
Heads or Tails: Genotyping of Hepatitis C Virus Concerning the 2k/1b Circulating Recombinant Form

 ,
,
Abstract
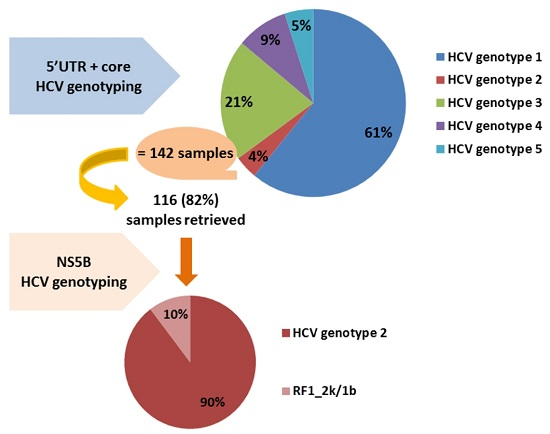
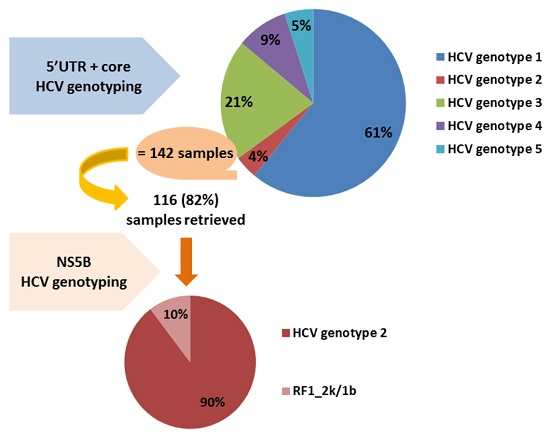
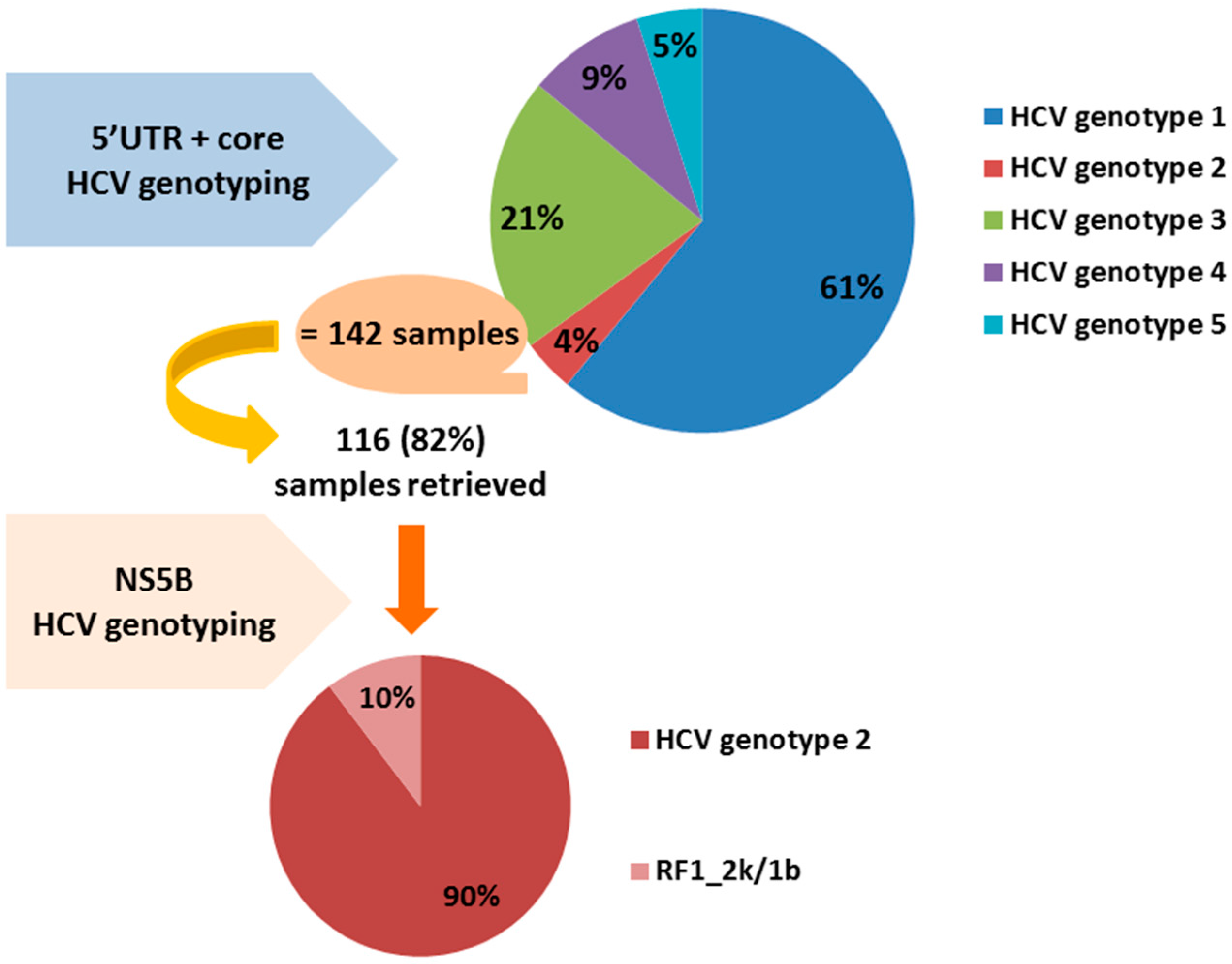
:
{kind=link}
{kind=link}
1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Serum Samples
4.2. RNA Extraction and cDNA Synthesis and Amplification
4.3. Reverse Hybridization
4.4. DNA Sequencing and Sequence Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- WHO. Hepatitis C. Available online: http://www.who.int/mediacentre/factsheets/fs164/en/ (accessed on 21 July 2015).
- Brook, G.; Soriano, V.; Bergin, C. European guideline for the management of hepatitis B and C virus infections, 2010. Int. J. STD AIDS 2010, 21, 669–678. [Google Scholar] [CrossRef] [PubMed]
- Pawlotsky, J.M.; Aghemo, A.; Back, D.; Dusheiko, G.; Forns, X.; Puoti, M.; Sarrazin, C. EASL recommendations on treatment of hepatitis C 2015. J. Hepatol. 2015, 63, 199–236. [Google Scholar]
- Farci, P.; Choo, Q.L.; Kuo, G.; Weiner, A.J.; Overby, L.R.; Bradley, D.W.; Houghton, M. Isolation of a cDNA clone derived from a blood-borne non-A, non-B viral hepatitis genome. J. Hepatol. 2015, 36, 582–585. [Google Scholar] [CrossRef]
- Simmonds, P.; Alberti, A.; Alter, H.J.; Bonino, F.; Bradley, D.W.; Brechot, C.; Brouwer, J.T.; Chan, S.W.; Chayama, K.; Chen, D.S.; et al. A proposed system for the nomenclature of hepatitis C viral genotypes. Hepatology 1994, 19, 1321–1324. [Google Scholar] [CrossRef] [PubMed]
- Penin, F.; Dubuisson, J.; Rey, F.A.; Moradpour, D.; Pawlotsky, J.-M. Structural biology of hepatitis C virus. Hepatology 2004, 39, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Chevaliez, S.; Pawlotsky, J.-M. HCV Genome and Life Cycle. In Hepatitis C Viruses: Genomes and Molecular Biology; Horizon Bioscience: Norfolk, UK, 2006. [Google Scholar]
- Li, H.-C.; Lo, S.-Y. Hepatitis C virus: Virology, diagnosis and treatment. World J. Hepatol. 2015, 7, 1377–1389. [Google Scholar] [CrossRef] [PubMed]
- Fishman, S.L.; Branch, A.D. The quasispecies nature and biological implications of the hepatitis C virus. Infect. Genet. Evol. 2009, 9, 1158–1167. [Google Scholar] [CrossRef] [PubMed]
- Kalinina, O.; Norder, H.; Mukomolov, S.; Magnius, O.; Magnius, L.O. A Natural intergenotypic recombinant of hepatitis C virus identified in St. Petersburg. Society 2002, 76, 4034–4043. [Google Scholar] [CrossRef]
- Zein, N.N. Clinical significance of hepatitis C virus genotypes. Clin. Microbiol. Rev. 2000, 13, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Markov, P.V.; Pepin, J.; Frost, E.; Deslandes, S.; Labbé, A.-C.; Pybus, O.G. Phylogeography and molecular epidemiology of hepatitis C virus genotype 2 in Africa. J. Gen. Virol. 2009, 90, 2086–2096. [Google Scholar] [CrossRef] [PubMed]
- Raghwani, J.; Thomas, X.V.; Koekkoek, S.M.; Schinkel, J.; Molenkamp, R.; van de Laar, T.J.; Takebe, Y.; Tanaka, Y.; Mizokami, M.; Rambaut, A.; et al. Origin and evolution of the unique hepatitis C virus circulating recombinant form 2k/1b. J. Virol. 2012, 86, 2212–2220. [Google Scholar] [CrossRef] [PubMed]
- Starr, D. Blood: An Epic History of Medicine and Commerce, 2000. Available online: https://books.google.be/books/about/Blood.html?id=pa5BAWw6L1oC&pgis=1 (accessed on 26 September 2015).
- Alter, M.J. HCV routes of transmission: What goes around comes around. Semin. Liver Dis. 2011, 31, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Moreau, I.; Hegarty, S.; Levis, J.; Sheehy, P.; Crosbie, O.; Kenny-Walsh, E.; Fanning, L.J. Serendipitous identification of natural intergenotypic recombinants of hepatitis C in Ireland. Virol. J. 2006, 3, 95. [Google Scholar] [CrossRef] [PubMed]
- Tallo, T.; Norder, H.; Tefanova, V.; Krispin, T.; Schmidt, J.; Ilmoja, M.; Orgulas, K.; Pruunsild, K.; Priimägi, L.; Magnius, L.O. Genetic characterization of hepatitis C virus strains in Estonia: Fluctuations in the predominating subtype with time. J. Med. Virol. 2007, 79, 374–382. [Google Scholar] [CrossRef] [PubMed]
- Kurbanov, F.; Tanaka, Y.; Avazova, D.; Khan, A.; Sugauchi, F.; Kan, N.; Kurbanova-Khudayberganova, D.; Khikmatullaeva, A.; Musabaev, E.; Mizokami, M. Detection of hepatitis C virus natural recombinant RF1_2k/1b strain among intravenous drug users in Uzbekistan. Hepatol. Res. 2008, 38, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Demetriou, V.L.; Kyriakou, E.; Kostrikis, L.G. Near-full genome characterisation of two natural intergenotypic 2k/1b recombinant hepatitis C virus isolates. Adv. Virol. 2011. [Google Scholar] [CrossRef] [PubMed]
- Ramière, C.; Tremeaux, P.; Caporossi, A.; Trabaud, M.; Lebossé, F.; Bailly, F.; Thélu, M.A.; Nana, J.; Leroy, V.; Morand, P.; et al. Recent evidence of underestimated circulation of hepatitis C virus intergenotypic recombinant strain RF2k/1b in the Rhône-Alpes region, France, January to August 2014: Implications for antiviral treatment. Eurosurveillance 2014, 19, 20944. [Google Scholar] [CrossRef] [PubMed]
- De Keukeleire, S.; Descheemaeker, P.; Reynders, M. Potential risk of misclassification HCV 2k/1b strains as HCV 2a/2c using VERSANT HCV Genotype 2.0 assay. Diagn. Microbiol. Infect. Dis. 2015, 82, 201–202. [Google Scholar] [CrossRef] [PubMed]
- Avó, A.P.; Agua-Doce, I.; Andrade, A.; Pádua, E. Hepatitis C virus subtyping based on sequencing of the C/E1 and NS5B genomic regions in comparison to a commercially available line probe assay. J. Med. Virol. 2013, 85, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Susser, V.P.S.; Dietz, J.; Barak, M.; Schlevogt, B.; Daniel, R.; Passmann, S.Z.S.; Cornberg, M.; Zuckerman, E.; Mangia, A.; Sarrazin, C. Prevalence and clinical importance of hepatitis C virus genotype 2k/1b chimeras. J. Hepatol. 2016, 64, S136. [Google Scholar]
- Verbeeck, J.; Stanley, M.J.; Shieh, J.; Celis, L.; Huyck, E.; Wollants, E.; Morimoto, J.; Farrior, A.; Sablon, E.; Jankowski-Hennig, M.; et al. Evaluation of versant hepatitis C virus genotype assay (LiPA) 2.0. J. Clin. Microbiol. 2008, 46, 1901–1906. [Google Scholar] [CrossRef] [PubMed]
- Messina, J.P.; Humphreys, I.; Flaxman, A.; Brown, A.; Cooke, G.S.; Pybus, O.G.; Barnes, E. Global distribution and prevalence of hepatitis C virus genotypes. Hepatology 2014, 61, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Morel, V.; Fournier, C.; François, C.; Brochot, E.; Helle, F.; Duverlie, G.; Castelain, S. Genetic recombination of the hepatitis C virus: Clinical implications. J. Viral Hepat. 2011, 18, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Ruta, S.; Cernescu, C. Injecting drug use: A vector for the introduction of new hepatitis C virus genotypes. World J. Gastroenterol. 2015, 21, 10811–10823. [Google Scholar] [CrossRef] [PubMed]
- Kurbanov, F.; Tanaka, Y.; Chub, E.; Maruyama, I.; Azlarova, A.; Kamitsukasa, H.; Ohno, T.; Bonetto, S.; Moreau, I.; Fanning, L.J.; et al. Molecular epidemiology and interferon susceptibility of the natural recombinant hepatitis C virus strain RF1_2k/1b. J. Infect. Dis. 2008, 198, 1448–1456. [Google Scholar] [CrossRef] [PubMed]
- Morel, V.; Descamps, V.; François, C.; Fournier, C.; Brochot, E.; Capron, D.; Duverlie, G.; Castelain, S. Emergence of a genomic variant of the recombinant 2k/1b strain during a mixed Hepatitis C infection: A case report. J. Clin. Virol. 2010, 47, 382–386. [Google Scholar] [CrossRef] [PubMed]
- Hedskog, C.; Doehle, B.; Chodavarapu, K.; Gontcharova, V.; Garcia, J.C.; de Knegt, R.; Drenth, J.P.; McHutchison, J.G.; Brainard, D.; Stamm, L.M.; et al. Characterization of hepatitis C virus intergenotypic recombinant strains and associated virological response to sofosbuvir/ribavirin. Hepatology 2015, 61, 471–480. [Google Scholar] [CrossRef] [PubMed]
- EASL. Recommendations on Treatment of Hepatitis C 2015. J. Hepatol. 2015, 63, 199–236. [Google Scholar]
- Murphy, D.G.; Willems, B.; Deschênes, M.; Hilzenrat, N.; Mousseau, R.; Sabbah, S. Use of sequence analysis of the NS5B region for routine genotyping of hepatitis C virus with reference to C/E1 and 5′ untranslated region sequences. J. Clin. Microbiol. 2007, 45, 1102–1112. [Google Scholar] [CrossRef] [PubMed]
- Sanger, F. Determination of nucleotide sequences in DNA. Biosci. Rep. 2004, 24, 237. [Google Scholar] [CrossRef] [PubMed]

© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schuermans, W.; Orlent, H.; Desombere, I.; Descheemaeker, P.; Van Vlierberghe, H.; Geerts, A.; Verhelst, X.; Reynders, M.; Padalko, E. Heads or Tails: Genotyping of Hepatitis C Virus Concerning the 2k/1b Circulating Recombinant Form. Int. J. Mol. Sci. 2016, 17, 1384. https://doi.org/10.3390/ijms17091384
Schuermans W, Orlent H, Desombere I, Descheemaeker P, Van Vlierberghe H, Geerts A, Verhelst X, Reynders M, Padalko E. Heads or Tails: Genotyping of Hepatitis C Virus Concerning the 2k/1b Circulating Recombinant Form. International Journal of Molecular Sciences. 2016; 17(9):1384. https://doi.org/10.3390/ijms17091384
Chicago/Turabian StyleSchuermans, Wim, Hans Orlent, Isabelle Desombere, Patrick Descheemaeker, Hans Van Vlierberghe, Anja Geerts, Xavier Verhelst, Marijke Reynders, and Elizaveta Padalko. 2016. "Heads or Tails: Genotyping of Hepatitis C Virus Concerning the 2k/1b Circulating Recombinant Form" International Journal of Molecular Sciences 17, no. 9: 1384. https://doi.org/10.3390/ijms17091384