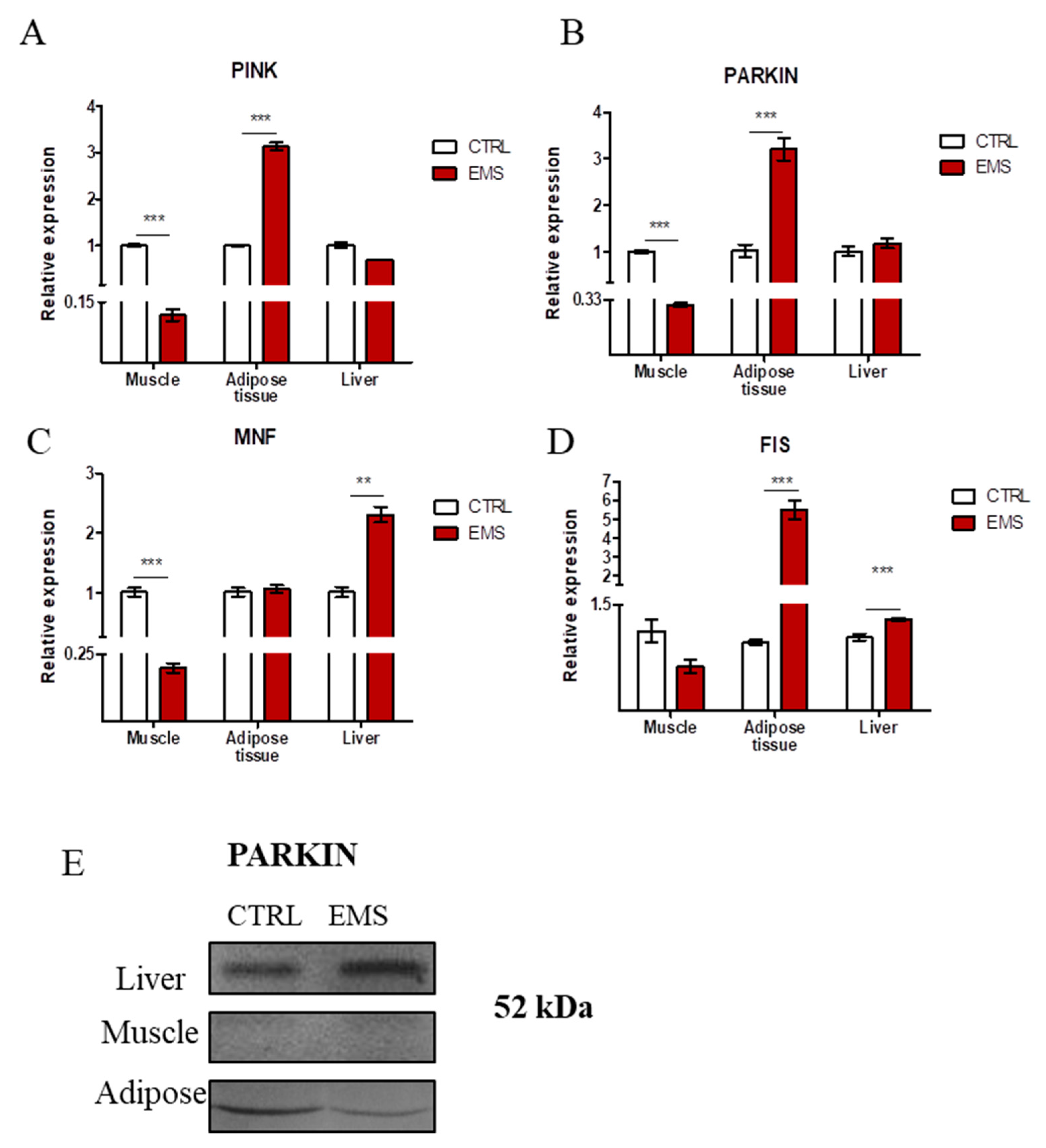
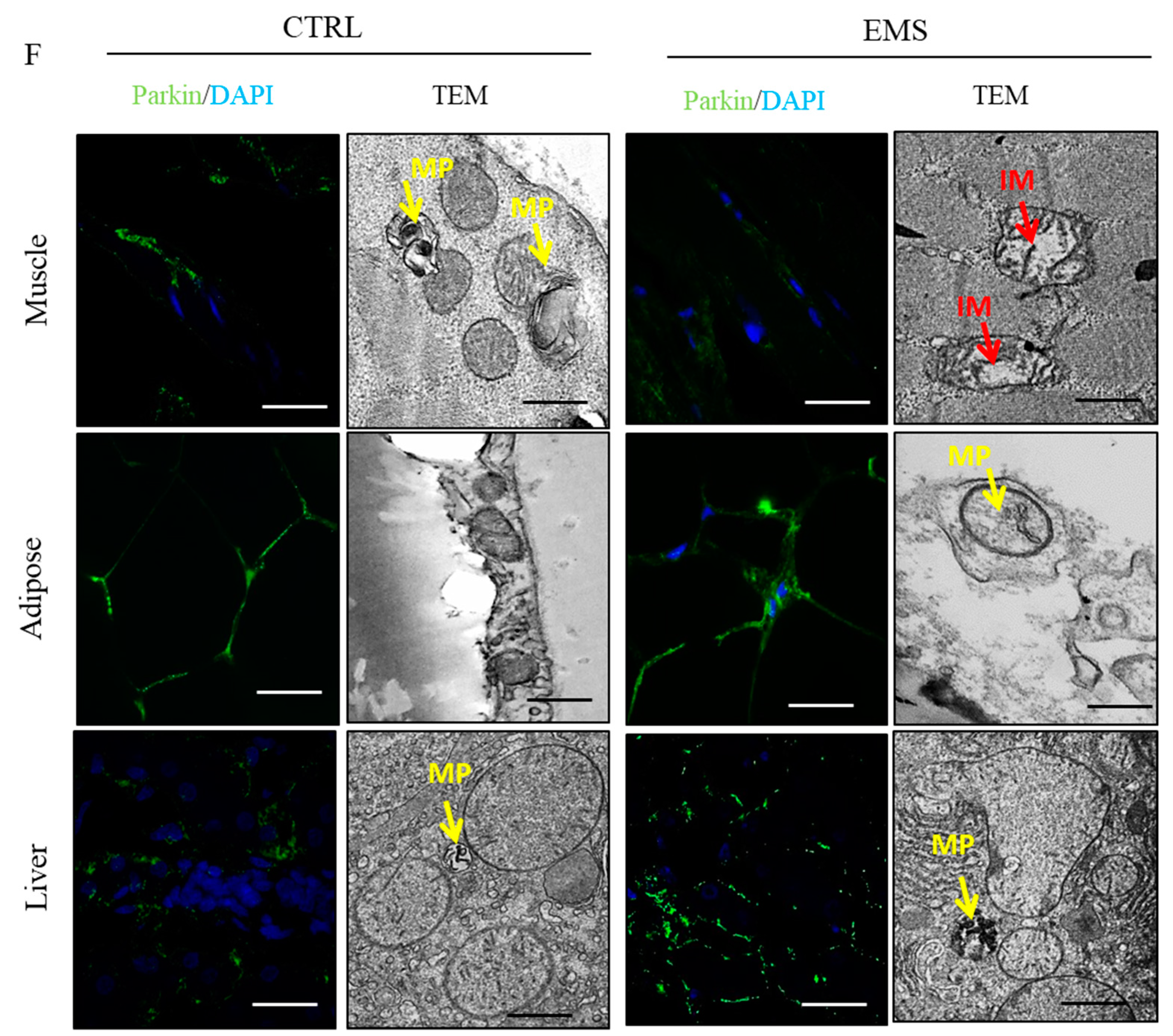
2.6. Mitophagy and Mitochondrial Dynamics
3. Discussion
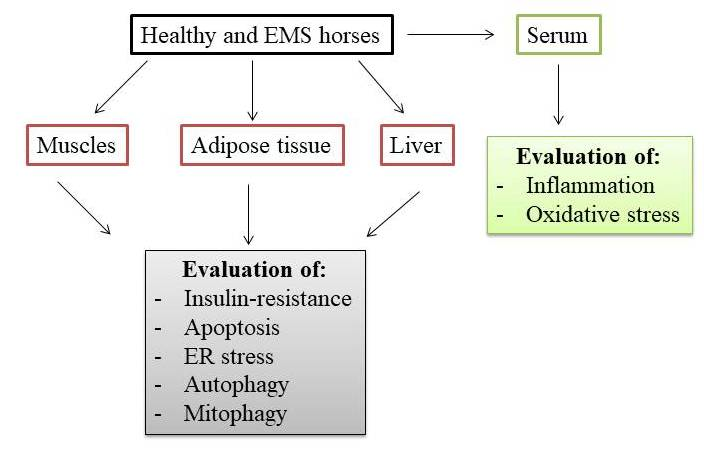
4. Materials and Methods
4.1. Horse Selection and EMS Diagnosis
4.2. Collection of Muscle, Adipose and Liver
4.3. Isolation of Proteins from Muscle, Adipose and Liver
4.4. Western Blotting
4.5. ELISA
4.6. Immunofluorescence and TEM
4.7. Oxidative Stress Factors in Serum Analysis
4.8. Analysis of Genes Expression
4.9. Statistics
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| BCS | Body condition score |
| CNS | Cresty neck score |
| EMS | Equine metabolic syndrome |
| Wt | Weight |
| AKT | RAC alpha serine/threonine-protein kinase |
| BAX | Bcl-2-associated X protein |
| Bcl-2 | B-cell lymphoma 2 |
| CHOP | CCAAT-enhancer-binding protein homologous protein |
| EMS | Equine metabolic syndrome |
| ER | Endoplasmic reticulum |
| FIS | Mitochondrial fission 1 protein |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| GLUT-4 | Glucose transporter 4 |
| IL-1β | Interleukin 1β |
| IL-10 | Interleukin 10 |
| IL-13 | Interleukin 13 |
| IL-4 | Interleukin 4 |
| IR | Insulin receptor |
| IRS | Insulin receptor substrate |
| LAMP2 | Lysosome-associated membrane protein 2 |
| LC-3 | Microtubule-associated protein 1A/1B-light chain 3 |
| MCP-1 | Monocyte chemotactic protein 1 |
| MNF | Mitofusin 1 |
| mTOR | The mechanistic target of rapamycin |
| NO | Nitric oxide |
| OS | Oxidative stress |
| p21 | Cyclin-dependent kinase inhibitor 1 |
| p53 | Tumor protein p53 |
| PARKIN | Parkin ligase |
| PERK | Eukaryotic translation initiation factor 2-alpha kinase 3 |
| PI3K | Phosphatidylinositol-4,5-bisphosphate 3-kinase |
| PINK1 | PTEN-induced putative kinase 1 |
| RBP4 | Retinol binding protein 4 |
| RICTOR | Rapamycin-insensitive companion of mammalian target of rapamycin |
| ROS | Reactive oxygen species |
| S6K1 | Ribosomal protein S6 kinase beta-1 |
| SOD | Superoxide dismutase |
| SREBP1C | Sterol regulatory element binding protein 1c |
| TNF-α | Tumor necrosis factor α |
References
- Geor, R.J.; McCue, M.E.; Schultz, N. Current understanding of the equine metabolic syndrome phenotype. J. Equine Vet. Sci. 2013, 33, 841–844. [Google Scholar] [CrossRef]
- Fantuzzi, G. Adipose tissue, adipokines, and inflammation. J. Allergy Clin. Immunol. 2005, 115, 911–919. [Google Scholar] [CrossRef] [PubMed]
- Singer, G.; Granger, D.N. Inflammatory responses underlying the microvascular dysfunction associated with obesity and insulin resistance. Microcirculation 2007, 14, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Tadros, E.M.; Frank, N.; Donnell, R.L. Effects of equine metabolic syndrome on inflammatory responses of horses to intravenous lipopolysaccharide infusion. Am. J. Vet. Res. 2013, 74, 1010–1019. [Google Scholar] [CrossRef] [PubMed]
- Basinska, K.; Marycz, K.; Śmieszek, A.; Nicpoń, J. The production and distribution of IL-6 and TNF-α in subcutaneous adipose tissue and their correlation with serum concentrations in Welsh ponies with equine metabolic syndrome. J. Vet. Sci. 2015, 16, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Burns, T.A.; Geor, R.J.; Mudge, M.C.; McCutcheon, L.J.; Hinchcliff, K.W.; Belknap, J.K. Proinflammatory cytokine and chemokine gene expression profiles in subcutaneous and visceral adipose tissue depots of insulin-resistant and insulin-sensitive light breed horses. J. Vet. Intern. Med. 2010, 24, 932–939. [Google Scholar] [CrossRef] [PubMed]
- Fujita, T. Insulin resistance and salt-sensitive hypertension in metabolic syndrome. Nephrol. Dial. Transplant. 2007, 22, 3102–3107. [Google Scholar] [CrossRef] [PubMed]
- Tangvarasittichai, S. Oxidative stress, insulin resistance, dyslipidemia and type 2 diabetes mellitus. World J. Diabetes 2015, 6, 456–480. [Google Scholar] [CrossRef] [PubMed]
- Soussi, H.; Clément, K.; Dugail, I. Adipose tissue autophagy status in obesity: Expression and flux—Two faces of the picture. Autophagy 2016, 12, 588–589. [Google Scholar] [CrossRef] [PubMed]
- Ozcan, U.; Cao, Q.; Yilmaz, E.; Lee, A.-H.; Iwakoshi, N.N.; Ozdelen, E.; Tuncman, G.; Görgün, C.; Glimcher, L.H.; Hotamisligil, G.S. Endoplasmic reticulum stress links obesity, insulin action, and type 2 diabetes. Science 2004, 306, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Cnop, M.; Foufelle, F.; Velloso, L.A. Endoplasmic reticulum stress, obesity and diabetes. Trends Mol. Med. 2012, 18, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Sarparanta, J.; García-Macia, M.; Singh, R. Autophagy and Mitochondria in Obesity and Type 2 Diabetes. Curr. Diabetes Rev. 2017, 13, 352–369. [Google Scholar] [CrossRef] [PubMed]
- McCue, M.E.; Geor, R.J.; Schultz, N. Equine Metabolic Syndrome: A Complex Disease Influenced by Genetics and the Environment. J. Equine Vet. Sci. 2015, 35, 367–375. [Google Scholar] [CrossRef]
- VanderJagt, D.J.; Harrison, J.M.; Ratliff, D.M.; Hunsaker, L.A.; Vander Jagt, D.L. Oxidative stress indices in IDDM subjects with and without long-term diabetic complications. Clin. Biochem. 2001, 34, 265–270. [Google Scholar] [CrossRef]
- Martín-Gallán, P.; Carrascosa, A.; Gussinyé, M.; Domínguez, C. Biomarkers of diabetes-associated oxidative stress and antioxidant status in young diabetic patients with or without subclinical complications. Free Radic. Biol. Med. 2003, 34, 1563–1574. [Google Scholar] [CrossRef]
- Ogihara, T.; Asano, T.; Ando, K.; Chiba, Y.; Sakoda, H.; Anai, M.; Shojima, N.; Ono, H.; Onishi, Y.; Fujishiro, M.; et al. Angiotensin II-induced insulin resistance is associated with enhanced insulin signaling. Hypertens. Dallas Tex 1979 2002, 40, 872–879. [Google Scholar] [CrossRef]
- Bardell, A.L.; MacLeod, K.M. Evidence for inducible nitric-oxide synthase expression and activity in vascular smooth muscle of streptozotocin-diabetic rats. J. Pharmacol. Exp. Ther. 2001, 296, 252–259. [Google Scholar] [PubMed]
- Tiedge, M.; Lortz, S.; Drinkgern, J.; Lenzen, S. Relation between antioxidant enzyme gene expression and antioxidative defense status of insulin-producing cells. Diabetes 1997, 46, 1733–1742. [Google Scholar] [CrossRef] [PubMed]
- Isogawa, A.; Yamakado, M.; Yano, M.; Shiba, T. Serum superoxide dismutase activity correlates with the components of metabolic syndrome or carotid artery intima-media thickness. Diabetes Res. Clin. Pract. 2009, 86, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Talior, I.; Yarkoni, M.; Bashan, N.; Eldar-Finkelman, H. Increased glucose uptake promotes oxidative stress and PKC-delta activation in adipocytes of obese, insulin-resistant mice. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E295–E302. [Google Scholar] [CrossRef] [PubMed]
- Rudich, A.; Tirosh, A.; Potashnik, R.; Hemi, R.; Kanety, H.; Bashan, N. Prolonged oxidative stress impairs insulin-induced GLUT4 translocation in 3T3-L1 adipocytes. Diabetes 1998, 47, 1562–1569. [Google Scholar] [CrossRef] [PubMed]
- Nayak, D.U.; Karmen, C.; Frishman, W.H.; Vakili, B.A. Antioxidant vitamins and enzymatic and synthetic oxygen-derived free radical scavengers in the prevention and treatment of cardiovascular disease. Heart Dis. Hagerstown Md 2001, 3, 28–45. [Google Scholar] [CrossRef]
- Ford, E.S. Vitamin Supplement Use and Diabetes Mellitus Incidence among Adults in the United States. Am. J. Epidemiol. 2001, 153, 892–897. [Google Scholar] [CrossRef] [PubMed]
- Codogno, P.; Meijer, A.J. Autophagy: A potential link between obesity and insulin resistance. Cell Metab. 2010, 11, 449–451. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, M.K.; Turner, N. Mitochondrial dysfunction and insulin resistance: An update. Endocr. Connect. 2014, 4, R1–R15. [Google Scholar] [CrossRef] [PubMed]
- Catalán, V.; Gómez-Ambrosi, J.; Rodríguez, A.; Ramírez, B.; Andrada, P.; Rotellar, F.; Valentí, V.; Moncada, R.; Martí, P.; Silva, C.; et al. Expression of S6K1 in human visceral adipose tissue is upregulated in obesity and related to insulin resistance and inflammation. Acta Diabetol. 2015, 52, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Ribel-Madsen, R.; Friedrichsen, M.; Vaag, A.; Poulsen, P. Retinol-Binding Protein 4 in Twins. Diabetes 2009, 58, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-H.; Hsieh, T.-J.; Lin, K.-D.; Lin, H.-Y.; Lee, M.-Y.; Hung, W.-W.; Hsiao, P.-J.; Shin, S.-J. Increased Unbound Retinol-binding Protein 4 Concentration Induces Apoptosis through Receptor-mediated Signaling. J. Biol. Chem. 2012, 287, 9694–9707. [Google Scholar] [CrossRef] [PubMed]
- Minamino, T.; Orimo, M.; Shimizu, I.; Kunieda, T.; Yokoyama, M.; Ito, T.; Nojima, A.; Nabetani, A.; Oike, Y.; Matsubara, H.; et al. A crucial role for adipose tissue p53 in the regulation of insulin resistance. Nat. Med. 2009, 15, 1082–1087. [Google Scholar] [CrossRef] [PubMed]
- Back, S.H.; Kaufman, R.J. Endoplasmic reticulum stress and type 2 diabetes. Annu. Rev. Biochem. 2012, 81, 767–793. [Google Scholar] [CrossRef] [PubMed]
- Machado, M.V.; Yang, Y.; Diehl, A.M. The benefits of restraint: A pivotal role for IL-13 in hepatic glucose homeostasis. J. Clin. Invest. 2013, 123, 115–117. [Google Scholar] [CrossRef] [PubMed]
- Guicciardi, M.E.; Gores, G.J. Apoptosis: A mechanism of acute and chronic liver injury. Gut 2005, 54, 1024–1033. [Google Scholar] [CrossRef] [PubMed]
- Sims-Robinson, C.; Bakeman, A.; Rosko, A.; Glasser, R.; Feldman, E.L. The Role of Oxidized Cholesterol in Diabetes-Induced Lysosomal Dysfunction in the Brain. Mol. Neurobiol. 2016, 53, 2287–2296. [Google Scholar] [CrossRef] [PubMed]
- Scott, I.; Youle, R.J. Mitochondrial fission and fusion. Essays Biochem. 2010, 47, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Barazzoni, R. Modulating Mitochondrial Fission to Lower Diabetic Oxidative Stress. Diabetes 2012, 61, 1915–1917. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhang, M.; Torres, G.; Wu, S.; Ouyang, C.; Xie, Z.; Zou, M.-H. Metformin Suppresses Diabetes-Accelerated Atherosclerosis via the Inhibition of Drp1-Mediated Mitochondrial Fission. Diabetes 2017, 66, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Schuver, A.; Frank, N.; Chameroy, K.A.; Elliott, S.B. Assessment of Insulin and Glucose Dynamics by Using an Oral Sugar Test in Horses. J. Equine Vet. Sci. 2014, 34, 465–470. [Google Scholar] [CrossRef]
- Marędziak, M.; Marycz, K.; Lewandowski, D.; Siudzińska, A.; Śmieszek, A. Static magnetic field enhances synthesis and secretion of membrane-derived microvesicles (MVs) rich in VEGF and BMP-2 in equine adipose-derived stromal cells (EqASCs)-a new approach in veterinary regenerative medicine. In Vitro Cell. Dev. Biol. Anim. 2015, 51, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Kornicka, K.; Marycz, K.; Tomaszewski, K.A.; Marędziak, M.; Śmieszek, A. The Effect of Age on Osteogenic and Adipogenic Differentiation Potential of Human Adipose Derived Stromal Stem Cells (hASCs) and the Impact of Stress Factors in the Course of the Differentiation Process. Oxid. Med. Cell. Longev. 2015, 2015, 309169. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Age | Baseline Serum Insulin; (mIU/mL) | Insulin (mIU/mL) 60 min Post Oral Sugar Administration | Baseline Glucose (mg/dL) | Glucose 60 min Post Oral Sugar Administration (mg/dL) | BCS | CNS | LEP (ng/mL) | Wt (Kg) |
|---|---|---|---|---|---|---|---|---|---|
| EMS | 12 ± 2 | 180 ± 20 | 260 ± 11 | 72 ± 11 | 163 ± 26 | 7.2 ± 0.2 | 3.9 ± 0.7 | 4.89 ± 1.2 | 680 ± 30 |
| Control | 11 ± 2 | 13 ± 4 | 36 ± 3 | 84 | 88 | 6.4±0.3 | 1.9±0.6 | 1.1±0.6 | 590 ± 20 |
| No. | Gene Name | Primer Sequences | Amplicon Size | Accession Number | |
|---|---|---|---|---|---|
| 1 | AKT | Forward 5′ → 3′ | AAGGAGATCATGCAGCACCG | 180 | XM_014854427.1 |
| Reverse 5′ → 3′ | CTCCATCGTGTCGTCTTGGT | ||||
| 2 | BAX | Forward 5′ → 3′ | GCCAGCAAATTGGTGCTCAA | 260 | XM_014830923.1 |
| Reverse 5′ → 3′ | AGCAGTCACTTCCATGGCTC | ||||
| 3 | Bcl-2 | Forward 5′ → 3′ | TTCTTTGAGTTCGGTGGGGT | 164 | XM_014843802.1 |
| Reverse 5′ → 3′ | GGGCCGTACAGTTCCACAA | ||||
| 4 | Beclin-3 | Forward 5′ → 3′ | GATGCGTTATGCCCAGATGC | 233 | XM_014833759.1 |
| Reverse 5′ → 3′ | AACGGCAGCTCCTCTGAAAT | ||||
| 5 | CHOP | Forward 5′ → 3′ | AGCCAAAATCAGAGCCGGAA | 272 | XM_014844003.1 |
| Reverse 5′ → 3′ | GGGGTCAAGAGTGGTGAAGG | ||||
| 6 | FIS | Forward 5′ → 3′ | GGTGCGAAGCAAGTACAACG | 118 | XM_014854003.1 |
| Reverse 5′ → 3′ | GTTGCCCACAGCCAGATAGA | ||||
| 7 | GLUT-4 | Forward 5′ → 3′ | AAGCCCTCGCTACCTCTACA | 210 | NM_001081866.2 |
| Reverse 5′ → 3′ | TGCAGCACAACTGCAATGAC | ||||
| 8 | IL-1β | Forward 5′ → 3′ | TATGTGTGTGATGCAGCTGTGC | 352 | XM_014852743.1 |
| Reverse 5′ → 3′ | GGCCACAGGTATCTTGTCAGT | ||||
| 9 | IL-4 | Forward 5′ → 3′ | TGACTGTAGCGGATGCCTTT | 129 | XM_014856772.1 |
| Reverse 5′ → 3′ | GTCCGCTCAGGCATTCTTTG | ||||
| 10 | IL-10 | Forward 5′ → 3′ | TGTTGTTGAACGGGTCCCTG | 242 | NM_001082490.1 |
| Reverse 5′ → 3′ | ACTCTTCACCTGCTCCACTG | ||||
| 11 | IL-13 | Forward 5′ → 3′ | AGCTGGTCAACATCACCCAG | 150 | XM_014730431.1 |
| Reverse 5′ → 3′ | GCATCTTCCGCGTGTTTTGG | ||||
| 12 | IR | Forward 5′ → 3′ | CCGTTTGAGTCTGAGGGGTC | 254 | XM_014862015.1 |
| Reverse 5′ → 3′ | ACCGTCACATTCCCGACATC | ||||
| 13 | IRS | Forward 5′ → 3′ | CTGCTGGGGGTTTGGAGAAT | 254 | XM_014862015.1 |
| Reverse 5′ → 3′ | TAAATCCTCACTGGAGCGGC | ||||
| 14 | LAMP2 | Forward 5′ → 3′ | GCACCCCTGGGAAGTTCTTA | 147 | XM_014831347.1 |
| Reverse 5′ → 3′ | ATCCAGCGAACACTCTTGGG | ||||
| 15 | LC3 | Forward 5′ → 3′ | TTACTGCTTTGCTCTGCCAC | 213 | XM_014835085.1 |
| Reverse 5′ → 3′ | AGCTGCTTCTCCCCCTTGTA | ||||
| 16 | mTOR | Forward 5′ → 3′ | GGGCAGCATTAGAGACGGTG | 221 | XM_005607537.2 |
| Reverse 5′ → 3′ | ATGGTTGATTCGGTGTCGCA | ||||
| 17 | MNF | Forward 5′ → 3′ | AAGTGGCATTTTTCGGCAGG | 217 | XM_014838357.1 |
| Reverse 5′ → 3′ | TCCATATGAAGGGCATGGGC | ||||
| 18 | PARKIN | Forward 5′ → 3′ | TCCCAGTGGAGGTCGATTCT | 218 | XM_014858374.1 |
| Reverse 5′ → 3′ | CCCTCCAGGTGTGTTCGTTT | ||||
| 19 | PERK | Forward 5′ → 3′ | GTGACTGCAATGGACCAGGA | 283 | XM_014852775.1 |
| Reverse 5′ → 3′ | TCACGTGCTCACGAGGATATT | ||||
| 20 | PINK1 | Forward 5′ → 3′ | GCACAATGAGCCAGGAGCTA | 298 | XM_014737247.1 |
| Reverse 5′ → 3′ | GGGGTATTCACGCGAAGGTA | ||||
| 21 | PI3K | Forward 5′ → 3′ | GACTTGCACTTGGGTGACATA | 152 | XM_014855332.1 |
| Reverse 5′ → 3′ | TAAGTTCCCGGAAAGTCCCC | ||||
| 22 | P21 | Forward 5′ → 3′ | GAAGAGAAACCCCCAGCTCC | 241 | XM_014853747.1 |
| Reverse 5′ → 3′ | TGACTGCATCAAACCCCACA | ||||
| 23 | P53 | Forward 5′ → 3′ | TACTCCCCTGCCCTCAACAA | 252 | U37120.1 |
| Reverse 5′ → 3′ | AGGAATCAGGGCCTTGAGGA | ||||
| 24 | RICTOR | Forward 5′ → 3′ | CTCCACATCGCGAGTCTGTC | 166 | XM_014738100.1 |
| Reverse 5′ → 3′ | ATCCAATTCAGCTCGCCCAA | ||||
| 25 | RBP4 | Forward 5′ → 3′ | AAGGGTCCAATCTGGCACG | 172 | NM_001081951.1 |
| Reverse 5′ → 3′ | CAAGTCCTGGCCTAGTCAGC | ||||
| 26 | SREBP1C | Forward 5′ → 3′ | TCAGCGAGGCGGCTTTGGACAG | 80 | XM_008542859.1 |
| Reverse 5′ → 3′ | CATGTCTTCGATGTCGGTCAG | ||||
| 27 | S6K1 | Forward 5′ → 3′ | GAGACAGGGAAGCTGAGGACAT | 245 | XM_014856960.1 |
| Reverse 5′ → 3′ | ACCAAGTACCCGAAGTAGCTC | ||||
| 28 | GAPDH | Forward 5′ → 3′ | GATGCCCCAATGTTTGTGA | 250 | XM_014866500.1 |
| Reverse 5′ → 3′ | AAGCAGGGATGATGTTCTGG | ||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marycz, K.; Kornicka, K.; Szlapka-Kosarzewska, J.; Weiss, C. Excessive Endoplasmic Reticulum Stress Correlates with Impaired Mitochondrial Dynamics, Mitophagy and Apoptosis, in Liver and Adipose Tissue, but Not in Muscles in EMS Horses. Int. J. Mol. Sci. 2018, 19, 165. https://doi.org/10.3390/ijms19010165
Marycz K, Kornicka K, Szlapka-Kosarzewska J, Weiss C. Excessive Endoplasmic Reticulum Stress Correlates with Impaired Mitochondrial Dynamics, Mitophagy and Apoptosis, in Liver and Adipose Tissue, but Not in Muscles in EMS Horses. International Journal of Molecular Sciences. 2018; 19(1):165. https://doi.org/10.3390/ijms19010165
Chicago/Turabian StyleMarycz, Krzysztof, Katarzyna Kornicka, Jolanta Szlapka-Kosarzewska, and Christine Weiss. 2018. "Excessive Endoplasmic Reticulum Stress Correlates with Impaired Mitochondrial Dynamics, Mitophagy and Apoptosis, in Liver and Adipose Tissue, but Not in Muscles in EMS Horses" International Journal of Molecular Sciences 19, no. 1: 165. https://doi.org/10.3390/ijms19010165