
Programming Effects of Prenatal Glucocorticoid Exposure with a Postnatal High-Fat Diet in Diabetes Mellitus


 , ,
, ,
Abstract
:
1. Introduction
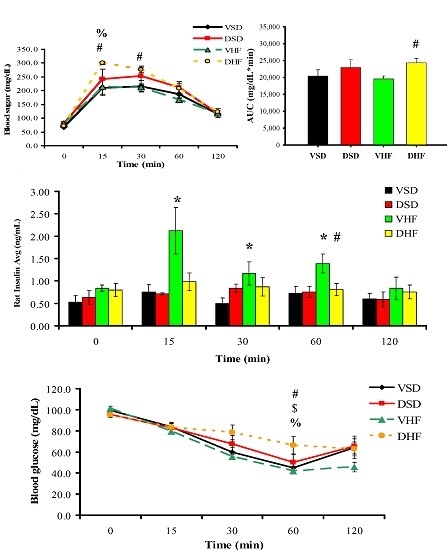
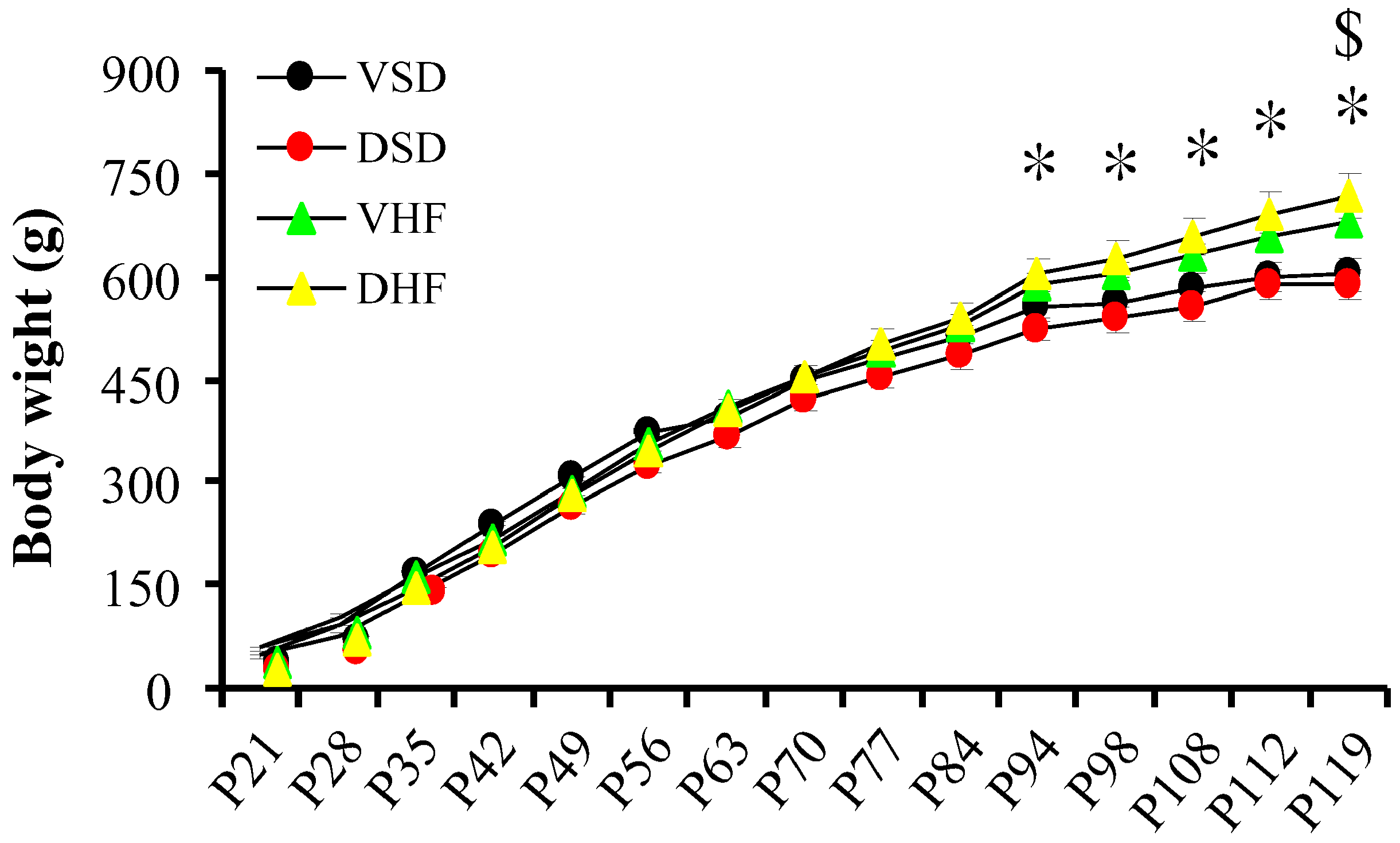
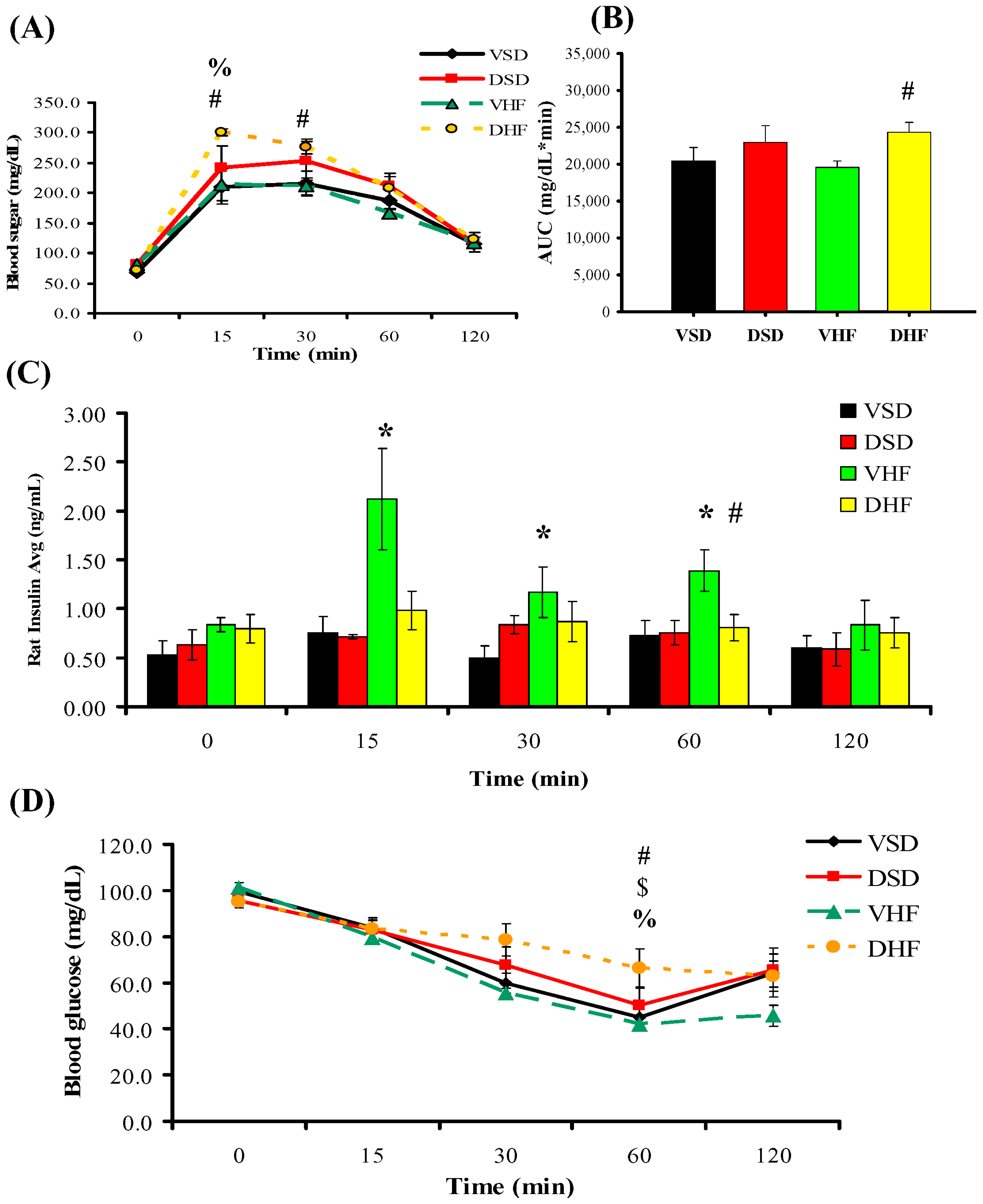
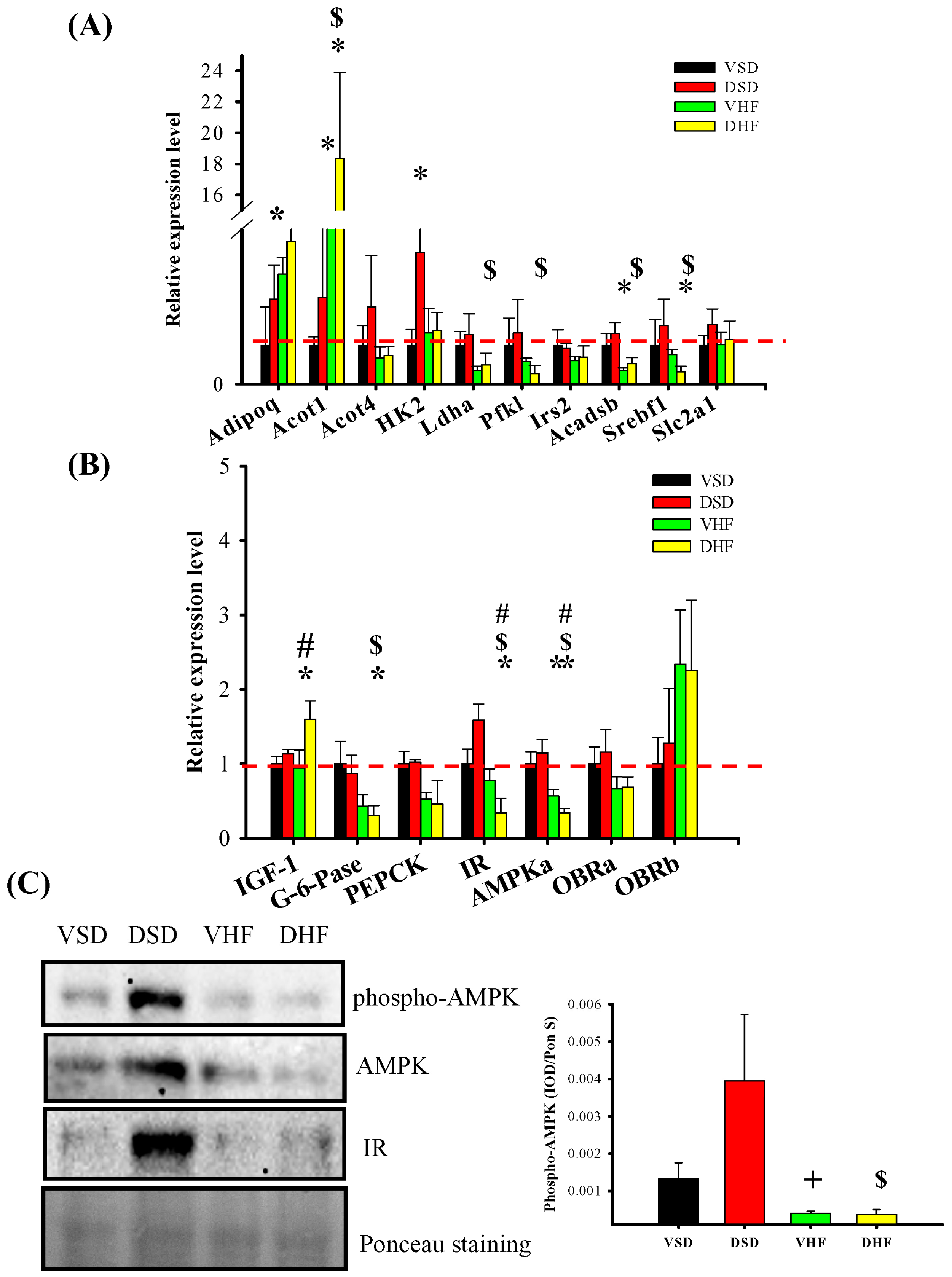
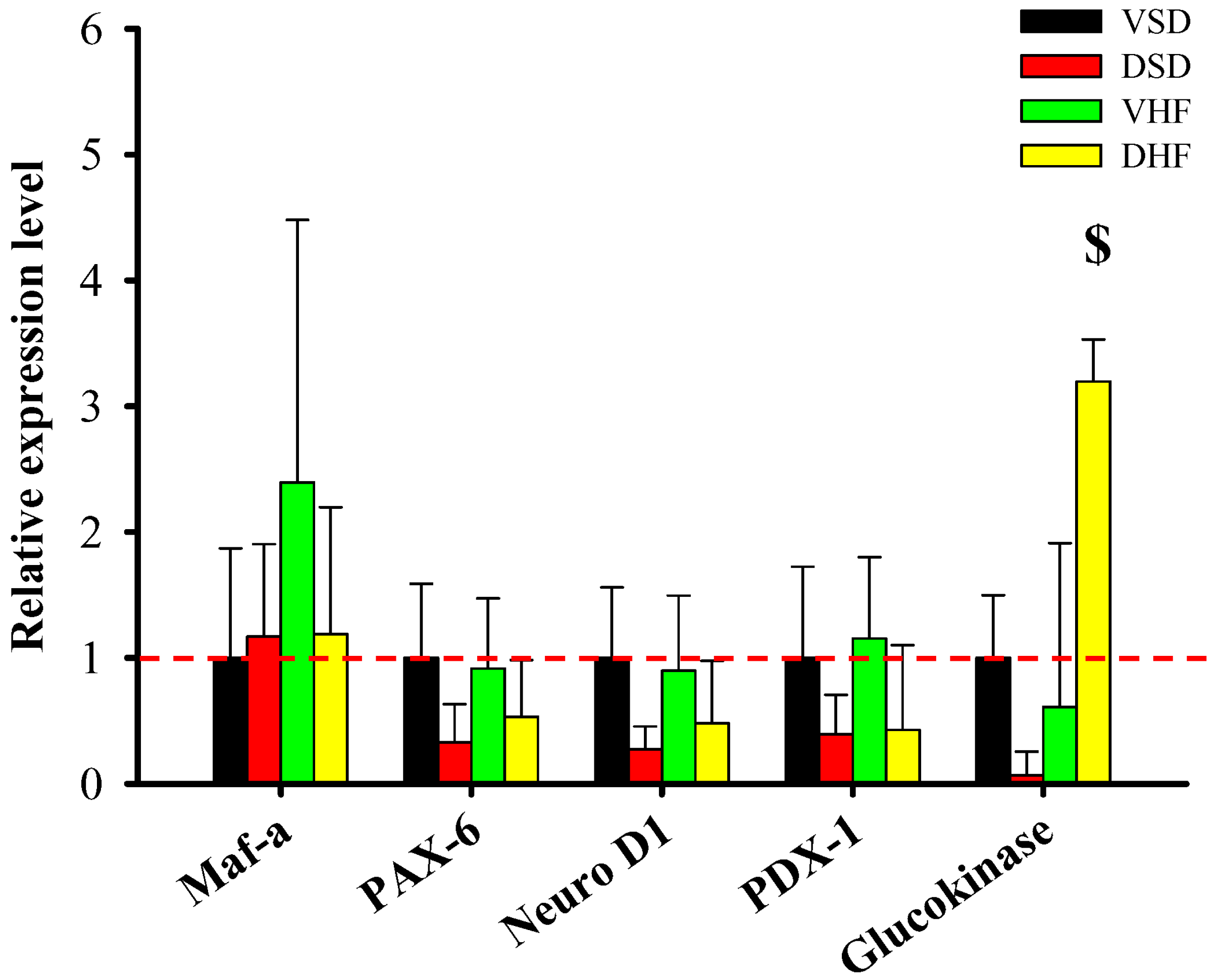
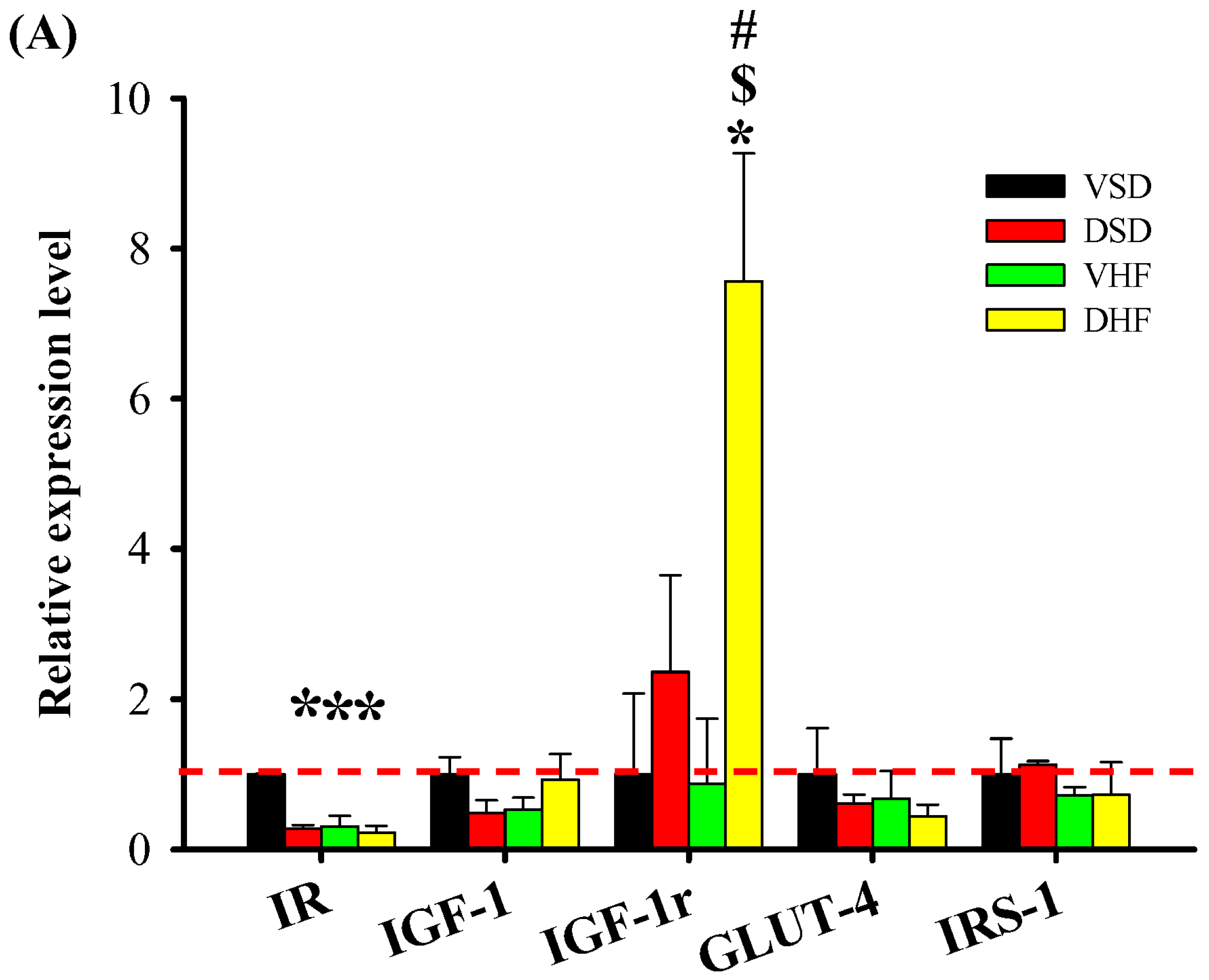
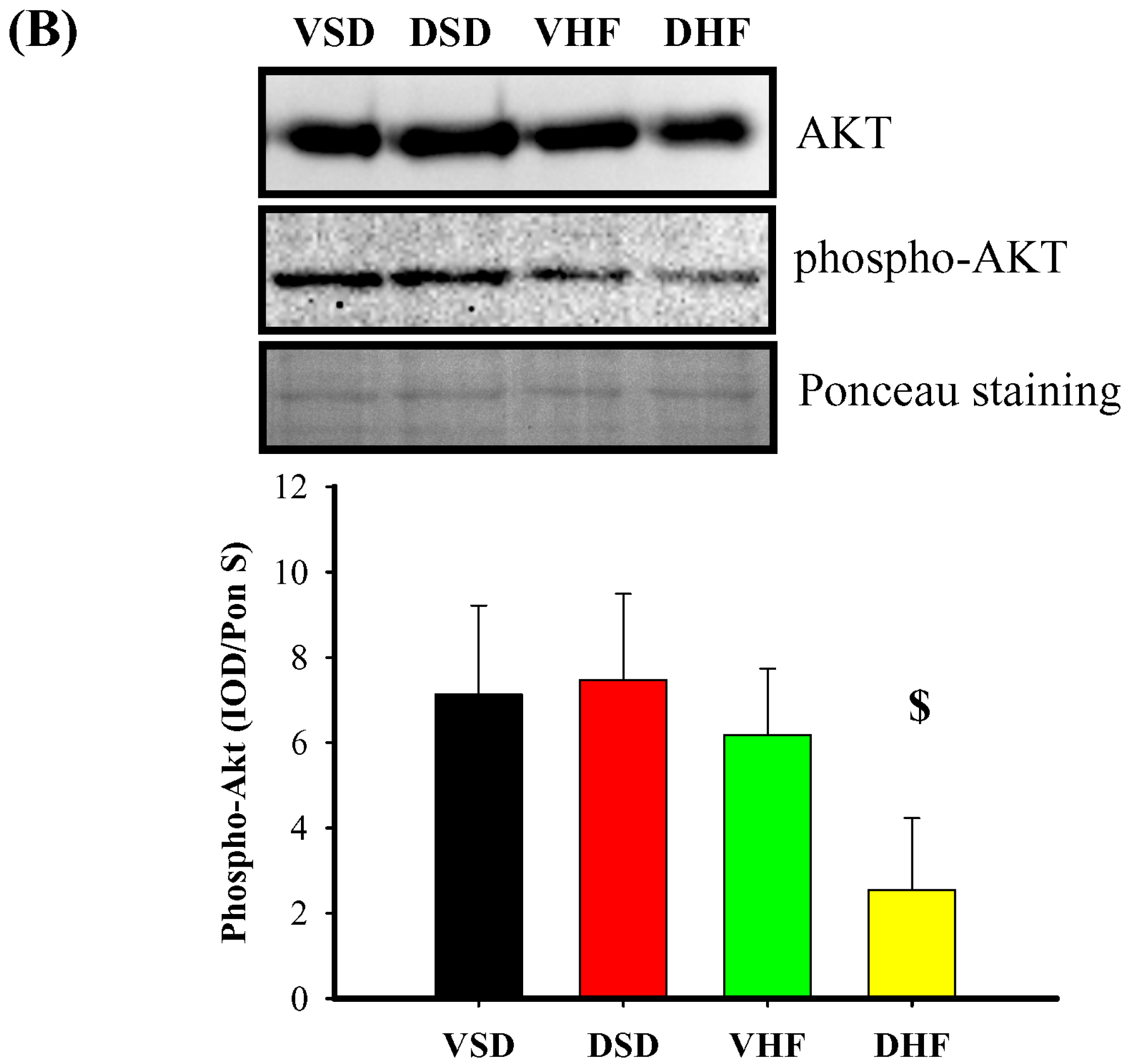
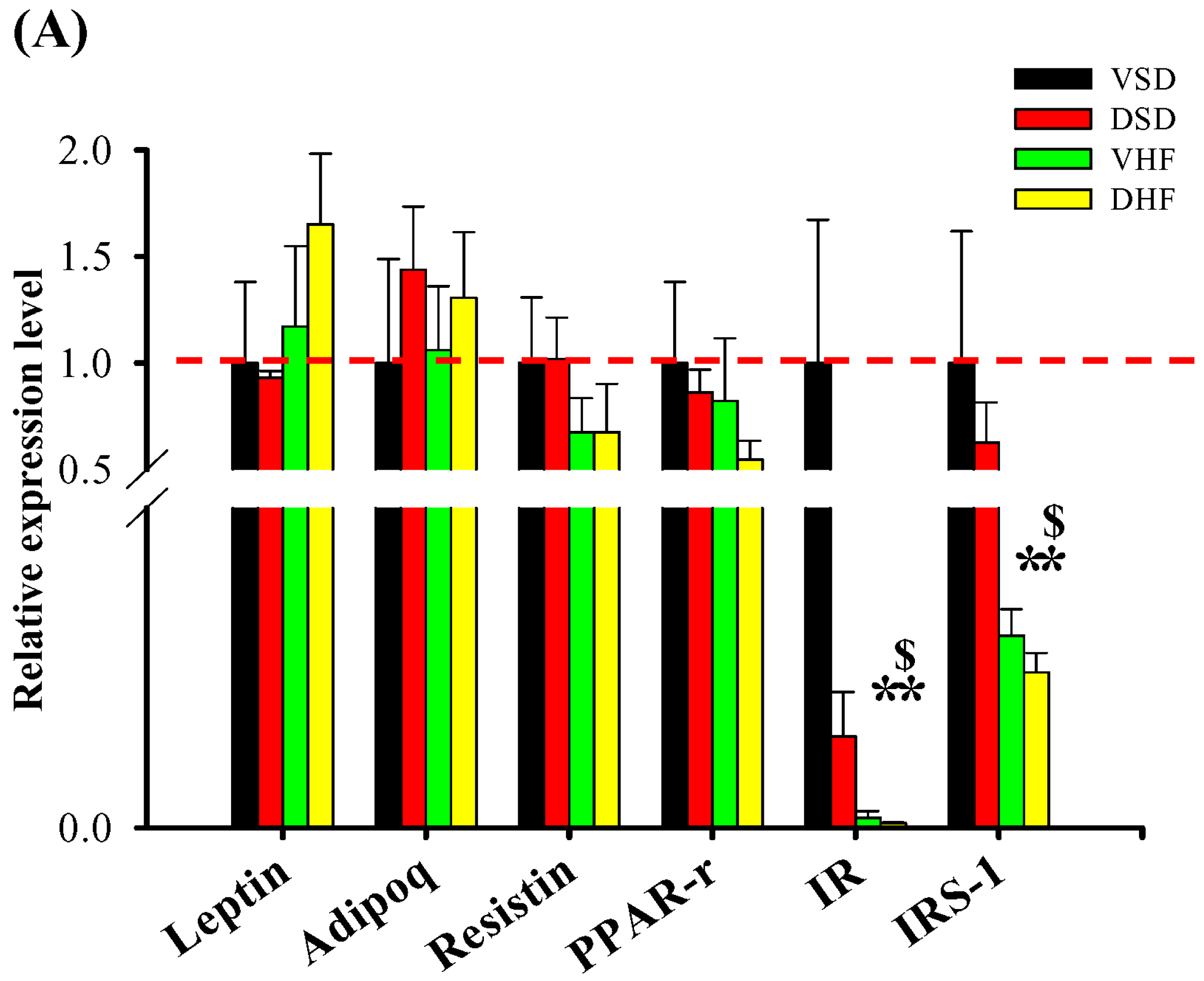
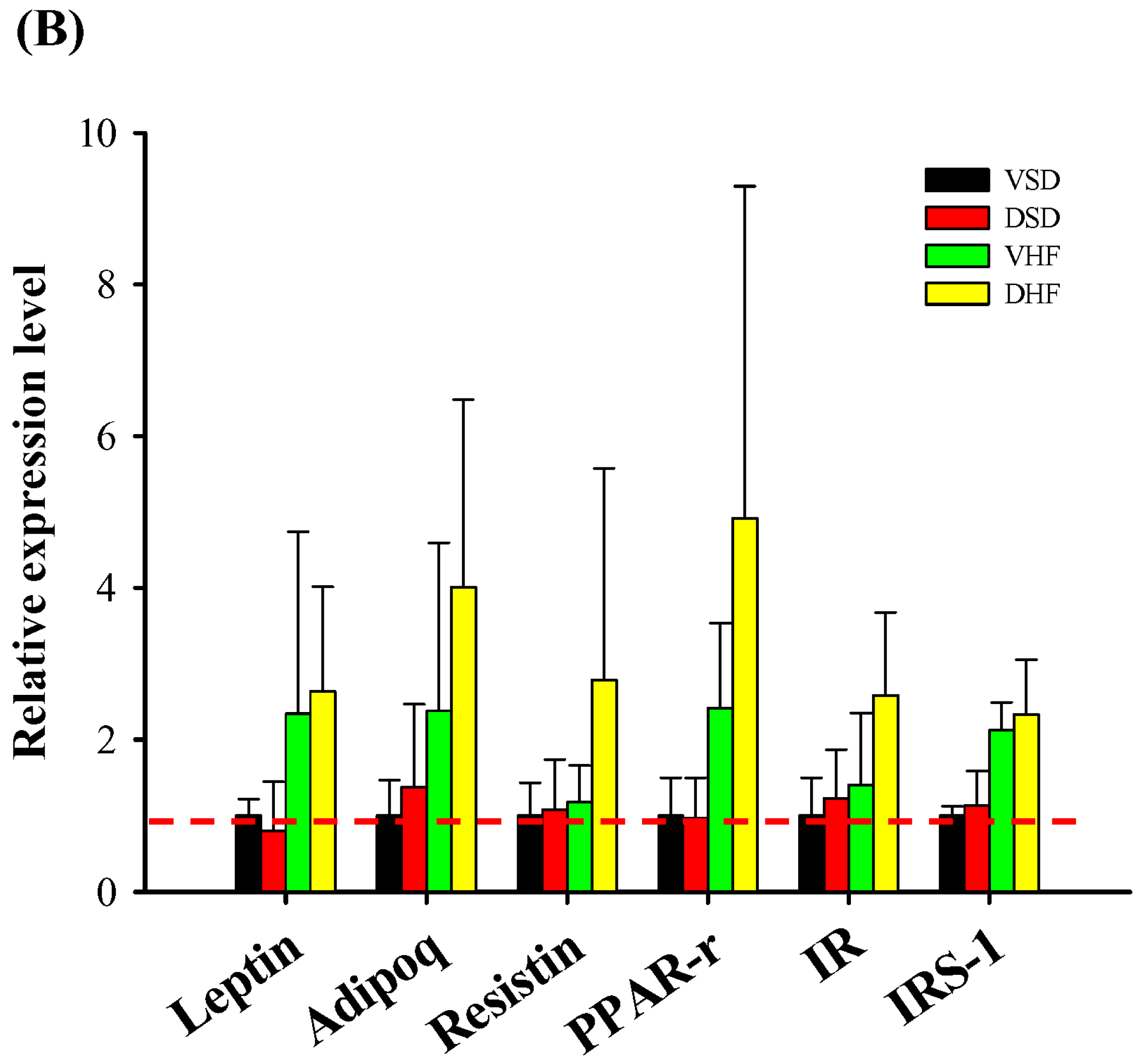
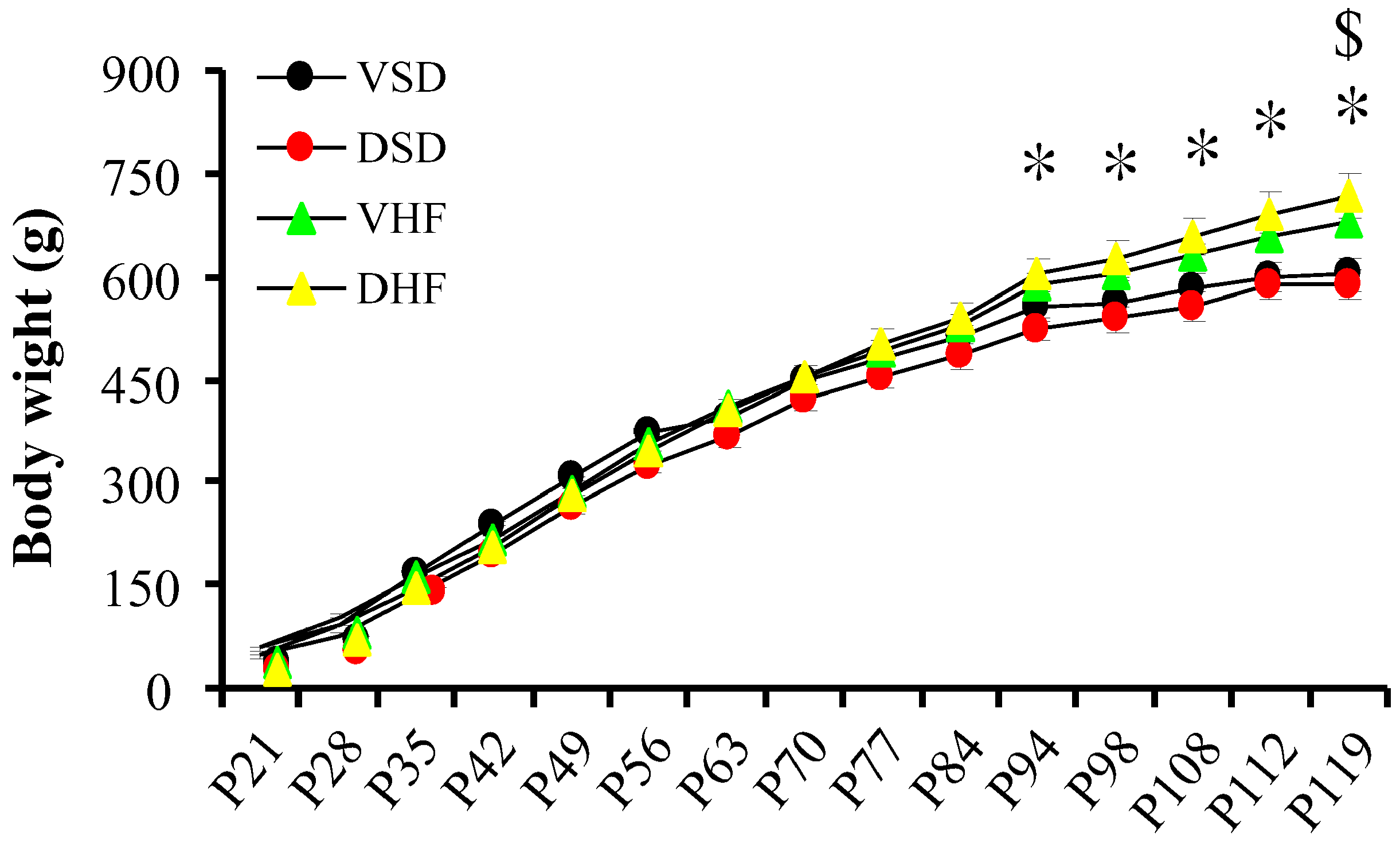
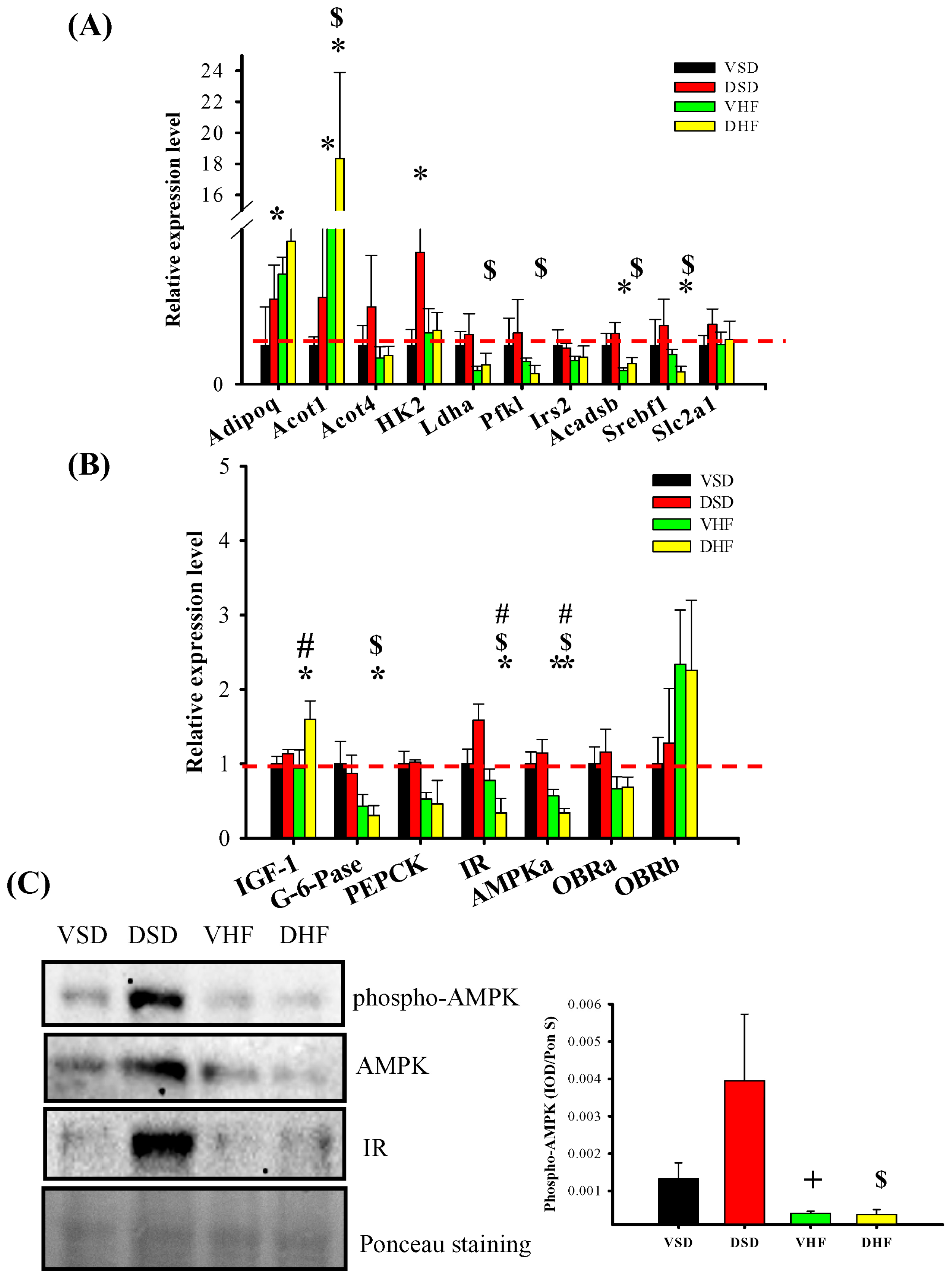
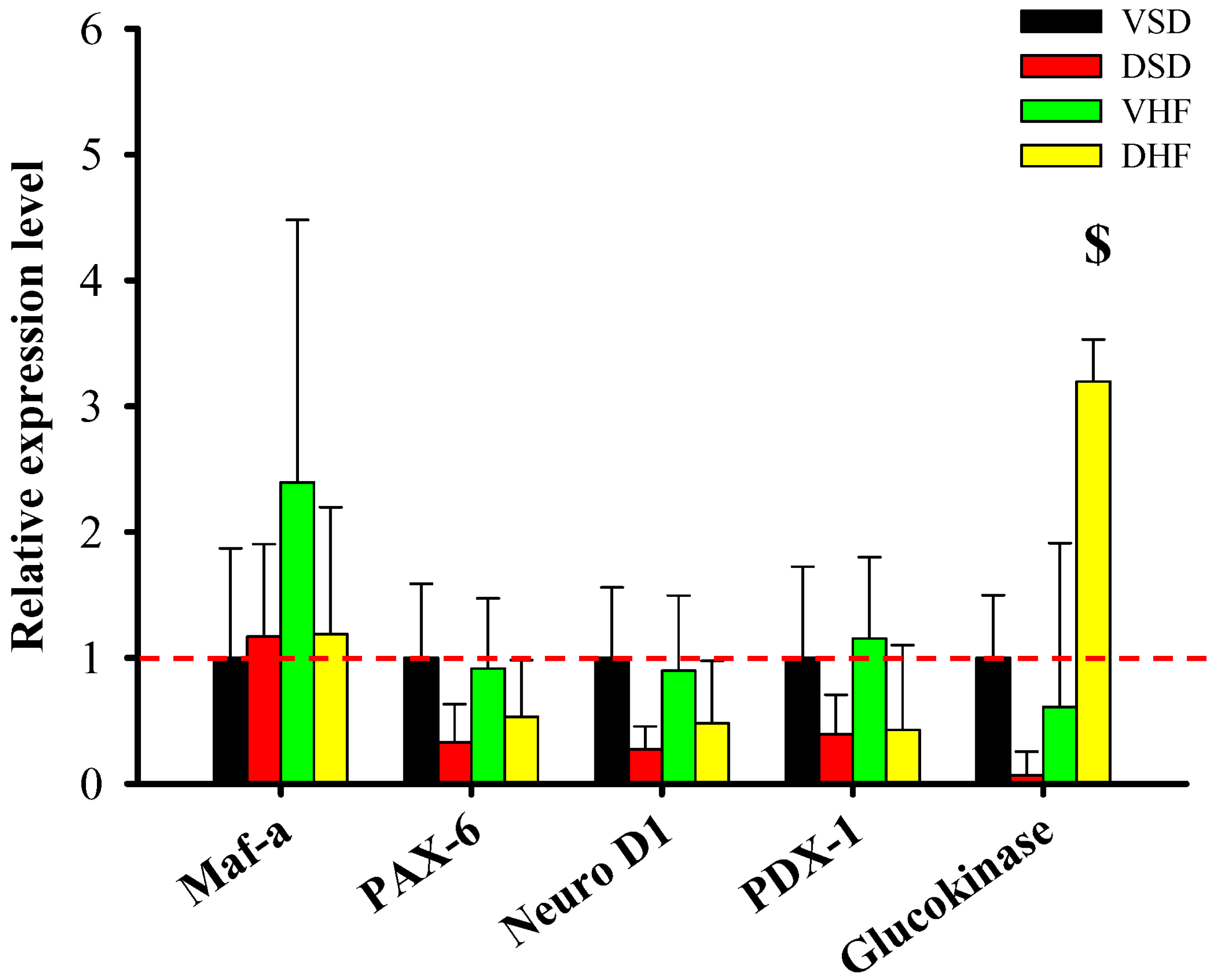
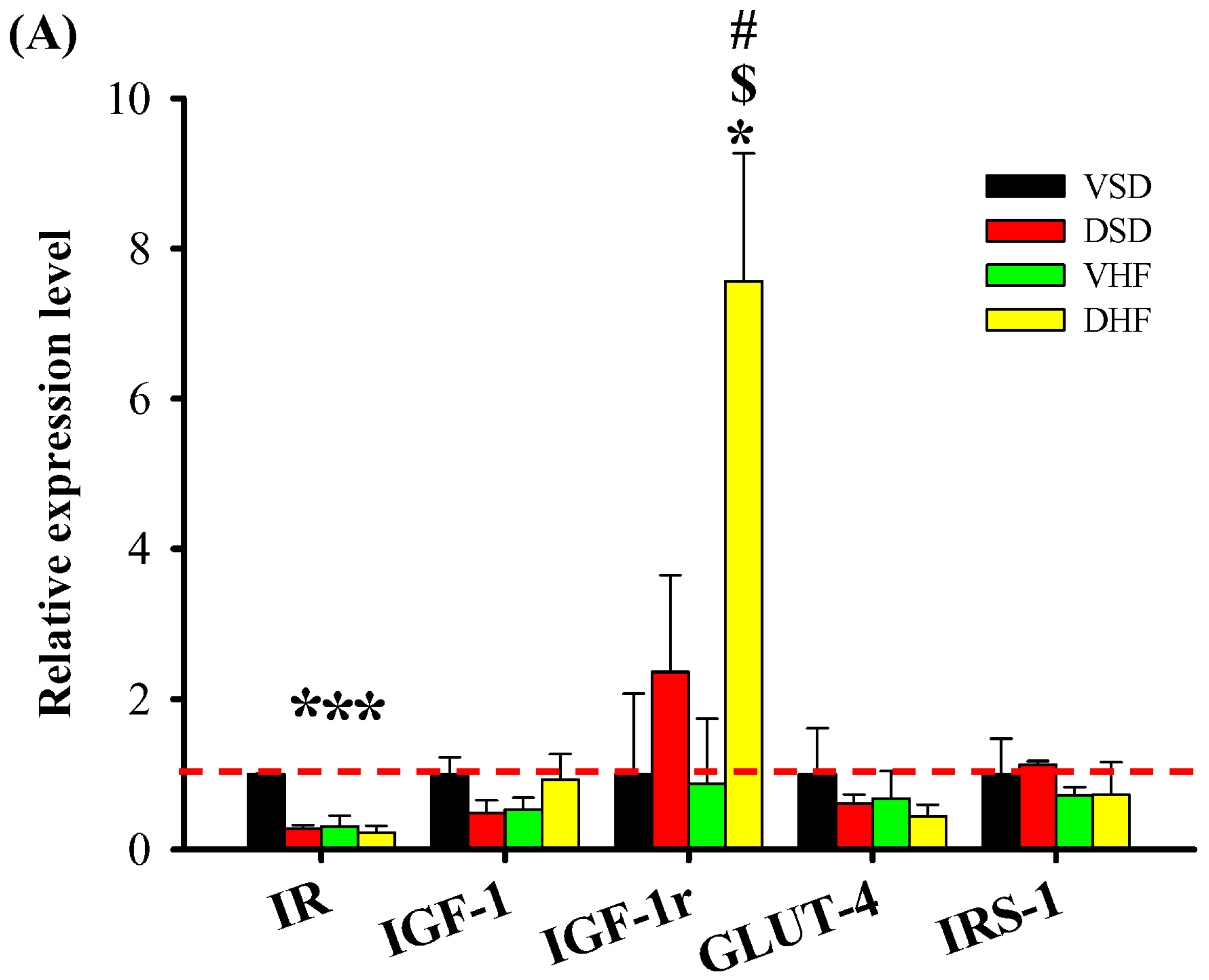
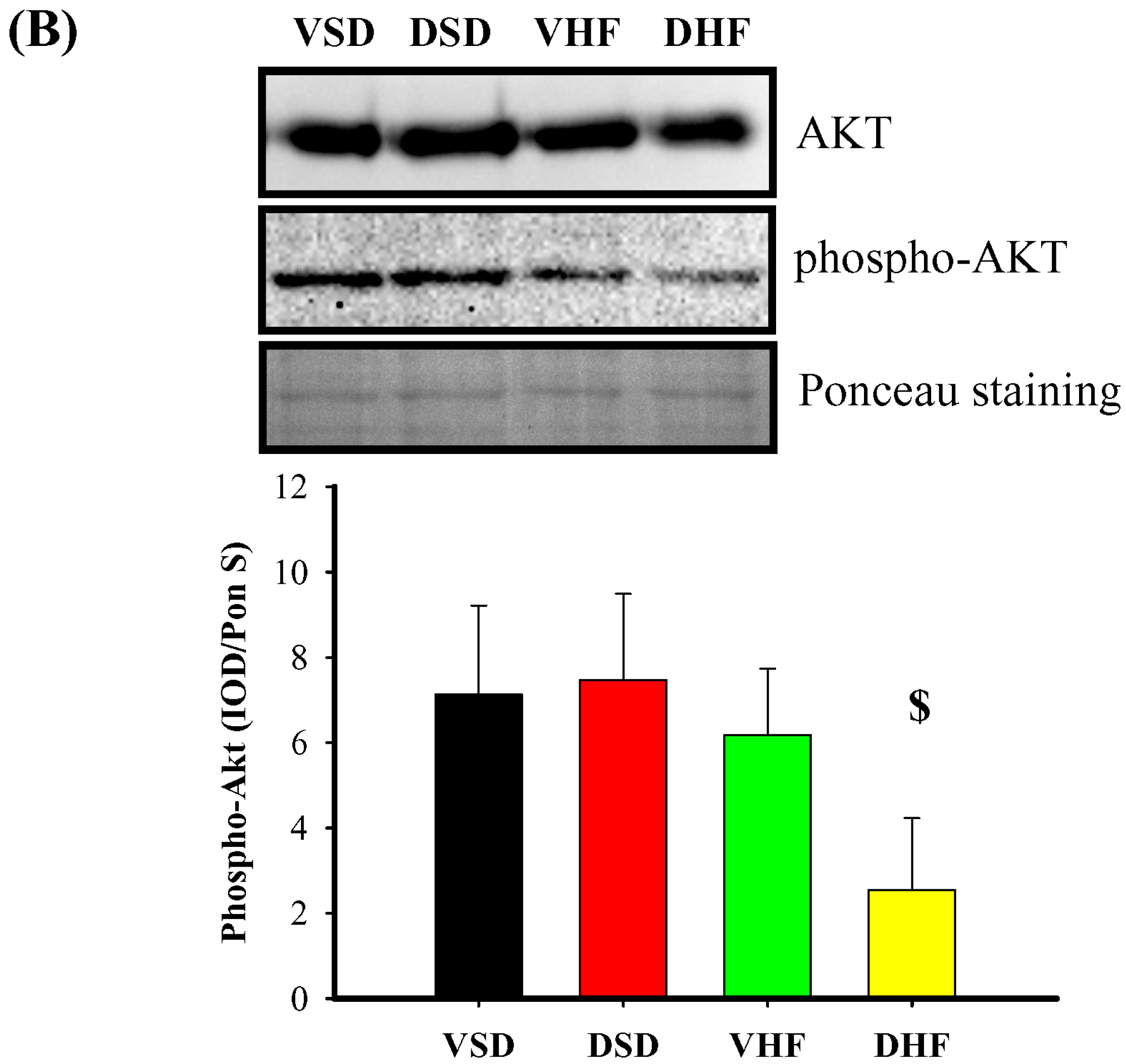
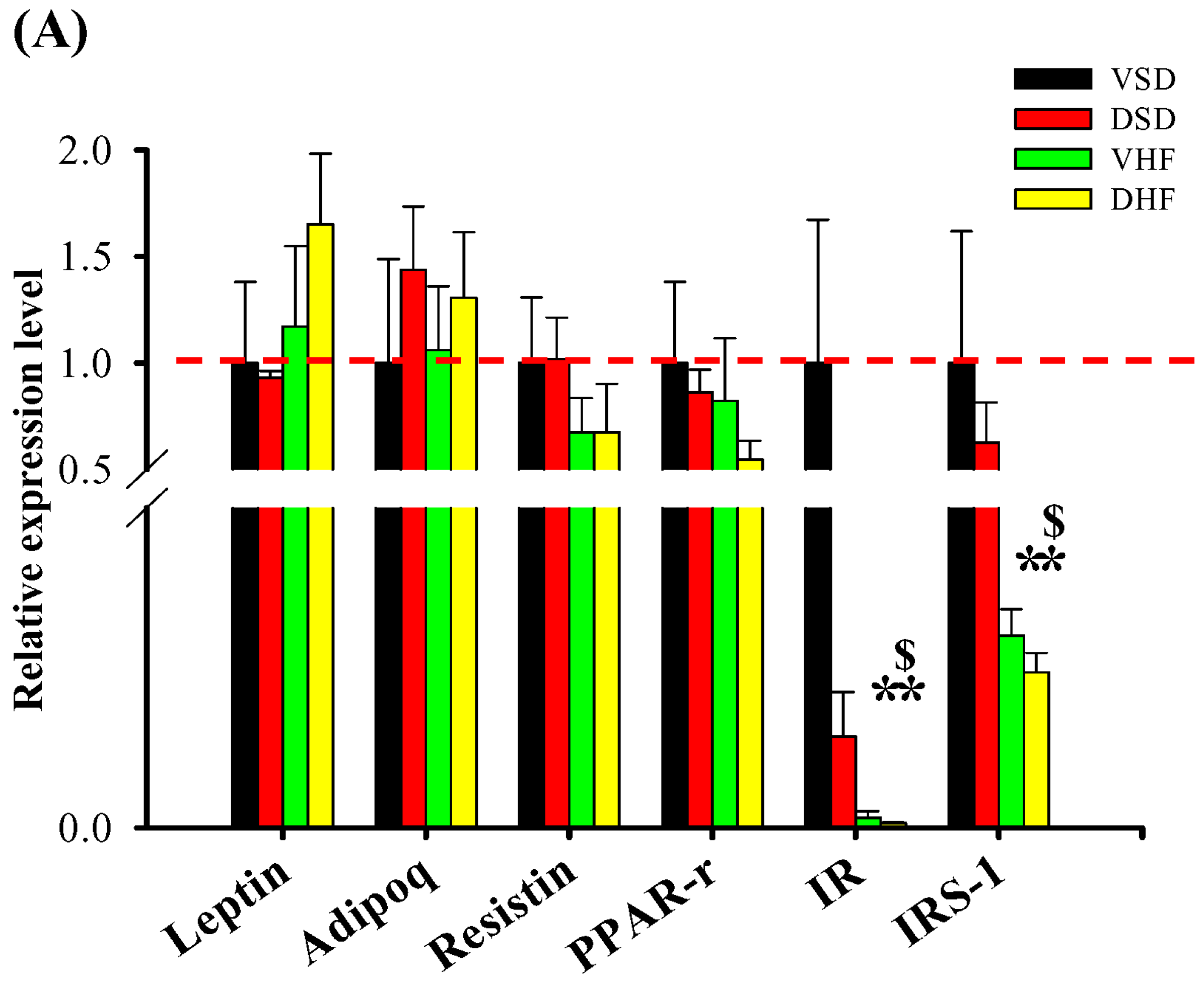
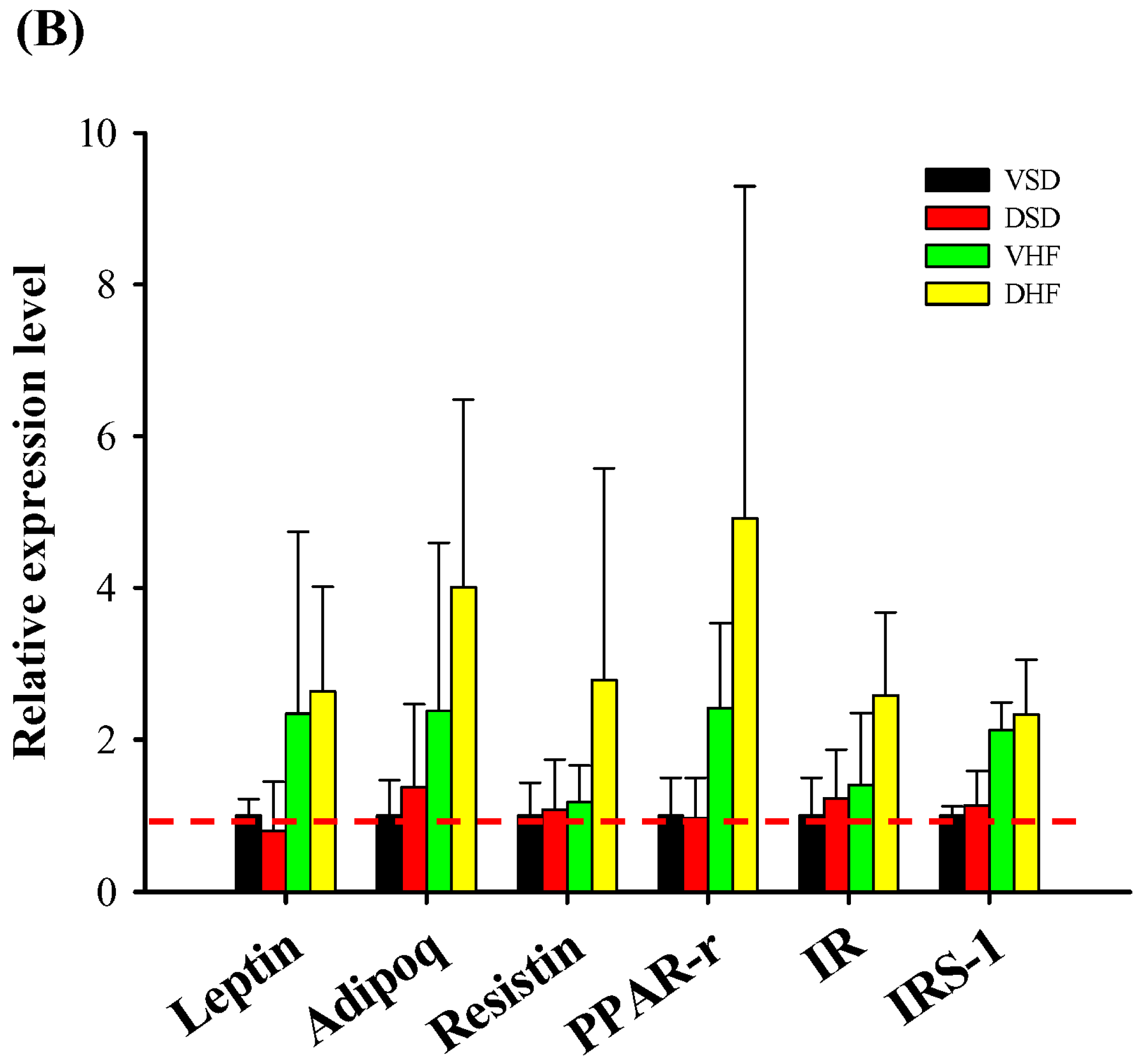
2. Results
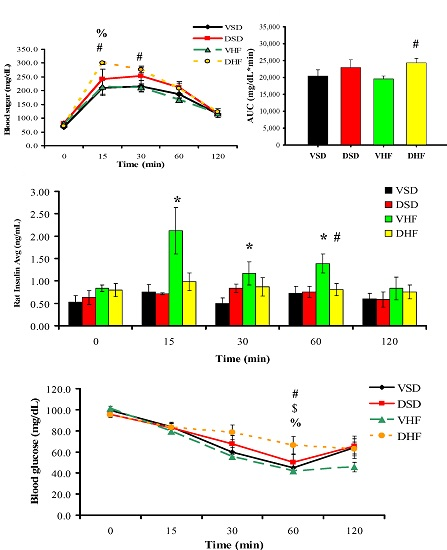
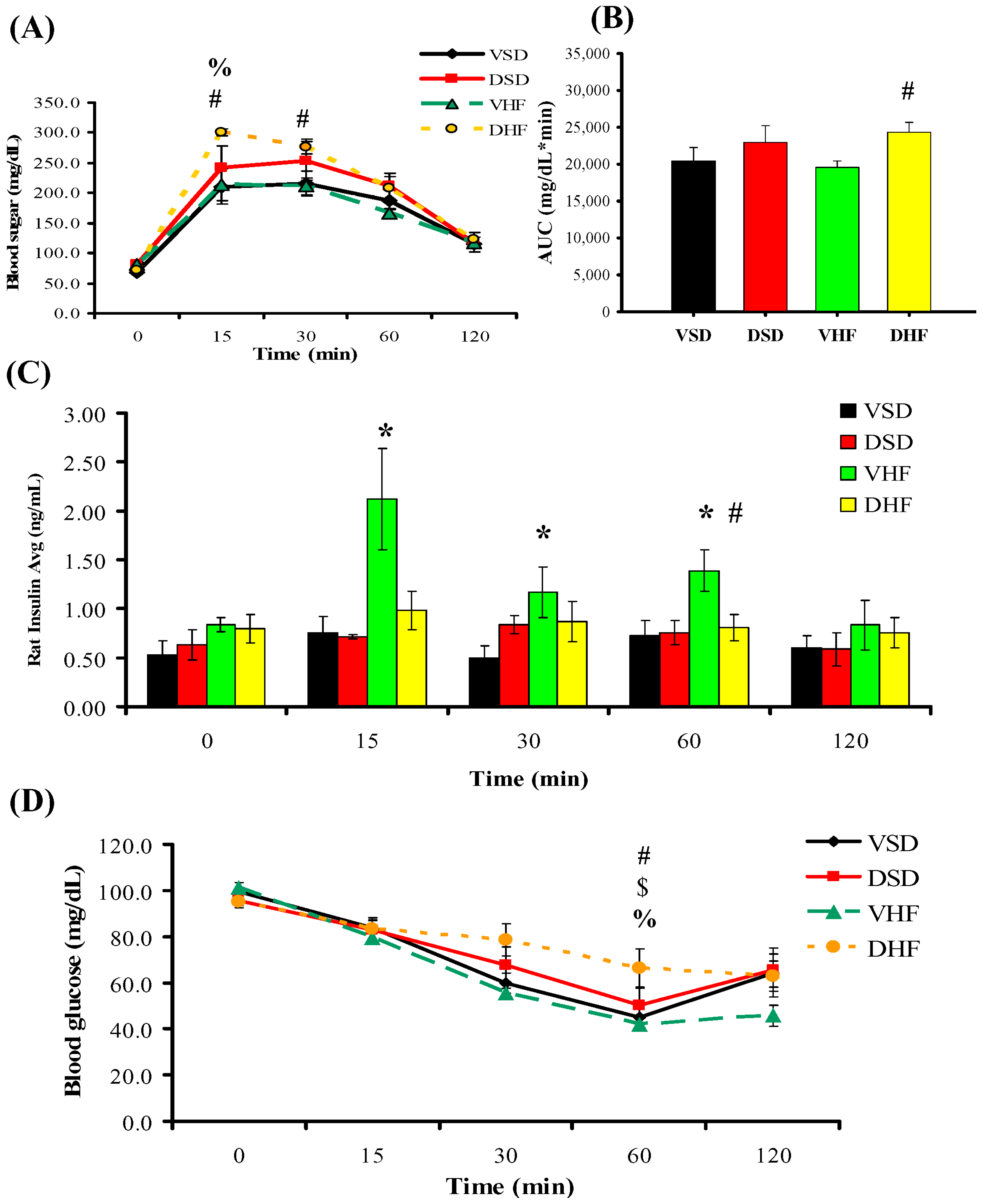
Intraperitoneal Glucose Tolerance Test (IPGTT) and Insulin Tolerance Test (ITT)
3. Discussion
4. Material and Methods
4.1. Animals
4.2. Prenatal Steroid Exposure and Postnatal High Fat Diet
- VSD group: Offspring of SD rats that received i.p. normal saline at GD 14–20; received normal diet after weaning, sacrificed at 4 months of age.
- DSD group: Offspring of SD rats that received i.p. dexamethasone at GD 14–20; received normal diet after weaning, sacrificed at 4 months of age.
- VHF group: Offspring of SD rats that received i.p. normal saline at GD 14–20; received high-fat diet (58% high-fat diet, Research Diet, D12330, soybean Oil 25 gm%, coconut Oil, hydrogenated 333.5 gm%) (Supplementary Table S1) from weaning, sacrificed at 4 months of age.
- DHF group: Offspring of SD rats that received i.p. dexamethasone at GD 14–20; received high-fat diet from weaning, sacrificed at 4 months of age.
4.3. I.P. Glucose Tolerance Test (IPGTT) and Insulin Tolerance Test (ITT)
4.4. Quantitative Real-Time Polymerase Chain Reaction (PCR) Analysis
4.5. Western Blot
4.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ozanne, S.E.; Constância, M. Mechanisms of disease: The developmental origins of disease and the role of the epigenotype. Nat. Clin. Pract. Endocrinol. Metab. 2007, 3, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.J. In utero programming of chronic disease. Clin. Sci. 1998, 95, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Hales, C.N.; Barker, D.J. Type 2 (non-insulin-dependent) diabetes mellitus: The thrifty phenotype hypothesis. Diabetologia 1992, 35, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Weinberger, B.; Anwar, M.; Henien, S.; Sosnovsky, A.; Hiatt, M.; Jochnowitz, N.; Witz, G.; Hegyi, T. Association of lipid peroxidation with antenatal betamethasone and oxygen radial disorders in preterm infants. Biol. Neonate 2004, 85, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Roberts, D.; Dalziel, S. Antenatal corticosteroids for accelerating fetal lung maturation for women at risk of preterm birth. Cochrane Database Syst. Rev. 2006, 3, CD004454. [Google Scholar] [CrossRef] [PubMed]
- Dalziel, S.R.; Walker, N.K.; Parag, V.; Mantell, C.; Rea, H.H.; Rodgers, A.; Harding, J.E. Cardiovascular risk factors after antenatal exposure to betamethasone: 30-year follow-up of a randomised controlled trial. Lancet 2005, 365, 1856–1862. [Google Scholar] [CrossRef]
- Pankevich, D.E.; Mueller, B.R.; Brockel, B.; Bale, T.L. Prenatal stress programming of offspring feeding behavior and energy balance begins early in pregnancy. Physiol. Behav. 2009, 98, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Van Uum, S.H.; Hermus, A.R.; Smits, P.; Thien, T.; Lenders, J.W. The role of 11b-hydroxysteroid dehydrogenase in the pathogenesis of hypertension. Cardiovasc. Res. 1998, 38, 16–24. [Google Scholar] [CrossRef]
- Reynolds, R.M.; Walker, B.R.; Syddall, H.E.; Andrew, R.; Wood, P.J.; Whorwood, C.B.; Phillips, D.I. Altered control of cortisol secretion in adult men with low birth weight and cardiovascular risk factors. J. Clin. Endocrinol. Metab. 2001, 86, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Gnanalingham, M.G.; Mostyn, A.; Symonds, M.E.; Stephenson, T. Ontogeny and nutritional programming of adiposity in sheep: Potential role of glucocorticoid action and uncoupling protein-2. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 289, R1407–R1415. [Google Scholar] [CrossRef] [PubMed]
- Tamashiro, K.L.; Terrillion, C.E.; Hyun, J.; Koenig, J.I.; Moran, T.H. Prenatal stress or high-fat diet increases susceptibility to diet-induced obesity in rat offspring. Diabetes 2009, 58, 1116–1125. [Google Scholar] [CrossRef] [PubMed]
- Storlien, L.H.; Pan, D.A.; Kriketos, A.D.; Baur, L.A. High fat diet-induced insulin resistance. Lessons and implications from animal studies. Ann. N. Y. Acad. Sci. 1993, 683, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Bray, G.A.; Lovejoy, J.C.; Smith, S.R.; DeLany, J.P.; Lefevre, M.; Hwang, D.; Ryan, D.H.; York, D.A. The influence of different fats and fatty acids on obesity, insulin resistance and inflammation. J. Nutr. 2002, 132, 2488–2491. [Google Scholar] [PubMed]
- Kuo, T.; McQueen, A.; Chen, T.C.; Wang, J.C. Regulation of glucose homeostasis by glucocorticoids. Adv. Exp. Med. Biol. 2015, 872, 99–126. [Google Scholar] [PubMed]
- Vegiopoulos, A.; Herzig, S. Glucocorticoids, metabolism and metabolic diseases. Mol. Cell. Endocrinol. 2007, 275, 43–61. [Google Scholar] [CrossRef] [PubMed]
- Franko, K.L.; Forhead, A.J.; Fowden, A.L. Differential effects of prenatal stress and glucocorticoid administration on postnatal growth and glucose metabolism in rats. J. Endocrinol. 2010, 204, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.C.; Huang, Y.H.; Sheen, J.M.; Tain, Y.L.; Yu, H.R.; Chen, C.C.; Tiao, M.M.; Kuo, H.C.; Huang, L.T. Prenatal dexamethasone exposure programs the development of the pancreas and the secretion of insulin in rats. Pediatr. Neonatol. 2016, in press. [Google Scholar]
- De Blasio, M.J.; Dodic, M.; Jefferies, A.J.; Moritz, K.M.; Wintour, E.M.; Owens, J.A. Maternal exposure to dexamethasone or cortisol in early pregnancy differentially alters insulin secretion and glucose homeostasis in adult male sheep offspring. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E75–E82. [Google Scholar] [CrossRef] [PubMed]
- Meier, J.J.; Bonadonna, R.C. Role of reduced β-cell mass versus impaired β-cell function in the pathogenesis of type 2 diabetes. Diabetes Care 2013, 36, S113–S119. [Google Scholar] [CrossRef] [PubMed]
- Rahier, J.; Guiot, Y.; Goebbels, R.M.; Sempoux, C.; Henquin, J.C. Pancreatic β-cell mass in European subjects with type 2 diabetes. Diabetes Obes. Metab. 2008, 10, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Matsushita, Y.; Nakagawa, T.; Hayashi, T.; Noda, M.; Mizoue, T. Circulating adiponectin levels and risk of type 2 diabetes in the Japanese. Nutr. Diabetes 2014, 4, e130. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Gomez, V.; Lu, L.; Yang, X.; Wu, X.; Xiao, S.Y. Expression of adiponectin and its receptors in livers of morbidly obese patients with non-alcoholic fatty liver disease. J. Gastroenterol. Hepatol. 2009, 24, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Li, J.B.; Wang, C.Y.; Chen, J.W.; Feng, Z.Q.; Ma, H.T. Expression of liver insulin-like growth factor 1 gene and its serum level in patients with diabetes. World J. Gastroenterol. 2004, 10, 255–259. [Google Scholar] [PubMed]
- Zisman, A.; Peroni, O.D.; Abel, E.D.; Michael, M.D.; Mauvais-Jarvis, F.; Lowell, B.B.; Wojtaszewski, J.F.; Hirshman, M.F.; Virkamaki, A.; Goodyear, L.J.; et al. Targeted disruption of the glucose transporter 4 selectively in muscle causes insulin resistance and glucose intolerance. Nat. Med. 2000, 6, 924–928. [Google Scholar] [PubMed]
- Viollet, B.; Foretz, M.; Guigas, B.; Horman, S.; Dentin, R.; Bertrand, L.; Hue, L.; Andreelli, F. Activation of AMP-activated protein kinase in the liver: A new strategy for the management of metabolic hepatic disorders. J. Physiol. 2006, 574, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Coughlan, K.A.; Valentine, R.J.; Ruderman, N.B.; Saha, A.K. AMPK activation: A therapeutic target for type 2 diabetes? Diabetes Metab. Syndr. Obes. 2014, 7, 241–253. [Google Scholar] [PubMed]
- Kahn, B.B.; Flier, J.S. Obesity and insulin resistance. J. Clin. Investig. 2000, 106, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Sheen, J.M.; Chen, C.C.; Yu, H.R.; Tiao, M.M.; Kuo, H.C.; Huang, L.T. Maternal citrulline supplementation prevents prenatal dexamethasone-induced programmed hypertension. Free Radic. Res. 2014, 48, 580–586. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sense (5′–3′) | Anti-Sense (3′–5′) |
|---|---|---|
| Adipocytokine signaling pathway | ||
| Adiponectin | GGAGACGCAGGTGTTCTTGG | AGCCCTACGCTGAATGCTGA |
| Leptin | CGGTTCCTGTGGCTTTGGT | CCGACTGCGTGTGTGAAATG |
| OBRa | CCTCTTGTGTCCTGCTGCTCGG | TTCTATGGACTGTTGGGAGGTTGGT |
| OBRb | GCATGCAGAATCAGTGATATTTGG | CAAGCTGTATCGACACTGATTTCTTC |
| Slc2a1 | GCTCCATTTAGGATTCGCCCA | TATACACAGCAGGGCAGGAGT |
| Resistin | TCATGCCCAGAACCGAGTTG | CAGCCCCAGGACAAGGAAGA |
| Visfatin | TCTGGAAATCCGCTCGACAC | CACTCCGTCCCCTTGAATGA |
| Fatty acid metabolism | ||
| Acadsb | GGACTGGCCCAAGGATGTTT | ATAAATGGCCTCCCGGCTTC |
| Acot1 | GTGATGGTTTTGGCAGGAAAAGT | AATGTGCTCTTTTCCCTTACAGC |
| Acot4 | TTGCCATCTCAATGGGGTAGAT | AGGGAGTCTCTCTTAACGTTTACC |
| Perilipin1 | GAGGGGCTGATCTGGCTTTG | GCATCTTTTGCCGTCCTGAA |
| PPAR-r | GGCTTCATGACAAGGGAGTTTC | AACTCAAACTTGGGCTCCATAAAG |
| Glycolysis/Gluconeogenesis | ||
| G6Pase | AACGTCTGTCTGTCCCGGATCTAC | ACCTCTGGAGGCTGGCATTG |
| Ldha | TCAGCGTCCCATGTATCCTG | CTGGACCAACTGGACTAACCA |
| PEPCK | CTCACCTCTGGCCAAGATTGGTA | GTTGCAGGCCCAGTTGTTGA |
| Pfkl | CTTACCGATCACCCTCGTTC | CCACAGGTGCTCTGTTCTGA |
| Insulin/IGF-1 signaling pathway | ||
| AMPKa | GTCGGCACCTTCGGCAAAGTGAA | AGAAATTCACCATCTGACATCATATTAGA |
| Glucokinase | CAGTGGAGCGTGAAGACAAA | AGGGAAGGAGAAGGTGGAGC |
| GLUT-4 | TTTCCAGTATGTTGCGGATG | TCAGTCATTCTCATCTGGCC |
| Hexokinase 2 | ATGGTCCTCCCCCACTCTAC | TCCCACCCAACATCTACCTC |
| IR | TCGAACCCTTCCTAACAG | CAGGTCCAAAGACAAACAGA |
| IRS-1 | ATCTTCCTTTGGCGCAGCTA | CAGCACGAAAAAGCGCTTA |
| IRS-2 | GAATCCCCCAGGGACAGTAG | GGGGAGGGGGAGTTTAGTGT |
| IGF-1 | TCAGTTCGTG TGTGGACCAG | TCACAGCTCCGGAAGCAAC |
| IGF-1r | TGGCAGAACTGCTGTCTGAG | AACGCAGGGTCTAGTTGAGC |
| SREBF1 | GCTGATGGAGACAGGGAGTT | GCAGTTGATGTAGAGGCTAAGC |
| Insulin secretion | ||
| Maf-a | GGAGGTCATCCGACT GAAACA | CCGCCAACTTCTCGTATTTCTC |
| Neuro D1 | CTCGCTGTGA GATCC CCATAG | TAATCGTGAAAGATGGCATTAAGC |
| PAX-6 | CTCCTCGTACTCCTGCATGCT | GGGCTGACTGTTCATGTGTGTT |
| Pdx-1 | GCTGGAGCTGGAGAAGGAAT | CGTTGTCCCGCTACGTT |
| β-Actin | TACTGCCCTGGCTCCTA | GGGCCGGACTCATCGTA |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sheen, J.-M.; Hsieh, C.-S.; Tain, Y.-L.; Li, S.-W.; Yu, H.-R.; Chen, C.-C.; Tiao, M.-M.; Chen, Y.-C.; Huang, L.-T. Programming Effects of Prenatal Glucocorticoid Exposure with a Postnatal High-Fat Diet in Diabetes Mellitus. Int. J. Mol. Sci. 2016, 17, 533. https://doi.org/10.3390/ijms17040533
Sheen J-M, Hsieh C-S, Tain Y-L, Li S-W, Yu H-R, Chen C-C, Tiao M-M, Chen Y-C, Huang L-T. Programming Effects of Prenatal Glucocorticoid Exposure with a Postnatal High-Fat Diet in Diabetes Mellitus. International Journal of Molecular Sciences. 2016; 17(4):533. https://doi.org/10.3390/ijms17040533
Chicago/Turabian StyleSheen, Jiunn-Ming, Chih-Sung Hsieh, You-Lin Tain, Shih-Wen Li, Hong-Ren Yu, Chih-Cheng Chen, Miao-Meng Tiao, Yu-Chieh Chen, and Li-Tung Huang. 2016. "Programming Effects of Prenatal Glucocorticoid Exposure with a Postnatal High-Fat Diet in Diabetes Mellitus" International Journal of Molecular Sciences 17, no. 4: 533. https://doi.org/10.3390/ijms17040533