
Comprehensive Identification of Immunodominant Proteins of Brucella abortus and Brucella melitensis Using Antibodies in the Sera from Naturally Infected Hosts


Abstract
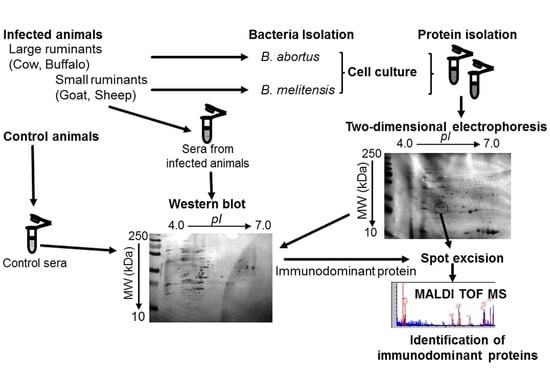
:
1. Introduction
2. Results
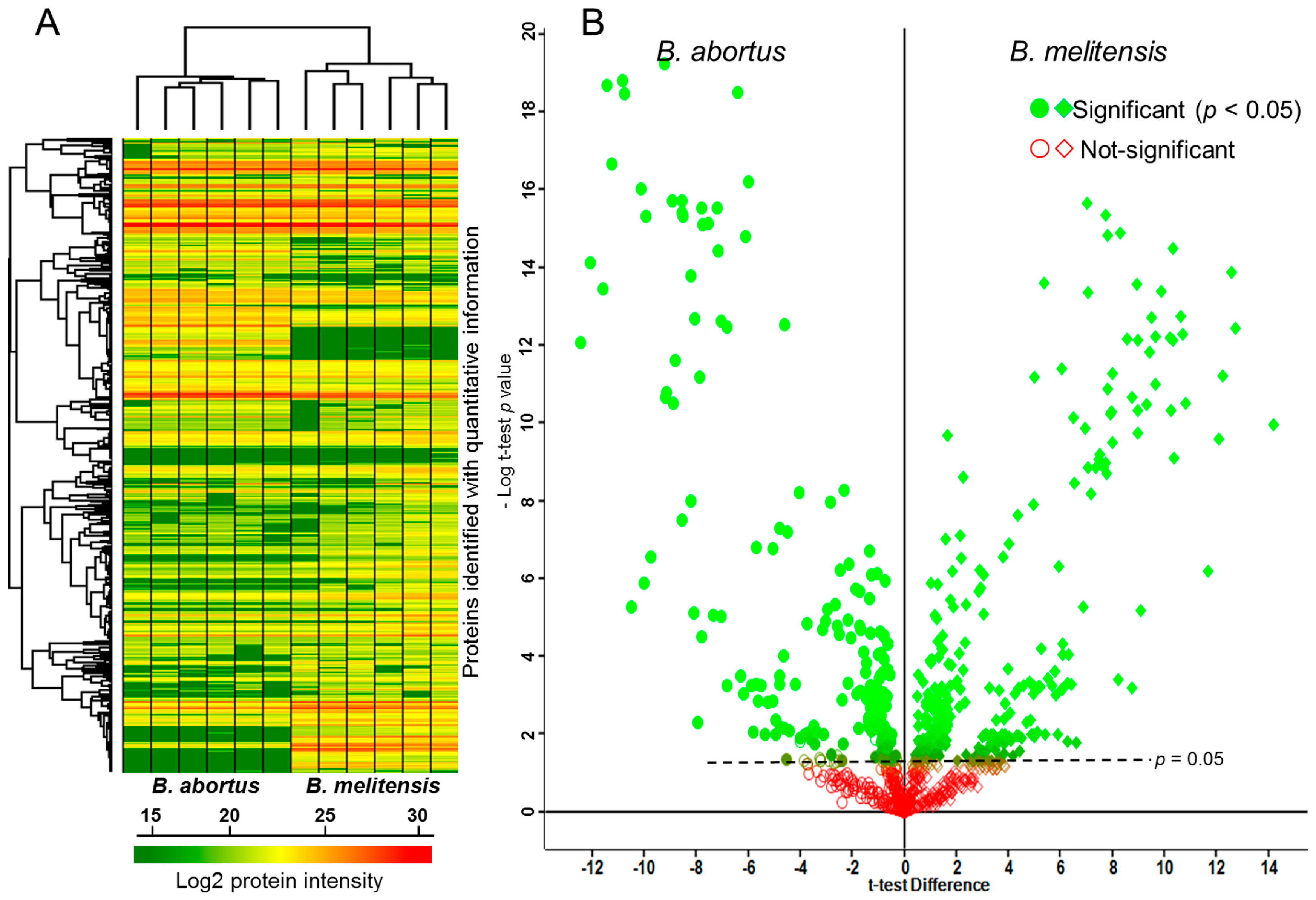
2.1. Comparative Proteomics Analysis of Brucella (B.) abortus and B. melitensis Field Strains
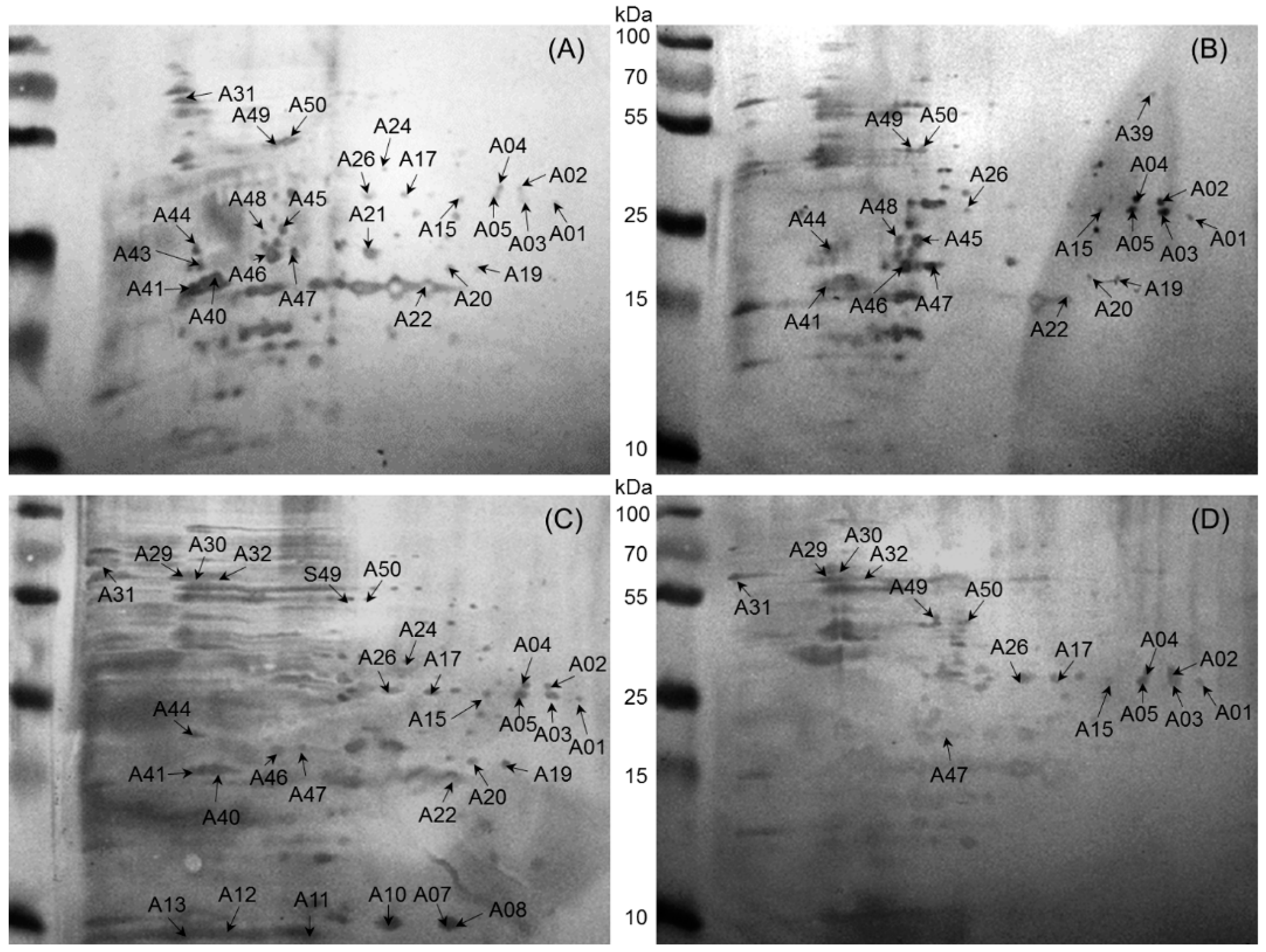
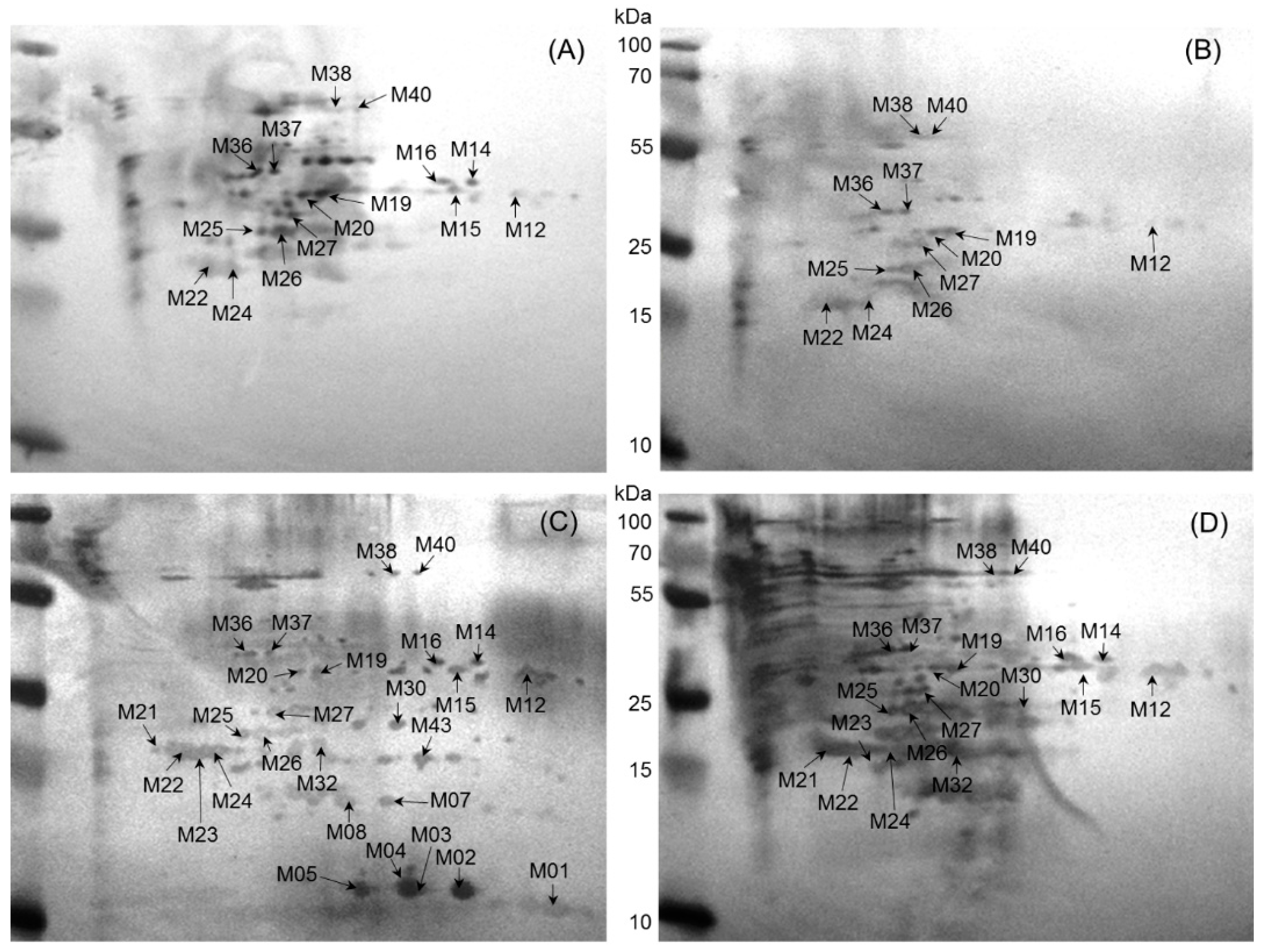
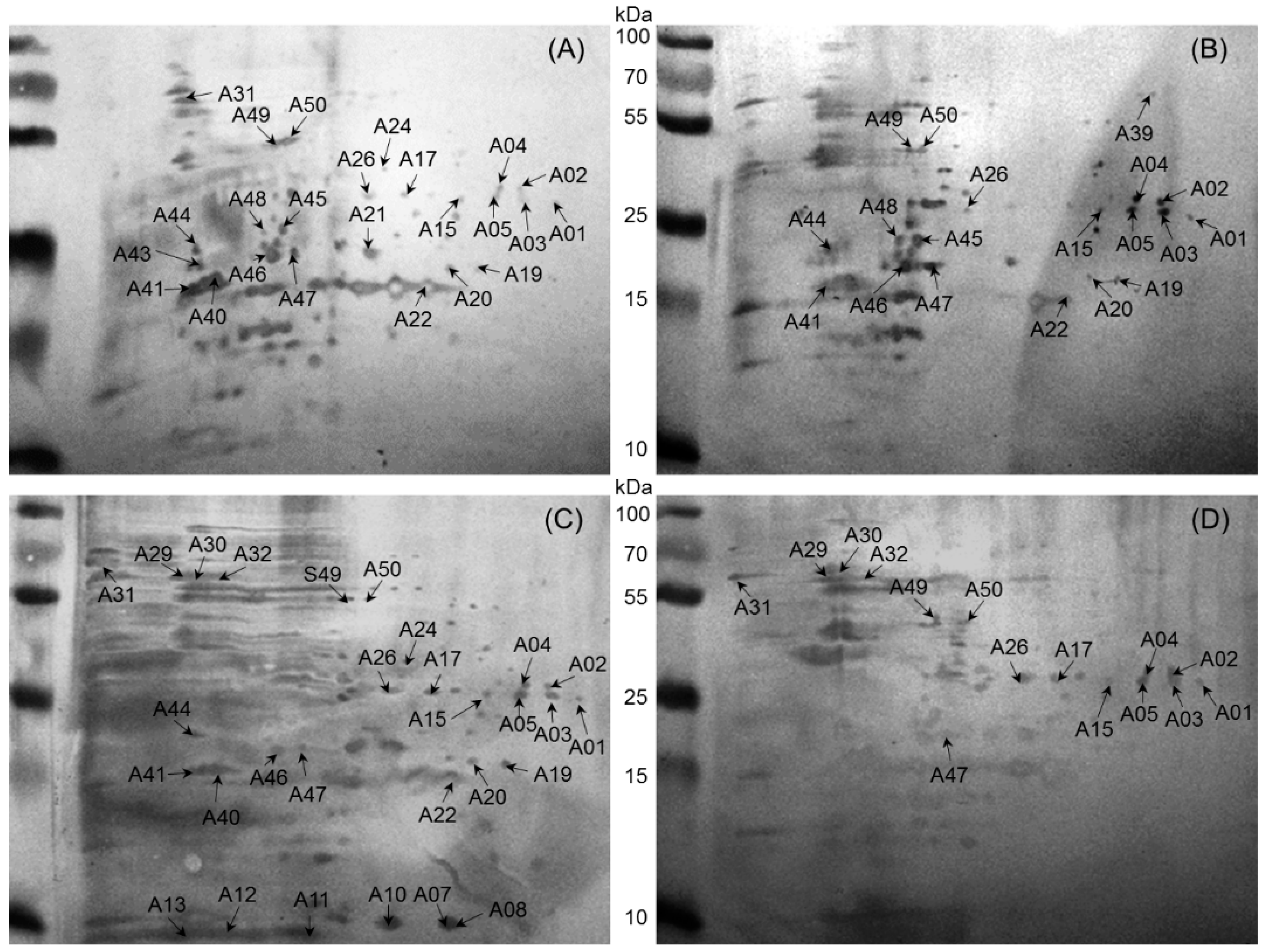
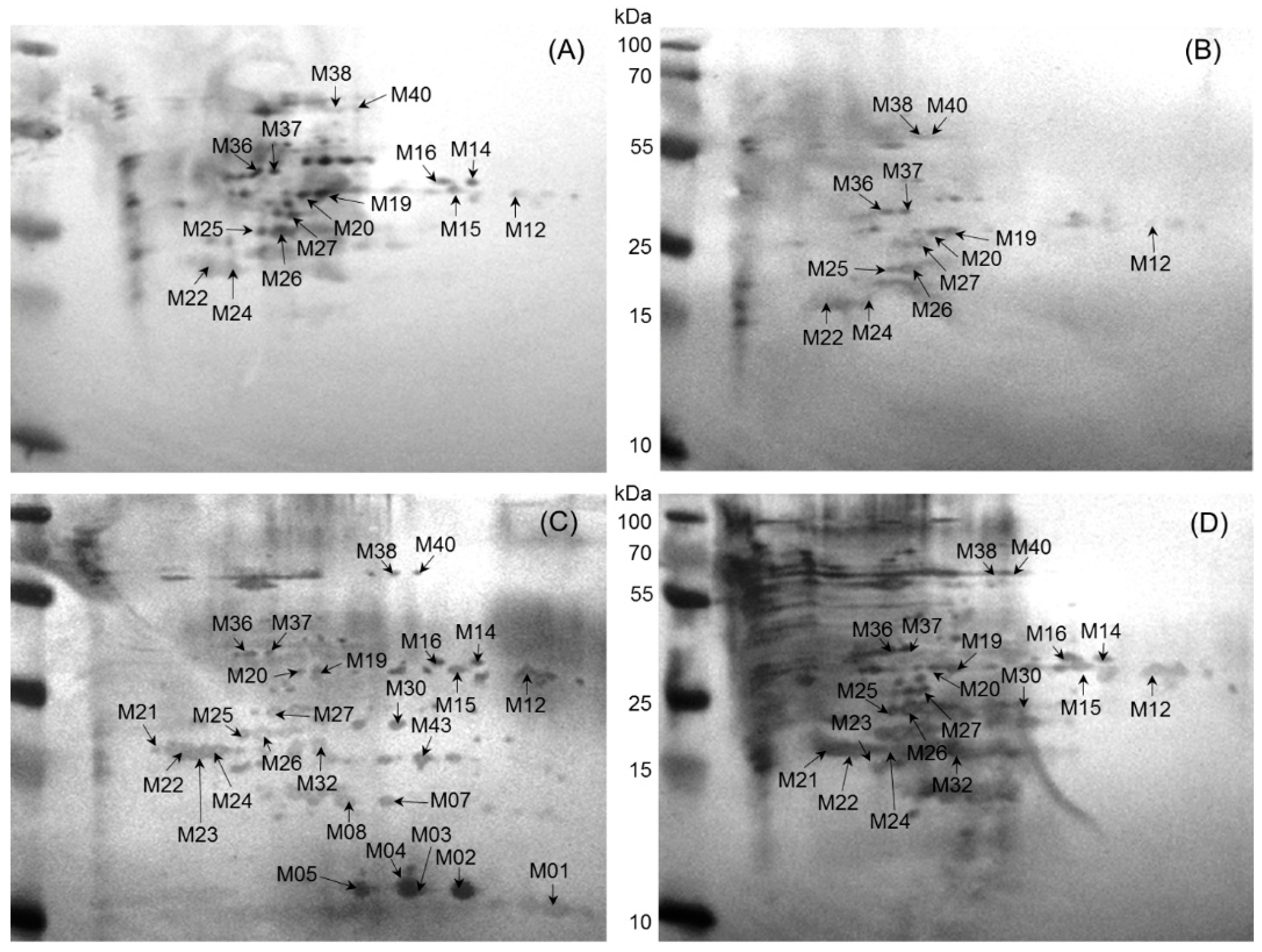
2.2. Immunoreactive Proteins of B. abortus
2.3. Immunoreactive Proteins of B. melitensis
2.4. Identification of Cross-Reactive Proteins between B. abortus and B. melitensis
2.5. Comparative Basic Local Alignment Search Tool (BLAST) Analysis
3. Discussion
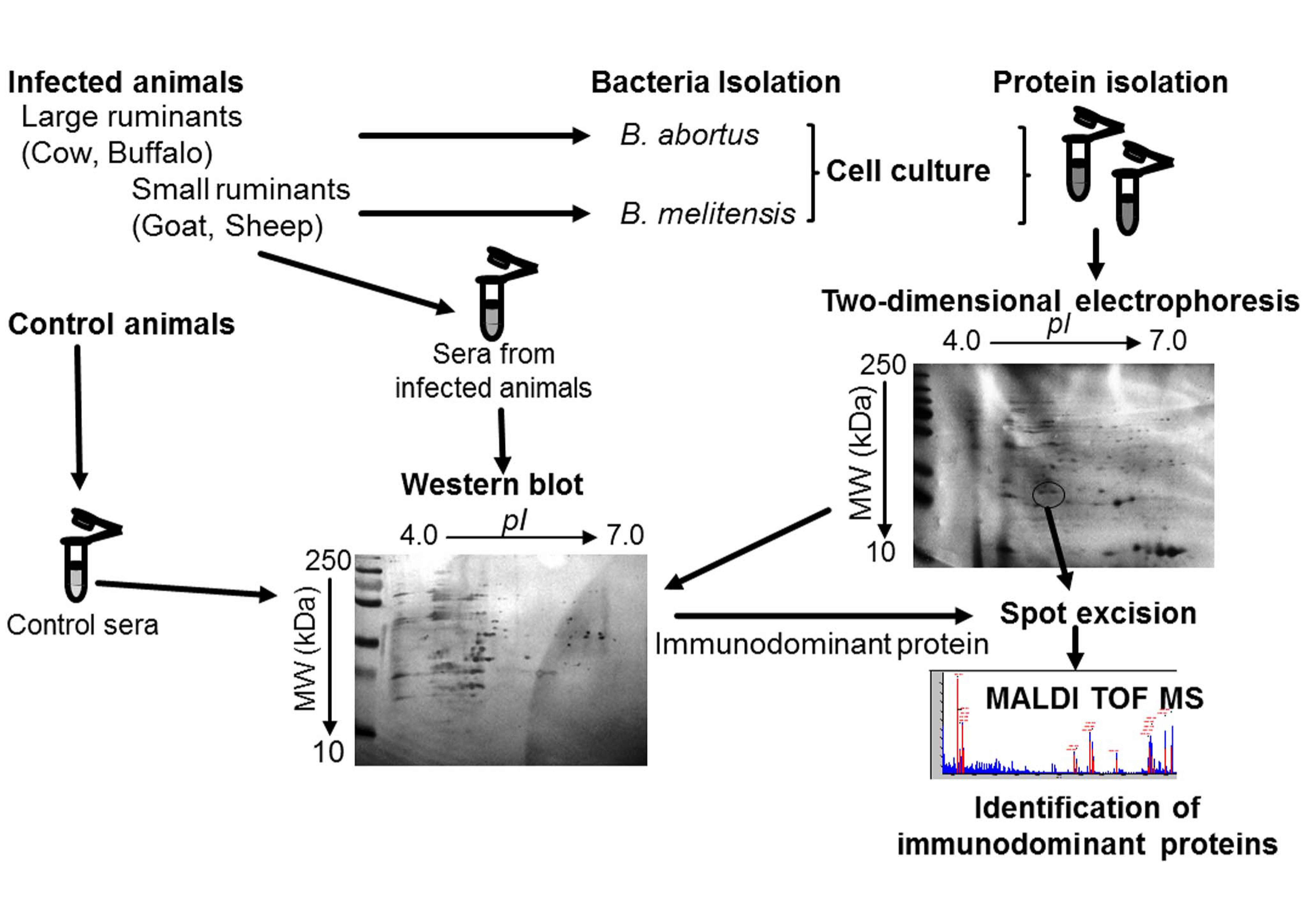
4. Materials and Methods
4.1. Bacterial Strains and Sera Selection
4.2. Ethics Statement
4.3. Cell Culture and Protein Extraction
4.4. In-Solution Trypsin Digestion
4.5. Liquid Chromatography-Electrospray Ionization-Tandem Mass Spectrometry (LC-ESI-MS/MS)
4.6. Protein Identification and Data Analysis
4.7. Two-Dimensional Electrophoresis
4.8. 2-D-PAGE Western Blotting
4.9. In-Gel Trypsin Digestion and Matrix-Assisted Laser Desorption/ionization-Tine of Flight-Mass Spectrometry (MALDI-TOF MS/MS)
4.10. Comparison of the Identified Proteins and Other Cross-Reactive Bacteria
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cutler, S.J.; Whatmore, A.M.; Commander, N.J. Brucellosis—New aspects of an old disease. J. Appl. Microbiol. 2005, 98, 1270–1281. [Google Scholar]
- Halling, S.M.; Peterson-Burch, B.D.; Bricker, B.J.; Zuerner, R.L.; Qing, Z.; Li, L.L.; Kapur, V.; Alt, D.P.; Olsen, S.C. Completion of the genome sequence of Brucella abortus and comparison to the highly similar genomes of Brucella melitensis and Brucella suis. J. Bacteriol. 2005, 187, 2715–2726. [Google Scholar] [CrossRef] [PubMed]
- Eschenbrenner, M.; Horn, T.A.; Wagner, M.A.; Mujer, C.V.; Miller-Scandle, T.L.; DelVecchio, V.G. Comparative proteome analysis of laboratory grown Brucella abortus 2308 and Brucella melitensis 16 m. J. Proteome Res. 2006, 5, 1731–1740. [Google Scholar] [CrossRef] [PubMed]
- Galińska, E.; Zagórski, J. Brucellosis in humans—Etiology, diagnostics, clinical forms. Ann. Agric. Environ. Med. 2013, 20, 233–238. [Google Scholar] [PubMed]
- Diaz, A.E. Epidemiology of brucellosis in domestic animals caused by Brucella melitensis, Brucella suis and Brucella abortus. Rev. Sci. Tech. 2013, 32, 43–51, 53–60. [Google Scholar]
- Alton, G.G.; Jones, L.M.; Angus, R.D.; Verger, J.M. Techniques for the Brucellosis Laboratory; Instituttional de la Recherche Agronomique: Paris, France, 1988; pp. 17–62. [Google Scholar]
- Al Dahouk, S.; Nöckler, K.; Scholz, H.C.; Tomaso, H.; Bogumil, R.; Neubauer, H. Immunoproteomic characterization of Brucella abortus 1119-3 preparations used for the serodiagnosis of Brucella infections. J. Immunol. Methods 2006, 309, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Samartino, L.; Gall, D.; Gregoret, R.; Nielsen, K. Validation of enzyme-linked immunosorbent assays for the diagnosis of bovine brucellosis. Vet. Microbiol. 1999, 70, 193–200. [Google Scholar] [CrossRef]
- Von Bargen, K.; Gorvel, J.P.; Salcedo, S.P. Internal affairs: Investigating the Brucella intracellular lifestyle. FEMS Microbiol. Rev. 2012, 36, 533–562. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate—A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar]
- Rosinha, G.; Myioshi, A.; Azevedo, V.; Splitter, G.; Oliveira, S. Molecular and immunological characterisation of recombinant Brucella abortus glyceraldehyde-3-phosphate-dehydrogenase, a T- and B-cell reactive protein that induces partial protection when co-administered with an interleukin-12-expressing plasmid in a DNA vaccine formulation. J. Med. Microbiol. 2002, 51, 661–671. [Google Scholar] [PubMed]
- Yang, Y.; Wang, L.; Yin, J.; Wang, X.; Cheng, S.; Lang, X.; Wang, X.; Qu, H.; Sun, C.; Wang, J.; et al. Immunoproteomic analysis of Brucella melitensis and identification of a new immunogenic candidate protein for the development of brucellosis subunit vaccine. Mol. Immunol. 2011, 49, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Al-Dahouk, S.; Loisel-Meyer, S.V.; Scholz, H.C.; Tomaso, H.; Kersten, M.; Harder, A.; Neubauer, H.; Köhler, S.; Jubier-Maurin, V.R. Proteomic analysis of Brucella suis under oxygen deficiency reveals flexibility in adaptive expression of various pathways. Proteomics 2009, 9, 3011–3021. [Google Scholar] [CrossRef] [PubMed]
- Eschenbrenner, M.; Wagner, M.A.; Horn, T.A.; Kraycer, J.A.; Mujer, C.V.; Hagius, S.; Elzer, P.; DelVecchio, V.G. Comparative proteome analysis of Brucella melitensis vaccine strain rev 1 and a virulent strain, 16 M. J. Bacteriol. 2002, 184, 4962–4970. [Google Scholar] [CrossRef] [PubMed]
- Del Vecchio, V.G.; Wagner, M.A.; Eschenbrenner, M.; Horn, T.; Kraycer, J.A.; Estock, F.; Elzer, P.; Mujer, C.V. Brucella proteomes—A review. Vet. Microbiol. 2002, 90, 593–603. [Google Scholar] [CrossRef]
- Connolly, J.P.; Comerci, D.; Alefantis, T.G.; Walz, A.; Quan, M.; Chafin, R.; Grewal, P.; Mujer, C.V.; Ugalde, R.A.; DelVecchio, V.G. Proteomic analysis of Brucella abortus cell envelope and identification of immunogenic candidate proteins for vaccine development. Proteomics 2006, 6, 3767–3780. [Google Scholar] [CrossRef] [PubMed]
- Teixeira-Gomes, A.; Cloeckaert, A.; Bézard, G.; Bowden, R.A.; Dubray, G.; Zygmunt, M.S. Identification and characterization of Brucella ovis immunogenic proteins using two-dimensional electrophoresis and immunoblotting. Electrophoresis 1997, 18, 1491–1497. [Google Scholar] [CrossRef] [PubMed]
- Teixeira-Gomes, A.; Cloeckaert, A.; Bézard, G.; Dubray, G.; Zygmunt, M.S. Mapping and identification of Brucella melitensis proteins by two-dimensional electrophoresis and microsequencing. Electrophoresis 1997, 18, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Ko, K.Y.; Kim, J.W.; Her, M.; Kang, S.I.; Jung, S.C.; Cho, D.H.; Kim, J.Y. Immunogenic proteins of Brucella abortus to minimize cross-reactions in brucellosis diagnosis. Vet. Microbiol. 2012, 156, 374–380. [Google Scholar] [CrossRef] [PubMed]
- Olsen, S.C.; Stoffregen, W.S. Essential role of vaccines in brucellosis control and eradication programs for livestock. Expert Rev. Vaccines 2005, 4, 915–928. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, K.; Smith, P.; Widdison, J.; Gall, D.; Kelly, L.; Kelly, W.; Nicoletti, P. Serological relationship between cattle exposed to Brucella abortus, Yersinia enterocolitica O:9 and Escherichia coli O157:H7. Vet. Microbiol. 2004, 100, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Yan, F.; Ji, W.; Luo, D.; Liu, X.; Xing, L.; Duan, Y.; Yang, P.; Shi, X.; Li, Z.; et al. Identification of immunoreactive proteins of Brucella melitensis by immunoproteomics. Sci. China Life Sci. 2011, 54, 880–887. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.J.; Simborio, H.L.; Reyes, A.W.; Kim, D.G.; Hop, H.T.; Min, W.; Her, M.; Jung, S.C.; Yoo, H.S.; Kim, S. Immunoproteomic identification of immunodominant antigens independent of the time of infection in Brucella abortus 2308-challenged cattle. Vet. Res. 2015, 46, 17. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Sung, S.R.; Lee, K.; Lee, H.K.; Kang, S.I.; Lee, J.J.; Jung, S.C.; Park, Y.H.; Her, M. Immunoproteomics of brucella abortus RB51 as candidate antigens in serological diagnosis of brucellosis. Vet. Immunol. Immunopathol. 2014, 160, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Skovpen, Y.; Palmer, D. Dihydrodipicolinate synthase from Campylobacter jejuni: Kinetic mechanism of cooperative allosteric inhibition and inhibitor-induced substrate cooperativity. Biochemistry 2013, 52, 5454–5462. [Google Scholar] [CrossRef] [PubMed]
- Fugier, E.; Salcedo, S.; de Chastellier, C.; Pophillat, M.; Muller, A.; Arce-Gorvel, V.; Fourquet, P.; Gorvel, J. The glyceraldehyde-3-phosphate dehydrogenase and the small GTPase Rab 2 are crucial for Brucella replication. PLoS Pathog. 2009, 5, e1000487. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Tong, Y.; Tian, M.; Sun, X.; Wang, S.; Ding, C.; Yu, S. Characterization of the immunogenicity and pathogenicity of malate dehydrogenase in Brucella abortus. World J. Microbiol. Biotechnol. 2014, 30, 2063–2070. [Google Scholar] [CrossRef] [PubMed]
- Tomm, J.; Krause, C.; Simon, J.; Treudler, R.; von Bergen, M.; Averbeck, M. Pyruvate kinase and phosphopyruvate hydratase as novel IGE reactive proteins in prawn. J. Investig. Allergol. Clin. Immunol. 2013, 23, 443–445. [Google Scholar] [PubMed]
- Kim, G.J.; Kim, H.S. Identification of the structural similarity in the functionally related amidohydrolases acting on the cyclic amide ring. Biochem. J. 1998, 330, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Schneider, E.; Hunke, S. ATP-binding-cassette (ABC) transport systems: Functional and structural aspects of the ATP-hydrolyzing subunits/domains. FEMS Microbiol. Rev. 1998, 22, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Kittelberger, R.; Reichel, M.; Joyce, M.; Staak, C. Serological crossreactivity between Brucella abortus and Yersinia enterocolitica 0:9. III. Specificity of the in vitro antigen-specific γ interferon test for bovine brucellosis diagnosis in experimentally Yersinia enterocolitica 0:9-infected cattle. Vet. Microbiol. 1997, 57, 361–371. [Google Scholar] [CrossRef]
- Wu, A.M.; MacKenzie, N.E.; Adams, G.; Pugh, R. Structural and immunochemical aspects of Brucella abortus endotoxins. Adv. Exp. Med. Biol. 1988, 228, 551–576. [Google Scholar] [PubMed]
- Sowa, B.A.; Crawforda, R.P.; Heck, F.C.; Williams, J.D.; Wu, A.M.; Kelly, K.A.; Adams, L.G. Size, charge and structural heterogeneity of Brucella abortus lipopolysaccharides demonstrated by twodimensional gel electrophoresis. Electrophoresi 1986, 7, 283–288. [Google Scholar] [CrossRef]
- Palmer, D.A.; Douglas, J.T. Analysis of Brucella lipopolysaccharide with specific and cross-reacting monoclonal antibodies. J. Clin. Microbiol. 1989, 27, 2331–2337. [Google Scholar] [PubMed]
- Probert, W.S.; Schrader, K.N.; Khuong, N.Y.; Bystrom, S.L.; Graves, M.H. Real-time multiplex PCR assay for detection of Brucella spp., B. abortus, and B. melitensis. J. Clin. Microbiol. 2004, 42, 1290–1293. [Google Scholar] [CrossRef] [PubMed]
- Murugaiyan, J.; Rockstroh, M.; Wagner, J.; Baumann, S.; Schorsch, K.; Trump, S.; Lehmann, I.; Bergen, M.V.; Tomm, J.M. Benzo[a]pyrene affects jurkat T cells in the activated state via the antioxidant response element dependent Nrf2 pathway leading to decreased IL-2 secretion and redirecting glutamine metabolism. Toxicol. Appl. Pharmacol. 2013, 269, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Markwell, M.A.; Haas, S.M.; Bieber, L.L.; Tolbert, N.E. A modification of the lowry procedure to simplify protein determination in membrane and lipoprotein samples. Anal. Biochem. 1978, 87, 206–210. [Google Scholar] [CrossRef]
- Rappsilber, J.; Ishihama, Y.; Mann, M. Stop and go extraction tips for matrix-assisted laser desorption/ionization, nanoelectrospray, and LC/MS sample pretreatment in proteomics. Anal. Chem. 2003, 75, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.; Neuhauser, N.; Michalski, A.; Scheltema, R.A.; Olsen, J.V.; Mann, M. Andromeda: A peptide search engine integrated into the maxquant environment. J Proteome Res. 2011, 10, 1794–1805. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.; Hein, M.Y.; Luber, C.A.; Paron, I.; Nagaraj, N.; Mann, M. Accurate proteome-wide label-free quantification by delayed normalization and maximal peptide ratio extraction, termed maxlfq. Mol. Cell. Proteom. 2014, 13, 2513–2526. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.; Matic, I.; Hilger, M.; Nagaraj, N.; Selbach, M.; Olsen, J.V.; Mann, M. A practical guide to the maxquant computational platform for silac-based quantitative proteomics. Nat. Protoc. 2009, 4, 698–705. [Google Scholar] [CrossRef] [PubMed]
- Murugaiyan, J.; Weise, C.; von Bergen, M.; Roesler, U. Two-dimensional proteome reference map of Prototheca zopfii revealed reduced metabolism and enhanced signal transduction as adaptation to an infectious life style. Proteomics 2013, 13, 2664–2669. [Google Scholar] [CrossRef] [PubMed]
- Candiano, G.; Ruschi, M.; Musante, L.; Santucci, L.; Ghiggeri, G.; Carnemolla, B.; Orecchia, P.; Zardi, L.; Righetti, P. Blue silver: A very sensitive colloidal coomassie G-250 staining for proteome analysis. Electrophoresis 2004, 25, 1327–1333. [Google Scholar] [CrossRef] [PubMed]
- Wareth, G.; Melzer, F.; Weise, C.; Neubauer, H.; Roesler, U.; Murugaiyan, J. Proteomics-based identification of immunodominant proteins of brucellae using sera from infected hosts points towards enhanced pathogen survival during the infection. Biochem. Biophys. Res. Commun. 2015, 456, 202–206. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Spot ID | Acc.ID | Protein | Mw | MOWSE Score a | pI | Sequence Coverage (%) | No. of Peptides Matching | Host | Reference/Cross-Matching Spot ID |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | A01 | gi|256369084 | Dihydrodipicolinate synthase | 31,892 | 244 | 6.26 | 67 | 13 | Cow, Buffalo, Sheep, Goat | |
| A03 | gi|256369084 | Dihydrodipicolinate synthase | 31,892 | 426 | 6.26 | 67 | 13 | Cow, Buffalo, Sheep, Goat | ||
| A05 | gi|493692811 | Dihydrodipicolinate synthase | 33,539 | 132 | 7.08 | 68 | 14 | Cow, Buffalo, Sheep, Goat | ||
| A15 | gi|495149454 | Dihydrodipicolinate synthase | 31,753 | 75 | 5.94 | 28 | 5 | Cow, Buffalo, Sheep, Goat | ||
| 2 | A02 | gi|496823699 | Glyceraldehyde-3-phosphate dehydrogenase | 36,385 | 356 | 6.26 | 48 | 13 | Cow, Buffalo, Sheep, Goat | [10] |
| A04 | gi|4165122 | Glyceraldehyde-3-phosphate dehydrogenase | 36,344 | 107 | 5.89 | 38 | 8 | Cow, Buffalo, Sheep, Goat | [10] | |
| 3 | A26 | gi|226887955 | Lactate malate dehydrogenase | 34,152 | 243 | 5.24 | 31 | 5 | Cow, Buffalo, Sheep, Goat | |
| 4 | A47 | gi|493015116 | Hypothetical protein (fumarylacetoacetate hydrolase family protein) | 29,383 | 343 | 5.09 | 36 | 7 | Cow, Buffalo, Sheep, Goat | M25 |
| 5 | A49 | gi|17987134 | Phosphopyruvate hydratase | 45,462 | 421 | 4.99 | 53 | 18 | Cow, Buffalo, Sheep, Goat | |
| A50 | gi|148560469 | Phosphopyruvate hydratase | 45,431 | 494 | 5.03 | 47 | 16 | Cow, Buffalo, Sheep, Goat | ||
| 6 | A20 | gi|148558534 | Metal-dependent hydrolase | 25,257 | 103 | 5.58 | 44 | 7 | Cow, Buffalo, Sheep | |
| 7 | A17 | gi|82700282 | Choloylglycine hydrolase | 36,868 | 108 | 5.62 | 29 | 9 | Cow, Sheep, Goat | |
| 8 | A22 | gi|490830157 | Hydrolase | 27,731 | 134 | 6.07 | 50 | 8 | Cow, Buffalo, Sheep | [11] M43 |
| A19 | gi|490830157 | Hydrolase | 27,731 | 383 | 6.07 | 48 | 8 | Cow, Buffalo, Sheep | [11] | |
| 9 | A44 | gi|320161003 | Putative DNA processing protein | 40,919 | 56 | 5.85 | 42 | 7 | Cow, Buffalo, Sheep | |
| 10 | A46 | gi|489055332 | 2-Hydroxyhepta-2,4-diene-1,7-dioate isomerase | 30,092 | 380 | 5.08 | 48 | 8 | Cow, Buffalo, Sheep | |
| 11 | A41 | gi|493691811 | Sugar ABC transporter substrate-binding protein | 33,258 | 440 | 5.11 | 52 | 11 | Cow, Buffalo, Sheep | M36 |
| 12 | A45 | gi|384211119 | Lysine-arginine-ornithine-binding periplasmic protein | 36,684 | 440 | 5.09 | 57 | 15 | Cow, Buffalo | M27 |
| 13 | A48 | gi|152013695 | ADP/ATP translocase | 20,876 | 63 | 9.63 | 31 | 4 | Cow, Buffalo | |
| 14 | A39 | gi|62317242 | Urocanate hydratase | 61,589 | 173 | 6.04 | 19 | 11 | Buffalo, Sheep | |
| 15 | A24 | gi|493147262 | Sulfate transporter subunit | 37,727 | 132 | 5.92 | 38 | 11 | Cow, Sheep | |
| 16 | A31 | gi|493053174 | Catalase | 55,556 | 223 | 6.62 | 36 | 18 | Cow, Goat | |
| 17 | A40 | gi|17988780 | d-ribose-binding periplasmic protein precursor | 31,030 | 193 | 5.60 | 57 | 9 | Cow, Sheep | [12,13,14] M24 |
| 18 | A29 | gi|148558491 | Chaperonin GroEL | 57,505 | 99 | 5.08 | 18 | 7 | Sheep, Goat | [7,15,16,17] |
| A30 | gi|14855849 | Chaperonin GroEL | 57,505 | 92 | 5.08 | 20 | 9 | Sheep, Goat | [7,15,16,17] | |
| 19 | A32 | gi|144108 | Heat shock protein | 57,534 | 94 | 5.33 | 27 | 12 | Sheep, Goat | |
| 20 | A43 | gi|492987884 | Protein grpE | 24,883 | 128 | 4.70 | 40 | 12 | Cow | |
| 21 | A07 | gi|384446825 | Superoxide dismutase, copper/zinc binding protein | 17,255 | 370 | 6.10 | 64 | 7 | Sheep | [7,16,17,18] M01 |
| A8 | gi|489058379 | Superoxide dismutase copper/zinc binding protein | 18,205 | 242 | 6.24 | 54 | 5 | Sheep | [7,16,17,18] | |
| 22 | A10 | gi|17989230 | 19 kDa periplasmic protein | 20,238 | 68 | 6.06 | 8 | 1 | Sheep | M05 |
| 23 | A11 | gi|222447132 | Ferritin (bacterioferritin) | 20,895 | 68 | 6.05 | 33 | 5 | Sheep | |
| A13 | gi|222447132 | Ferritin (bacterioferritin) | 20,895 | 183 | 5.05 | 36 | 4 | Sheep | M12 | |
| 24 | A12 | gi|493690773 | Bacterioferritin, partial | 16,118 | 220 | 4.81 | 33 | 3 | Sheep | [16,17] |
| 25 | A21 | gi|89258175 | 31 kDa cell surface protein | 31,084 | 293 | 5.50 | 38 | 9 | Cow | [16] M21 |
| No. | Spot ID | Acc.ID | Protein | Mw | MOWSE Score a | pI | Sequence Coverage (%) | No. of Peptides Matching | Host | Reference/Cross-Matching Spot ID |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | M12 | gi|222447132 | Ferritin (Bacterioferritin) | 20,895 | 183 | 5.05 | 36 | 4 | Sheep, Goat, Cow, Buffalo | A13 |
| 2 | M19 | gi|225852817 | Sulfate ABC transporter substrate-binding protein | 37,151 | 324 | 5.51 | 44 | 12 | Sheep, Goat, Cow, Buffalo | |
| 3 | M20 | gi|17986956 | Thiosulfate-binding protein precursor | 37,152 | 34 | 5.31 | 5 | 1 | Sheep, Goat, Cow, Buffalo | |
| 4 | M24 | gi|17988780 | d-ribose-binding periplasmic protein precursor | 31,030 | 280 | 5.60 | 29 | 5 | Sheep, Goat, Cow, Buffalo | [12,13,14] A40 |
| 5 | M25 | gi|225851771 | Fumarylacetoacetate hydrolase domain-containing protein 2 | 30,118 | 471 | 5.00 | 61 | 11 | Sheep, Goat, Cow, Buffalo | A47 |
| M26 | gi|225851771 | Fumarylacetoacetate hydrolase domain-containing protein 2 | 30,118 | 492 | 5.00 | 56 | 11 | Sheep, Goat, Cow, Buffalo | ||
| 6 | M27 | gi|384211119 | Lysine-arginine-ornithine-binding periplasmic protein | 36,684 | 240 | 5.09 | 31 | 10 | Sheep, Goat, Cow, Buffalo | A45 |
| 7 | M36 | gi|516360216 | Sugar ABC transporter substrate-binding protein, partial | 44,963 | 121 | 5.15 | 50 | 15 | Sheep, Goat, Cow, Buffalo | A41 |
| 8 | M37 | gi|493172683 | Amino acid ABC transporter substrate-binding protein | 31,331 | 178 | 5.24 | 48 | 7 | Sheep, Goat, Cow, Buffalo | [16] |
| 9 | M40 | gi|384410242 | Amidohydrolase 3 | 63,567 | 265 | 5.47 | 42 | 20 | Sheep, Goat, Cow, Buffalo | |
| 10 | M22 | gi|493003797 | Hypothetical protein similar to amino acid ABC transporter substrate-binding protein | 21,946 | 90 | 5.06 | 33 | 4 | Sheep, Goat, Cow, Buffalo | |
| M38 | gi|493155701 | Hypothetical protein similar to ABC transporter substrate-binding protein | 58,947 | 437 | 4.97 | 43 | 24 | Sheep, Goat, Cow, Buffalo | ||
| M32 | gi|492818336 | Hypothetical protein similar to ABC transporter substrate-binding protein | 31,905 | 113 | 5.57 | 33 | 7 | Sheep, Goat | ||
| 11 | M14 | gi|490823297 | Alcohol dehydrogenase | 36,537 | 116 | 6.07 | 21 | 7 | Sheep, Goat, Cow | |
| M16 | gi|489059662 | Alcohol dehydrogenase | 43,149 | 99 | 7.66 | 25 | 9 | Sheep, Goat, Cow | ||
| 12 | M15 | gi|493009422 | Thiamine-binding periplasmic protein | 36,829 | 164 | 5.71 | 43 | 9 | Sheep, Goat, Cow | |
| 13 | M21 | gi|89258175 | 31 kDa cell surface protein | 31,084 | 96 | 5.50 | 52 | 10 | Sheep, Goat | [16] A21 |
| M30 | gi|89258175 | 31 kDa cell surface protein | 31,084 | 166 | 5.50 | 16 | 5 | Sheep, Goat | [16] | |
| 14 | M23 | gi|225686619 | rhizopine-binding protein | 33,294 | 257 | 5.11 | 55 | 11 | Sheep, Goat | |
| 15 | M01 | gi|384446825 | Superoxide dismutase, copper/zinc binding protein | 17,255 | 222 | 6.10 | 51 | 6 | Sheep | [7,16,17,18] A07 |
| M03 | gi|384446825 | Superoxide dismutase, copper/zinc binding protein | 17,255 | 86 | 6.1 | 57 | 6 | Sheep | [7,16,17,18] | |
| 16 | M02 | gi|118137288 | Copper/zinc superoxide dismutase | 16,176 | 297 | 6.11 | 63 | 7 | Sheep | |
| M04 | gi|551701922 | Copper/zinc superoxide dismutase | 16,176 | 83 | 6.11 | 61 | 6 | Sheep | ||
| 17 | M05 | gi|384446516 | 19 kDa periplasmic protein | 18,735 | 219 | 5.65 | 20 | 4 | Sheep | A10 |
| 18 | M07 | gi|495782928 | Transaldolase | 23,554 | 264 | 5.47 | 41 | 8 | Sheep | [18] |
| 19 | M08 | gi|493009465 | Fructose-6-phosphate aldolase | 23,554 | 244 | 5.47 | 22 | 5 | Sheep | |
| 20 | M43 | gi|490830157 | Hydrolase | 27,731 | 371 | 6.07 | 48 | 8 | Sheep | A22 |
| No. | Acc.ID | Protein | B. abortus | B. melitensis | ||
|---|---|---|---|---|---|---|
| Spot ID | Host | Spot ID | Host | |||
| 1 | gi|493015116 | Fumarylacetoacetate hydrolase family protein | A47 | Cow, Buffalo, Sheep, Goat | M25 | Cow, Buffalo, Sheep, Goat |
| 2 | gi|490830157 | Hydrolase | A22 | Cow, Buffalo, Sheep | M43 | Sheep |
| 3 | gi|493691811 | Sugar ABC transporter substrate-binding protein | A41 | Cow, Buffalo, Sheep | M36 | Cow, Buffalo, Sheep, Goat |
| 4 | gi|384211119 | Lysine-arginine-ornithine-binding periplasmic protein | A45 | Cow, Buffalo | M27 | Cow, Buffalo, Sheep, Goat |
| 5 | gi|17988780 | d-ribose-binding periplasmic protein precursor | A40 | Cow, Sheep | M24 | Cow, Buffalo, Sheep, Goat |
| 6 | gi|384446825 | Superoxide dismutase, copper/zinc binding protein | A07 | Sheep | M01 | Sheep |
| 7 | gi|384446516 | 19 kDa periplasmic protein | A10 | Sheep | M05 | Sheep |
| 8 | gi|222447132 | Ferritin (Bacterioferritin) | A13 | Sheep | M12 | Cow, Buffalo, Sheep, Goat |
| 9 | gi|89258175 | 31 kDa cell surface protein | A21 | Cow | M21 | Sheep, Goat |
| No. | Spot ID | Acc.ID | Protein | Locus, Query Cover (QC) and Identity (I) | Host | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 100% Identity | Brucella spp. | B. suis | B. ovis | Ochrobactrum spp. | Y. enterocolitica | Y. pseudotuberculosis | S. enterica | E. coli O:157 | |||||
| 1 | A47/M25 | gi|493015116 MW 29383 | FAHD2 | WP_006093223 B. Abortus QC 100% I 100% | WP_006162877 QC 80% I 96% | WP_006200925 QC 100% I 96% | YP_001258270 FAHD QC 100% I 96% | WP_006470802 QC 100% I 92% | Not found 16.04.2014 | YP_001401380 FAHD QC 98% I 62% | YP_001588666 QC 78% I 41% | Not found 16.04.2014 | Cow, Buffalo, Sheep, Goat |
| 2 | A01 | gi|256369084 MW 31892 | Dihydrodi-picolinate synthase | YP_003106592 B. microti CCM 4915 | WP_006165259 QC 100% I 99% | NP_697660 QC 100% I 99% | YP_001257393 QC 98% I 29% | WP_021587874 QC 100% I 95% | YP_006003506 QC 99% I 46% | YP_071290 QC 99% I 46% | WP_023259918 QC 99% I 45% | NP_311367 QC 99% I 45% | Cow, Buffalo, Sheep, Goat |
| 3 | A02 | gi|496823699 MW 36385 | Glyceraldehyde-3-phosphate dehydrogenase | WP_009374365 Brucella spp. QC 100% I 100% | WP_009374365 QC 100% I 100% | NP_698712 QC 100% I 99% | NP_698712.1 QC 100% I 99% | WP_021588015 QC 100% I 96% | WP_019083593Q C 98% I 54% | YP_071698 QC 99% I 46% | WP_000218344 QC 99% I 46% | ELW37260 QC 97% I 52% | Cow, Buffalo, Sheep, Goat |
| 4 | A26 | gi|226887955 MW 34152 | Lactate malate dehydrogenase | WP_002970355 B. abortus QC 98% I 100% | 3GVH_A B. melitensis QC 100% I 100% | YP_001628354 QC 98% I 99% | YP_001259751 QC 98% I 99% | WP_007872232 L/M dehydrogenase QC 98% I 98% | WP_019080697 QC 67% I 33% | YP_069003 QC 67% I 33% | YP_218284.1 QC 86% I 31% | ELV66131 QC 86% I 31% | Cow, Buffalo, Sheep, Goat |
| 5 | A49 | gi|17987134 MW 45462 | Phosphopyruvate hydratase | NP_539768 B. melitensis QC 100% I 100% | YP_008839865 QC 100% I 99% enolase | NP_698137 QC 100% I 99% | YP_001259054 QC 100% I 99% | YP_001370601 QC 100% I 97% | YP_001005091. QC 99% I 60% | YP_069296.1 QC 99% I 60% | WP_016735109 QC 99% I 61% | ELV67289 QC 99% I 61% | Cow, Buffalo, Sheep, Goat |
| 6 | M20 | gi|17986956 MW 37152 | Thiosulfate-binding protein precursor | NP_539590.1 B. melitensis QC 100% I 100% | WP_008934207 QC 100% I 99% | WP_020628554 QC 100% I 100% | YP_001259236 QC 100% I 99% | WP_021586689 QC 100% I 91% | AHM75213.1 QC 98% I 55% | YP_071244.1 QC 99% I 55% | WP_000290287 QC 93% I 57% | NP_288986 QC 92% I 57% | Sheep, Goat, Cow, Buffalo |
| 7 | M37 | gi|493172683 MW 31331 | Amino acid ABC transporter substrate-binding protein | WP_004685846 B. melitensis QC 100% I 94% | WP_006161567 Brucella spp QC 100% I 95% | NP_698767 putative branch QC 100% I 95% | NP_698767 putative branch QC 100% I 95% | WP_006467797 QC 100% I 90% | WP_019080170 QC 95% I 40% | Not found 16.04.2014 | WP_000822979 leucine branch QC 95% I 40% | ELV65532 leucine specific QC 95% I 42% | Sheep, Goat, Cow, Buffalo |
| 8 | M40 | gi|384410242 MW 63567 | Amidohydrolase 3 | YP_005602224 B. melitensis M5 QC 100% I 100% | YP_005114197 B. abortus QC 100% I 99% | WP_004689025 QC 100% I 99% | YP_001257534 amidohydrolase QC 88% I 31% | YP_001371888 QC 100% I 53% | Not found 16.04.2014 | Not found 16.04.2014 | WP_023220860 amidohydrolase QC 90% I 26% | Not found 16.04.2014 | Sheep, Goat, Cow, Buffalo |
| 9 | M22 | gi|493003797 MW 21946 | Hypothetical protein (amino acid ABC transporter substrate-binding protein) | WP_023080384 B. melitensis QC 100% I 100% | WP_006085596 B. abortus QC 100% I 100% | WP_023080435 QC 100% I 84% | YP_001258837 ABC transporter QC 100% I 100% | WP_006466755 ABC transporter QC 99% I 94% | YP_001006291. ABC transporter QC 98% I 39% | Not found 16.04.2014 | Not found 16.04.2014 | Not found 16.04.2014 | sheep, goat, cow, buffalo |
| 10 | M38 | gi|493155701 MW 58947 | Hypothetical protein (ABC transporter substrate-binding protein) | WP_006256535 B. melitensis QC 100% I 99% | WP_006164780 QC 100% I 100% | WP_006197818 QC 100% I 99% | WP_006157758 QC 99% I 70% | WP_010658797 ABC transporter QC 100% I 89% | WP_019083182 ABC transporter QC 97% I 40% | Not found 16.04.2014 | WP_023210061 ABC transporter QC 93% I 40% | Not found 16.04.2014 | sheep, goat, cow, buffalo |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wareth, G.; Eravci, M.; Weise, C.; Roesler, U.; Melzer, F.; Sprague, L.D.; Neubauer, H.; Murugaiyan, J. Comprehensive Identification of Immunodominant Proteins of Brucella abortus and Brucella melitensis Using Antibodies in the Sera from Naturally Infected Hosts. Int. J. Mol. Sci. 2016, 17, 659. https://doi.org/10.3390/ijms17050659
Wareth G, Eravci M, Weise C, Roesler U, Melzer F, Sprague LD, Neubauer H, Murugaiyan J. Comprehensive Identification of Immunodominant Proteins of Brucella abortus and Brucella melitensis Using Antibodies in the Sera from Naturally Infected Hosts. International Journal of Molecular Sciences. 2016; 17(5):659. https://doi.org/10.3390/ijms17050659
Chicago/Turabian StyleWareth, Gamal, Murat Eravci, Christoph Weise, Uwe Roesler, Falk Melzer, Lisa D. Sprague, Heinrich Neubauer, and Jayaseelan Murugaiyan. 2016. "Comprehensive Identification of Immunodominant Proteins of Brucella abortus and Brucella melitensis Using Antibodies in the Sera from Naturally Infected Hosts" International Journal of Molecular Sciences 17, no. 5: 659. https://doi.org/10.3390/ijms17050659