
EDIM-TKTL1/Apo10 Blood Test: An Innate Immune System Based Liquid Biopsy for the Early Detection, Characterization and Targeted Treatment of Cancer

Abstract
:1. Introduction
2. Biomarker Detection with EDIM Technology
3. DNaseX: A Biomarker for Abnormal Apoptosis in Tumor Cells
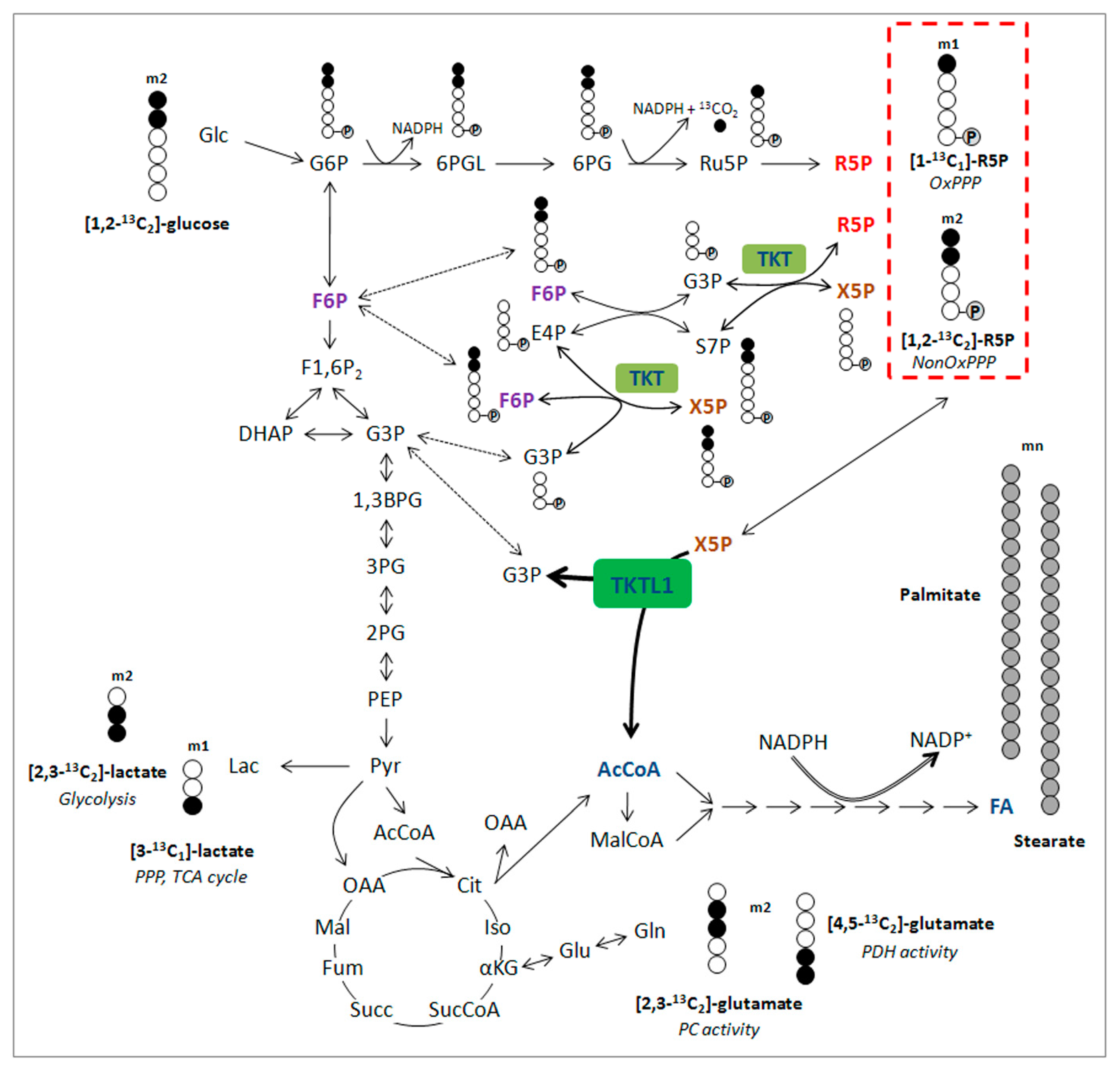
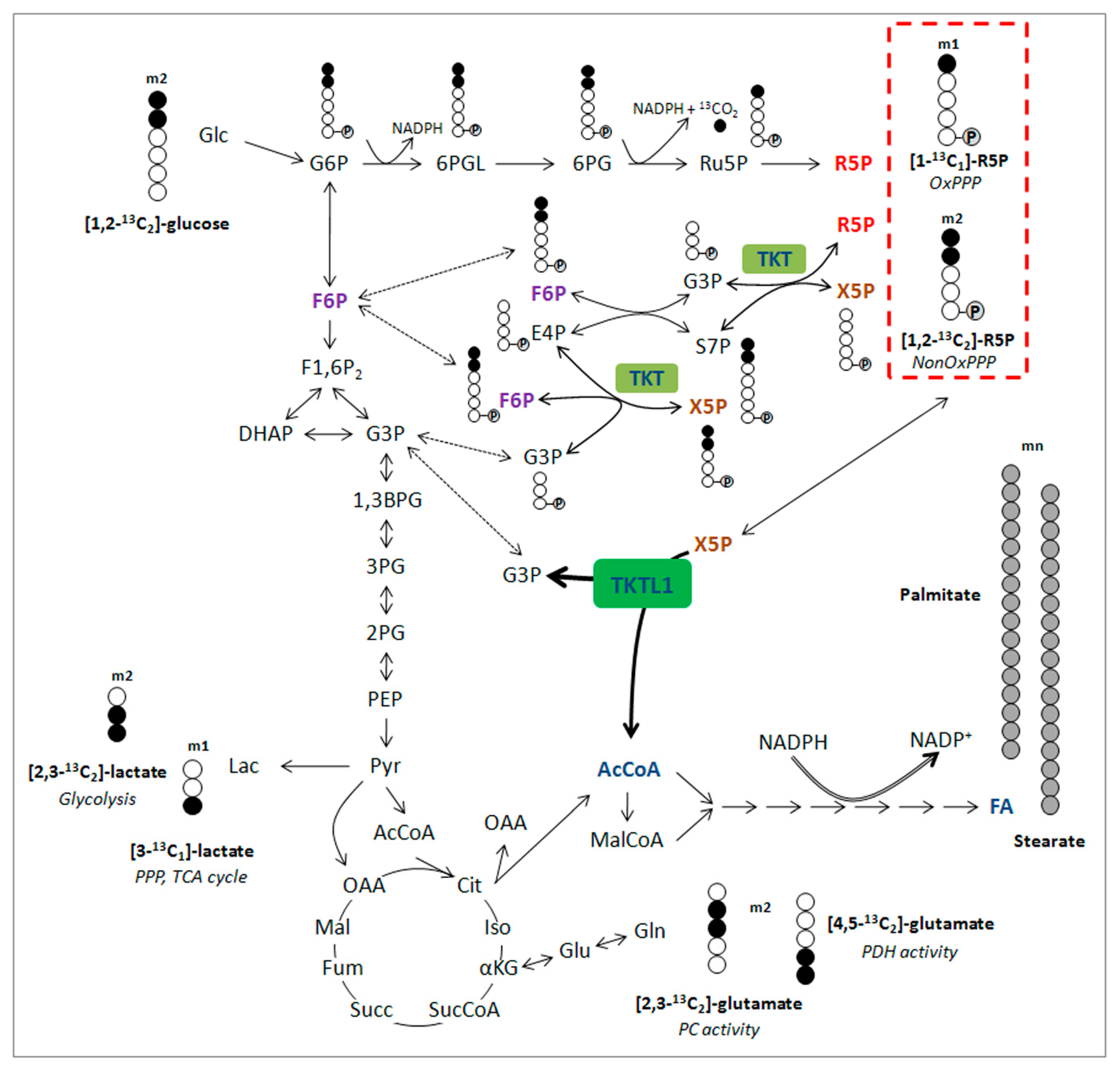
4. Diagnostic Significance of the TKTL1 Gene
5. The Second Way of Lactic Acid Fermentation in Mammals: A Great Moment during Evolution of Vertebrates
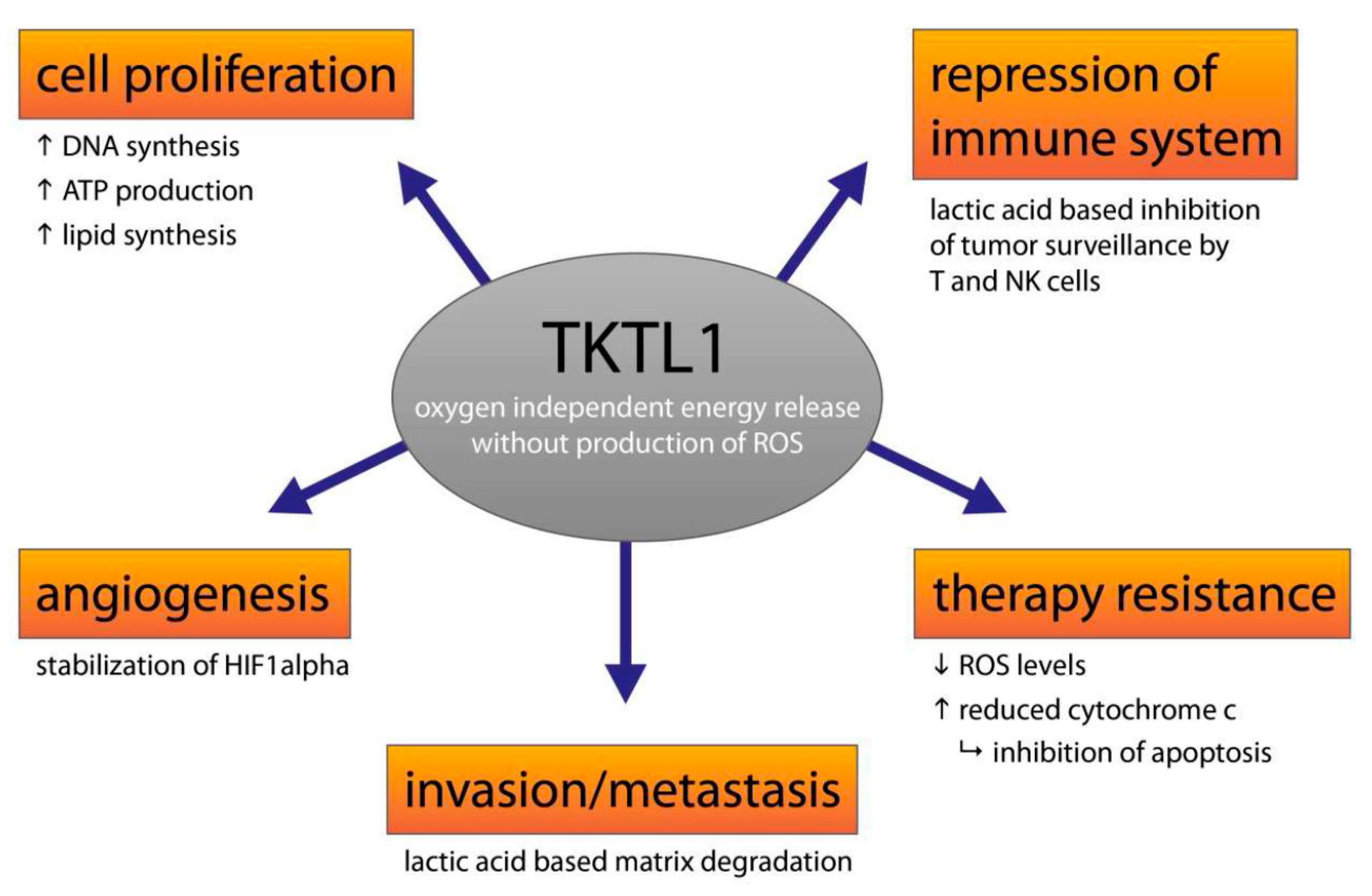
6. The Clinical Significance of Activating the TKTL1 Gene
7. Benfo-Oxythiamine: The Vitamin Antagonist Oxythiamine in a New Dress
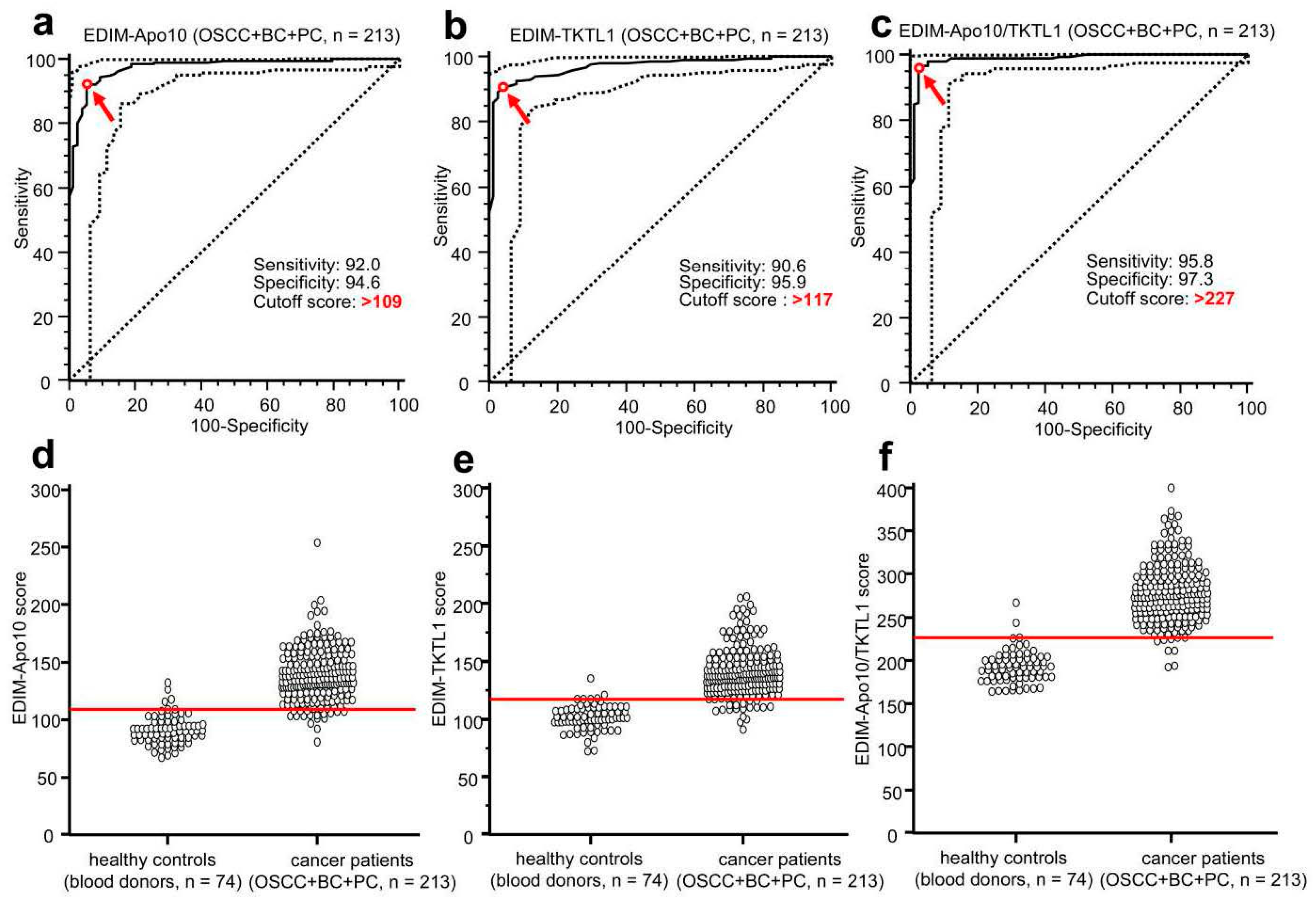
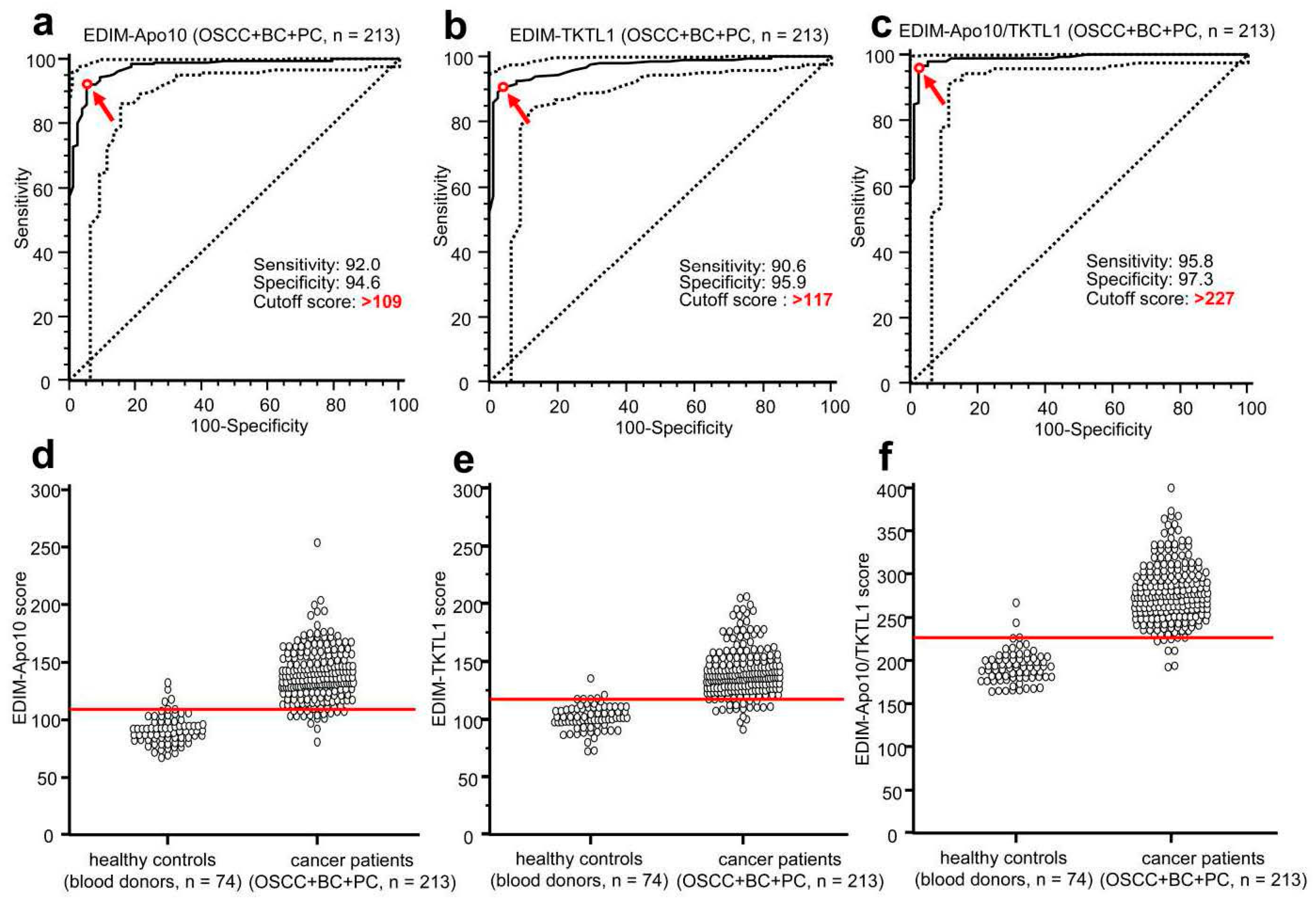
8. EDIM-TKTL1/Apo10 Blood Test: An Innate Immune System Based Liquid Biopsy for the Detection of Cancer Patients Eligible for a Targeted Treatment
9. EDIM-TKTL1/Apo10 Blood Test for the Detection of Patients Eligible for 18FDG-PET Imaging of Tumors
10. EDIM-TKTL1/Apo10 Blood Test for the Monitoring of Cancer Therapies
11. EDIM-TKTL1/Apo10 Blood Test for the Monitoring of Surgical Removal of Tumors
12. EDIM-TKTL1/Apo10 Blood Test for the Early Detection of Cancer Recurrence
13. EDIM-TKTL1/Apo10 Blood Test for the Early Detection of Cancer
14. Future Directions and Limitations
Acknowledgments
Conflicts of Interest
References
- Feyen, O.; Coy, J.F.; Prasad, V.; Schierl, R.; Saenger, J.; Baum, R.P. EDIM-TKTL1 blood test: A noninvasive method to detect upregulated glucose metabolism in patients with malignancies. Future Oncol. 2012, 8, 1349–1359. [Google Scholar] [CrossRef] [PubMed]
- Grimm, M.; Schmitt, S.; Teriete, P.; Biegner, T.; Stenzl, A.; Hennenlotter, J.; Muhs, H.J.; Munz, A.; Nadtotschi, T.; König, K.; et al. A biomarker based detection and characterization of carcinomas exploiting two fundamental biophysical mechanisms in mammalian cells. BMC Cancer 2013, 13, 569. [Google Scholar] [CrossRef] [PubMed]
- Grimm, M.; Kraut, W.; Hoefert, S.; Krimmel, M.; Biegner, T.; Teriete, P.; Cetindis, M.; Polligkeit, J.; Kluba, S.; Munz, A.; et al. Evaluation of a biomarker based blood test for monitoring surgical resection of oral squamous cell carcinomas. Clin. Oral Investig. 2016, 20, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Jansen, N.; Coy, J.F. Diagnostic use of epitope detection in monocytes blood test for early detection of colon cancer metastasis. Future Oncol. 2013, 9, 605–609. [Google Scholar] [CrossRef] [PubMed]
- Penault-Llorca, F.; Radosevic-Robin, N. Ki67 assessment in breast cancer: An update. Pathology 2017, 49, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Huszno, J.; Nowara, E. Current therapeutic strategies of anti-HER2 treatment in advanced breast cancer patients. Contemp. Oncol. 2016, 20, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ueland, F.R. A Perspective on Ovarian Cancer Biomarkers: Past, Present and Yet-To-Come. Diagnostics 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Recker, F.; Seiler, D.; Seifert, B.; Randazzo, M.; Kwiatkowski, M. PSA screening 2013: Background and perspectives. Urol. A 2014, 53, 875–881. [Google Scholar] [CrossRef] [PubMed]
- Herwig, R.; Pelzer, A.; Horninger, W.; Rehder, P.; Klocker, H.; Ramoner, R.; Pinggera, G.M.; Gozzi, C.; Konwalinka, G.; Bartsch, G. Measurement of intracellular versus extracellular prostate-specific antigen levels in peripheral macrophages: A new approach to noninvasive diagnosis of prostate cancer. Clin. Prostate Cancer 2004, 3, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Grimm, M.; Feyen, O.; Coy, J.F.; Hofmann, H.; Teriete, P.; Reinert, S. Analysis of circulating CD14+/CD16+ monocyte-derived macrophages (MDMs) in the peripheral blood of patients with oral squamous cell carcinoma. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2016, 121, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Leers, M.P.; Nap, M.; Herwig, R.; Delaere, K.; Nauwelaers, F. Circulating PSA-containing macrophages as a possible target for the detection of prostate cancer: A three-color/five-parameter flow cytometric study on peripheral blood samples. Am. J. Clin. Pathol. 2008, 129, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Japink, D.; Leers, M.P.; Sosef, M.N.; Nap, M. CEA in activated macrophages. New diagnostic possibilities for tumor markers in early colorectal cancer. Anticancer Res. 2009, 29, 3245–3251. [Google Scholar] [PubMed]
- Faber, T.J.; Japink, D.; Leers, M.P.; Sosef, M.N.; von Meyenfeldt, M.F.; Nap, M. Activated macrophages containing tumor marker in colon carcinoma: Immunohistochemical proof of a concept. Tumor Biol. 2012, 33, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Grimm, M.; Hoefert, S.; Krimmel, M.; Biegner, T.; Feyen, O.; Teriete, P.; Reinert, S. Monitoring carcinogenesis in a case of oral squamous cell carcinoma using a panel of new metabolic blood biomarkers as liquid biopsies. Oral Maxillofac. Surg. 2016, 20, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Coy, J.F.; Velhagen, I.; Himmele, R.; Delius, H.; Poustka, A.; Zentgraf, H. Isolation, differential splicing and protein expression of a DNase on the human X chromosome. Cell Death Differ. 1996, 3, 199–206. [Google Scholar] [PubMed]
- Taper, H.S. Altered deoxyribonuclease activity in cancer cells and its role in non-toxic adjuvant cancer therapy with mixed vitamins C and K3. Anticancer Res. 2008, 28, 2727–2732. [Google Scholar] [PubMed]
- Zanotti, S.; Fisseler-Eckhoff, A.; Mannherz, H.G. Changes in the topological expression of markers of differentiation and apoptosis in defined stages of human cervical dysplasia and carcinoma. Gynecol. Oncol. 2003, 89, 376–384. [Google Scholar] [CrossRef]
- Coy, J.F.; Dübel, S.; Kioschis, P.; Thomas, K.; Micklem, G.; Delius, H.; Poustka, A. Molecular cloning of tissue-specific transcripts of a transketolase-related gene: Implications for the evolution of new vertebrate genes. Genomics 1996, 32, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Coy, J.F.; Dressler, D.; Wilde, J.; Schubert, P. Mutations in the transketolase-like gene TKTL1: Clinical implications for neurodegenerative diseases, diabetes and cancer. Clin. Lab. 2005, 51, 257–273. [Google Scholar] [PubMed]
- Langbein, S.; Zerilli, M.; Zur Hausen, A.; Staiger, W.; Rensch-Boschert, K.; Lukan, N.; Popa, J.; Ternullo, M.P.; Steidler, A.; Weiss, C.; et al. Expression of transketolase TKTL1 predicts colon and urothelial cancer patient survival: Warburg effect reinterpreted. Br. J. Cancer 2006, 94, 578–585. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zur Hausen, A.; Coy, J.F.; Löchelt, M. Transketolase-like protein 1 (TKTL1) is required for rapid cell growth and full viability of human tumor cells. Int. J. Cancer 2009, 124, 1330–1337. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Liu, Y.; Glazer, C.A.; Shao, C.; Bhan, S.; Demokan, S.; Zhao, M.; Rudek, M.A.; Ha, P.K.; Califano, J.A. TKTL1 is activated by promoter hypomethylation and contributes to head and neck squamous cell carcinoma carcinogenesis through increased aerobic glycolysis and HIF1α stabilization. Clin. Cancer Res. 2010, 16, 857–866. [Google Scholar] [CrossRef] [PubMed]
- Jayachandran, A.; Lo, P.H.; Chueh, A.C.; Prithviraj, P.; Molania, R.; Davalos-Salas, M.; Anaka, M.; Walkiewicz, M.; Cebon, J.; Behren, A. Transketolase-like 1 ectopic expression is associated with DNA hypomethylation and induces the Warburg effect in melanoma cells. BMC Cancer 2016, 16, 134. [Google Scholar] [CrossRef] [PubMed]
- Warburg, O.; Posener, K.; Negelein, E. Über den Stoffwechsel der Carcinomzelle. Biochem. Z. 1924, 152, 309–344. [Google Scholar] [CrossRef]
- Diaz-Moralli, S.; Aguilar, E.; Marin, S.; Coy, J.F.; Dewerchin, M.; Antoniewicz, M.R.; Meca-Cortés, O.; Notebaert, L.; Ghesquière, B.; Eelen, G.; et al. A key role for Transketolase-like 1 in tumor metabolic reprogramming. Oncotarget 2016, 7, 51875–51897. [Google Scholar] [CrossRef] [PubMed]
- Hartmannsberger, D.; Mack, B.; Eggert, C.; Denzel, S.; Stepp, H.; Betz, C.S.; Gires, O. Transketolase-like protein 1 confers resistance to serum withdrawal in vitro. Cancer Lett. 2011, 300, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Rolland, A.D.; Lavigne, R.; Dauly, C.; Calvel, P.; Kervarrec, C.; Freour, T.; Evrard, B.; Rioux-Leclercq, N.; Auger, J.; Pineau, C. Identification of genital tract markers in the human seminal plasma using an integrative genomics approach. Hum. Reprod. 2013, 28, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Iglesias-Pedraz, J.M.; Chen, L.Y.; Yin, F.; Cadenas, E.; Reddy, S.; Comai, L. Downregulation of the Werner syndrome protein induces a metabolic shift that compromises redox homeostasis and limits proliferation of cancer cells. Aging Cell 2014, 13, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Demetrius, L.A.; Coy, J.F.; Tuszynski, J.A. Cancer proliferation and therapy: The Warburg effect and quantum metabolism. Theor. Biol. Med. Model. 2010, 7, 2. [Google Scholar] [CrossRef] [PubMed]
- Szent-Györgyi, A. The living state and cancer. Physiol. Chem. Phys. 1980, 12, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Horecker, L.; Gibbs, M.; Klenow, H.; Smyrniotis, P.Z. The mechanism of pentose phosphate conversion to hexose monophosphate. I. With a liver enzyme preparation. J. Biol. Chem. 1954, 207, 393–403. [Google Scholar] [PubMed]
- Vaughn, A.E.; Deshmukh, M. Glucose metabolism inhibits apoptosis in neurons and cancer cells by redox inactivation of cytochrome c. Nat. Cell Biol. 2008, 10, 1477–1483. [Google Scholar] [CrossRef] [PubMed]
- Newington, J.T.; Pitts, A.; Chien, A.; Arseneault, R.; Schubert, D.; Cumming, R.C. Amyloid beta resistance in nerve cell lines is mediated by the Warburg effect. PLoS ONE 2011, 6, e19191. [Google Scholar] [CrossRef] [PubMed]
- Prüfer, K.; Racimo, F.; Patterson, N.; Jay, F.; Sankararaman, S.; Sawyer, S.; Heinze, A.; Renaud, G.; Sudmant, P.H.; de Filippo, C.; et al. The complete genome sequence of a Neanderthal from the Altai Mountains. Nature 2014, 505, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Schwaab, J.; Horisberger, K.; Ströbel, P.; Bohn, B.; Gencer, D.; Kähler, G.; Kienle, P.; Post, S.; Wenz, F.; Hofmann, W.K.; et al. Expression of Transketolase like gene 1 (TKTL1) predicts disease-free survival in patients with locally advanced rectal cancer receiving neoadjuvant chemoradiotherapy. BMC Cancer 2011, 11, 363. [Google Scholar] [CrossRef] [PubMed]
- Kohrenhagen, N.; Voelker, H.U.; Schmidt, M.; Kapp, M.; Krockenberger, M.; Frambach, T.; Dietl, J.; Kammerer, U. Expression of transketolase-like 1 (TKTL1) and p-Akt correlates with the progression of cervical neoplasia. J. Obstet. Gynaecol. Res. 2008, 34, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Yue, J.X.; Yang, S.H.; Ding, H.; Zhao, R.W.; Zhang, S. Overexpression of transketolase-like gene 1 is associated with cell proliferation in uterine cervix cancer. J. Exp. Clin. Cancer Res. 2009, 28, 43. [Google Scholar] [CrossRef] [PubMed]
- Zerilli, M.; Amato, M.C.; Martorana, A.; Cabibi, D.; Coy, J.F.; Cappello, F.; Pompei, G.; Russo, A.; Giordano, C.; Rodolico, V. Increased expression of transketolase-like-1 in papillary thyroid carcinomas smaller than 1.5 cm in diameter is associated with lymph-node metastases. Cancer 2008, 113, 936–944. [Google Scholar] [CrossRef] [PubMed]
- Kayser, G.; Sienel, W.; Kubitz, B.; Mattern, D.; Stickeler, E.; Passlick, B.; Werner, M.; Zur Hausen, A. Poor outcome in primary non-small cell lung cancers is predicted by transketolase TKTL1 expression. Pathology 2011, 43, 719–724. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Liu, D.; He, G. TKTL1 and p63 are biomarkers for the poor prognosis of gastric cancer patients. Cancer Biomark. 2015, 15, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhu, S.C.; Li, S.G.; Zhao, Y.; Xu, J.R.; Song, C.Y. TKTL1 promotes cell proliferation and metastasis in esophageal squamous cell carcinoma. Biomed. Pharmacother. 2015, 74, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Ahopelto, K.; Böckelman, C.; Hagström, J.; Koskensalo, S.; Haglund, C. Transketolase-like protein 1 expression predicts poor prognosis in colorectal cancer. Cancer Biol. Ther. 2016, 17, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Fischer, K.; Hoffmann, P.; Voelkl, S.; Meidenbauer, N.; Ammer, J.; Edinger, M.; Gottfried, E.; Schwarz, S.; Rothe, G.; Hoves, S.; et al. Inhibitory effect of tumor cell-derived lactic acid on human T cells. Blood 2007, 109, 3812–3819. [Google Scholar] [CrossRef] [PubMed]
- Brand, A.; Singer, K.; Koehl, G.E.; Kolitzus, M.; Schoenhammer, G.; Thiel, A.; Matos, C.; Bruss, C.; Klobuch, S.; Peter, K.; et al. LDHA-Associated Lactic Acid Production Blunts Tumor Immunosurveillance by T and NK Cells. Cell Metab. 2016, 24, 657–671. [Google Scholar] [CrossRef] [PubMed]
- Xu, I.M.; Lai, R.K.; Lin, S.H.; Tse, A.P.; Chiu, D.K.; Koh, H.Y.; Law, C.T.; Wong, C.M.; Cai, Z.; Wong, C.C.; et al. Transketolase counteracts oxidative stress to drive cancer development. Proc. Natl. Acad. Sci. USA 2016, 113, E725–E734. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Mancuso, A.; Bui, T.V.; Tong, X.; Gruber, J.J.; Swider, C.R.; Sanchez, P.V.; Lum, J.J.; Sayed, N.; Melo, J.V.; et al. Imatinib resistance associated with BCR-ABL upregulation is dependent on HIF-1alpha-induced metabolic reprograming. Oncogene 2010, 29, 2962–2972. [Google Scholar] [CrossRef] [PubMed]
- Monteleone, F.; Rosa, R.; Vitale, M.; D’Ambrosio, C.; Succoio, M.; Formisano, L.; Nappi, L.; Romano, M.F.; Scaloni, A.; Tortora, G.; et al. Increased anaerobic metabolism is a distinctive signature in a colorectal cancer cellular model of resistance to antiepidermal growth factor receptor antibody. Proteomics 2013, 13, 866–877. [Google Scholar] [CrossRef] [PubMed]
- Ricciardelli, C.; Lokman, N.A.; Cheruvu, S.; Tan, I.A.; Ween, M.P.; Pyragius, C.E.; Ruszkiewicz, A.; Hoffmann, P.; Oehler, M.K. Transketolase is upregulated in metastatic peritoneal implants and promotes ovarian cancer cell proliferation. Clin. Exp. Metastasis 2015, 32, 441–455. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Wang, M. Knockdown of TKTL1 additively complements cisplatin-induced cytotoxicity in nasopharyngeal carcinoma cells by regulating the levels of NADPH and ribose-5-phosphate. Biomed. Pharmacother. 2017, 85, 672–678. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Biomarker | Healthy Cell | Function | Inhibition/Absence > Disease | Activation > Disease |
|---|---|---|---|---|
| DNaseX/Apo10 | endonuclease activity leading to 300 bp DNA fragments | executing final step of apoptosis leading to the elimination of unwanted cells/tumor cells | abnormal cell proliferation and inhibition of apoptosis > arise of tumor cells | |
| TKTL1 | energy release from carbohydrates, glucogenic amino acids and glycerin without generation of ROS | avoiding production of ROS and protection from ROS induced cell damages in particular in retina, testis and stem cells | high levels of ROS leading to cell damages, enhanced aging and premature cell death leading to
| activation in tumor cells leads to prevention and suppression of ROS, reduction of cytochrome c concomitant with increased malignity of tumor cells, inhibition of apoptosis and increased therapy resistance towards radiotherapy, many chemotherapies (e.g., platin derivatives) and targeted therapies like imatinib, sorafenib. |
| conversion/degradation of carbohydrates, glucogenic amino acids and glycerin to acetyl-CoA without loss of carbon atoms | generation of acetyl-CoA as the most important metabolite for synthesis of lipids for cell proliferation. TKTL1 dependent generation of acetyl-CoA is used for anabolic conditions, whereas pyruvatedehydrogenase dependent generation of acetyl-CoA is used for catabolic conditions and energy release by citric acid (Krebs) cycle | inhibition of TKTL1 by thiamine deficiency increase/induce chronic diabetes complications. Inhibition of TKTL1/TKT by oxythiamine created by certain cooking conditions leads to end-stage renal disease. | ||
| conversion/degradation of carbohydrates, glucogenic amino acids and glycerin to ribose and desoxyribose for DNA/mRNA synthesis and increased generation of NADPH by the oxidative part of the pentose phosphate pathway | generation of DNA and mRNA. Control of RedOx homeostasis by NADPH and glutathione. | enhanced DNA damage in healthy cells lead to premature aging | enhanced repair of DNA damages in tumor cells leads to therapy resistance towards radio- and many chemotherapies e.g., platin derivatives | |
| oxygen independent energy release and production of lactic acid even in the presence of oxygen and stabilization of HIF1α | cell survival under hypoxic conditions represents a survival mechanism for cells with no more access to blood oxygen caused by ischemia or infarct | absence of lactic acid based matrix degradation and invasive growth lead to inhibition of wound healing e.g., in diabetes patients | activation in tumor cells leads to a lactic acid based matrix degradation and concomitant to invasive growth and metastasis as well as resistance towards anti-angiogenic treatment e.g., avastin and erbitux | |
| activation in tumor cells leads lactic acid based inhibition of T and NK cells = protection from immune system attack | ||||
| confers cell survival and resistance to growth stimulus withdrawal e.g., hormone ablation | allows survival as single cell migrating in the body | resistance towards hormone ablation therapy e.g., androgen ablation therapy |
| (a) Pre- and postoperative epitope detection in monocytes (EDIM)-Apo10 and TKTL1 scores in patients with oral squamous cell carcinoma (n = 3). | |||||
| Characteristics | Preoperative | Postoperative | |||
| Patients | Total n = 3 | Apo10 score | TKTL1 score | Apo10 score | TKTL1 score |
| Patient 1 | 143 | 134 | 102 | 111 | |
| Patient 2 | 119 | 146 | 99 | 102 | |
| Patient 3 | 124 | 121 | 100 | 93 | |
| (b) Pre- and postoperative epitope detection in monocytes (EDIM)-Apo10 and TKTL1 scores in patients with breast cancer (n = 3). | |||||
| Characteristics | Preoperative | Postoperative | |||
| Patients | Total n = 3 | Apo10 score | TKTL1 score | Apo10 score | TKTL1 score |
| Patient 1 | 161 | 129 | 81 | 100 | |
| Patient 2 | 126 | 155 | 96 | 77 | |
| Patient 3 | 133 | 132 | 98 | 89 | |
| (c) Pre- and postoperative epitope detection in monocytes (EDIM)-Apo10 and TKTL1 scores in patients with prostate cancer (n = 6). | |||||
| Characteristics | Preoperative | Postoperative | |||
| Patients | Total n = 6 | Apo10 score | TKTL1 score | Apo10 score | TKTL1 score |
| Patient 1 | 155 | 149 | 98 | 99 | |
| Patient 2 | 144 | 166 | 93 | 106 | |
| Patient 3 | 153 | 165 | 93 | 101 | |
| Patient 4 | 162 | 143 | 105 | 104 | |
| Patient 5 | 139 | 149 | 102 | 88 | |
| Patient 6 | 158 | 144 | 95 | 95 | |
© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coy, J.F. EDIM-TKTL1/Apo10 Blood Test: An Innate Immune System Based Liquid Biopsy for the Early Detection, Characterization and Targeted Treatment of Cancer. Int. J. Mol. Sci. 2017, 18, 878. https://doi.org/10.3390/ijms18040878
Coy JF. EDIM-TKTL1/Apo10 Blood Test: An Innate Immune System Based Liquid Biopsy for the Early Detection, Characterization and Targeted Treatment of Cancer. International Journal of Molecular Sciences. 2017; 18(4):878. https://doi.org/10.3390/ijms18040878
Chicago/Turabian StyleCoy, Johannes F. 2017. "EDIM-TKTL1/Apo10 Blood Test: An Innate Immune System Based Liquid Biopsy for the Early Detection, Characterization and Targeted Treatment of Cancer" International Journal of Molecular Sciences 18, no. 4: 878. https://doi.org/10.3390/ijms18040878
APA StyleCoy, J. F. (2017). EDIM-TKTL1/Apo10 Blood Test: An Innate Immune System Based Liquid Biopsy for the Early Detection, Characterization and Targeted Treatment of Cancer. International Journal of Molecular Sciences, 18(4), 878. https://doi.org/10.3390/ijms18040878