Mucin 1 Gene (MUC1) and Gastric-Cancer Susceptibility

Abstract
:1. Introduction
2. Association between GC (Gastric Cancer) and MUC1
3. MUC1 Expression in Gastric Carcinogenesis
4. MUC1 Function in Normal Gastric Epithelial Cells
5. MUC1 Function in Gastric Carcinogenesis
6. Perspective and Conclusions
Acknowledgments
Conflicts of Interest
References
- Brenner, H.; Rothenbacher, D.; Arndt, V. Epidemiology of gastric cancer. In Methods of Molecular Biology, Cancer Epidemiology; Verma, M., Ed.; Humana Press: Totowa, NJ, USA, 2009; Volume 472, p. 467. [Google Scholar]
- Yasui, W.; Sentani, K.; Motoshita, J.; Nakayama, H. Molecular pathobiology of gastric cancer. Scand. J. Surg 2006, 95, 225–231. [Google Scholar]
- Peek, R.M., Jr.; Blaser, M.J. Helicobacter pylori and gastrointestinal tract adenocarcinomas. Nat. Rev. Cancer 2002, 2, 28–37. [Google Scholar]
- Ricci, V.; Romano, M.; Boquet, P. Molecular cross-talk between Helicobacter pylori and human gastric mucosa. World J. Gastroenterol 2011, 17, 1383–1399. [Google Scholar]
- Forman, D. Helicobacter pylori and gastric cancer. Scand. J. Gastroenterol. Suppl 1996, 220, 23–26. [Google Scholar]
- Pilpilidis, I.; Kountouras, J.; Zavos, C.; Katsinelos, P. Upper gastrointestinal carcinogenesis: H. pylori and stem cell cross-talk. J. Surg. Res 2011, 166, 255–264. [Google Scholar]
- Fock, K.M.; Ang, T.L. Epidemiology of Helicobacter pylori infection and gastric cancer in Asia. J. Gastroenterol. Hepatol 2010, 25, 479–486. [Google Scholar]
- Sakamoto, H.; Yoshimura, K.; Saeki, N.; Katai, H.; Shimoda, T.; Matsuno, Y.; Saito, D.; Sugimura, H.; Tanioka, F.; Kato, S.; et al. Genetic variation in PSCA is associated with susceptibility to diffuse-type gastric cancer. Nat. Genet 2008, 40, 730–740. [Google Scholar]
- Saeki, N.; Saito, A.; Choi, I.J.; Matsuo, K.; Ohnami, S.; Totsuka, H.; Chiku, S.; Kuchiba, A.; Lee, Y.S.; Yoon, K.A.; et al. A functional single nucleotide polymorphism in mucin 1, at chromosome 1q22, determines susceptibility to diffuse-type gastric cancer. Gastroenterology 2011, 140, 892–902. [Google Scholar]
- Shi, Y.; Hu, Z.; Wu, C.; Dai, J.; Li, H.; Dong, J.; Wang, M.; Miao, X.; Zhou, Y.; Lu, F.; et al. A genome-wide association study identifies new susceptibility loci for non-cardia gastric cancer at 3q13.31 and 5p13.1. Nat. Genet 2011, 43, 1215–1218. [Google Scholar]
- Abnet, C.C.; Freedman, N.D.; Hu, N.; Wang, Z.; Yu, K.; Shu, X.O.; Yuan, J.M.; Zheng, W.; Dawsey, S.M.; Dong, L.M.; et al. A shared susceptibility locus in PLCE1 at 10q23 for gastric adenocarcinoma and esophageal squamous cell carcinoma. Nat. Genet 2010, 42, 764–767. [Google Scholar]
- Gibson, G. Rare and common variants: Twenty arguments. Nat. Rev. Genet 2012, 13, 135–145. [Google Scholar]
- Palmer, L.J.; Cardon, L.R. Shaking the tree: Mapping complex disease genes with linkage disequilibrium. Lancet 2005, 366, 1223–1234. [Google Scholar]
- Hirakawa, M.; Tanaka, T.; Hashimoto, Y.; Kuroda, M.; Takagi, T.; Nakamura, Y. JSNP: A database of common gene variations in the Japanese population. Nucleic Acids Res 2002, 30, 158–162. [Google Scholar]
- Yoshida, T.; Ono, H.; Kuchiba, A.; Saeki, N.; Sakamoto, H. Genome-wide germline analyses on cancer susceptibility and GeMDBJ database: Gastric cancer as an example. Cancer Sci 2010, 101, 1582–1589. [Google Scholar]
- Miki, D.; Kubo, M.; Takahashi, A.; Yoon, K.A.; Kim, J.; Lee, G.K.; Zo, J.I.; Lee, J.S.; Hosono, N.; Morizono, T.; et al. Variation in TP63 is associated with lung adenocarcinoma susceptibility in Japanese and Korean populations. Nat. Genet 2010, 42, 893–896. [Google Scholar]
- Aoki, A.; Ozaki, K.; Sato, H.; Takahashi, A.; Kubo, M.; Sakata, Y.; Onouchi, Y.; Kawaguchi, T.; Lin, T.H.; Takano, H.; et al. SNPs on chromosome 5p15.3 associated with myocardial infarction in Japanese population. J. Hum. Genet 2011, 56, 47–51. [Google Scholar]
- Hirota, T.; Takahashi, A.; Kubo, M.; Tsunoda, T.; Tomita, K.; Doi, S.; Fujita, K.; Miyatake, A.; Enomoto, T.; Miyagawa, T.; et al. Genome-wide association study identifies three new susceptibility loci for adult asthma in the Japanese population. Nat. Genet 2011, 43, 893–896. [Google Scholar]
- Low, S.K.; Takahashi, A.; Cha, P.C.; Zembutsu, H.; Kamatani, N.; Kubo, M.; Nakamura, Y. Genome-wide association study for intracranial aneurysm in the Japanese population identifies three candidate susceptible loci and a functional genetic variant at EDNRA. Hum. Mol. Genet 2012, 21, 2102–2110. [Google Scholar]
- Onouchi, Y.; Ozaki, K.; Burns, J.C.; Shimizu, C.; Terai, M.; Hamada, H.; Honda, T.; Suzuki, H.; Suenaga, T.; Takeuchi, T.; et al. A genome-wide association study identifies three new risk loci for Kawasaki disease. Nat. Genet 2012, 44, 517–521. [Google Scholar]
- Wu, C.; Wang, G.; Yang, M.; Huang, L.; Yu, D.; Tan, W.; Lin, D. Two genetic variants in prostate stem cell antigen and gastric cancer susceptibility in a Chinese population. Mol. Carcinog 2009, 48, 1131–1138. [Google Scholar]
- Matsuo, K.; Tajima, K.; Suzuki, T.; Kawase, T.; Watanabe, M.; Shitara, K.; Misawa, K.; Ito, S.; Sawaki, A.; Muro, K.; et al. Association of prostate stem cell antigen gene polymorphisms with the risk of stomach cancer in Japanese. Int. J. Cancer 2009, 125, 1961–1964. [Google Scholar]
- Lu, Y.; Chen, J.; Ding, Y.; Jin, G.; Wu, J.; Huang, H.; Deng, B.; Hua, Z.; Zhou, Y.; Shu, Y.; et al. Genetic variation of PSCA gene is associated with the risk of both diffuse- and intestinal-type gastric cancer in a Chinese population. Int. J. Cancer 2010, 127, 2183–2189. [Google Scholar]
- Ou, J.; Li, K.; Ren, H.; Bai, H.; Zeng, D.; Zhang, C. Association and haplotype analysis of prostate stem cell antigen with gastric cancer in Tibetans. DNA Cell Biol 2010, 29, 319–323. [Google Scholar]
- Lochhead, P.; Frank, B.; Hold, G.L.; Rabkin, C.S.; Ng, M.T.; Vaughan, T.L.; Risch, H.A.; Gammon, M.D.; Lissowska, J.; Weck, M.N.; et al. Genetic variation in the prostate stem cell antigen gene and upper gastrointestinal cancer in white individuals. Gastroenterology 2011, 140, 435–441. [Google Scholar]
- Zeng, Z.; Wu, X.; Chen, F.; Yu, J.; Xue, L.; Hao, Y.; Wang, Y.; Chen, M.; Sung, J.J.; Hu, P. Polymorphisms in prostate stem cell antigen gene rs2294008 increase gastric cancer risk in Chinese. Mol. Carcinog 2011, 50, 353–358. [Google Scholar]
- Song, H.R.; Kim, H.N.; Piao, J.M.; Kweon, S.S.; Choi, J.S.; Bae, W.K.; Chung, I.J.; Park, Y.K.; Kim, S.H.; Choi, Y.D.; et al. Association of a common genetic variant in prostate stem-cell antigen with gastric cancer susceptibility in a Korean population. Mol. Carcinog 2011, 50, 871–875. [Google Scholar]
- Sala, N.; Muñoz, X.; Travier, N.; Agudo, A.; Duell, E.J.; Moreno, V.; Overvad, K.; Tjonneland, A.; Boutron-Ruault, M.C.; Clavel-Chapelon, F.; et al. Prostate stem-cell antigen gene is associated with diffuse and intestinal gastric cancer in Caucasians: Results from the EPIC-EURGAST study. Int. J. Cancer 2011, 130, 2417–2427. [Google Scholar]
- Ng, W.; Loh, A.X.; Teixeira, A.S.; Pereira, S.P.; Swallow, D.M. Genetic regulation of MUC1 alternative splicing in human tissues. Br. J. Cancer 2008, 99, 978–985. [Google Scholar]
- Song, H.R.; Kim, H.N.; Kweon, S.S.; Choi, J.S.; Shim, H.J.; Cho, S.H.; Chung, I.J.; Park, Y.K.; Kim, S.H.; Choi, Y.D.; et al. Common genetic variants at 1q22 and 10q23 and gastric cancer susceptibility in a Korean population. Tumour Biol 2014, 35, 3133–3137. [Google Scholar]
- Xu, Q.; Yuan, Y.; Sun, L.P.; Gong, Y.H.; Xu, Y.; Yu, X.W.; Dong, N.N.; Lin, G.D.; Smith, P.N.; Li, R.W. Risk of gastric cancer is associated with the MUC1 568 A/G polymorphism. Int. J. Oncol 2009, 35, 1313–1320. [Google Scholar]
- Li, F.; Zhong, M.Z.; Li, J.H.; Liu, W.; Li, B. Case-control study of single nucleotide polymorphisms of PSCA and MUC1 genes with gastric cancer in a Chinese. Asian Pac. J. Cancer Prev 2012, 13, 2593–2596. [Google Scholar]
- Jia, Y.; Persson, C.; Hou, L.; Zheng, Z.; Yeager, M.; Lissowska, J.; Chanock, S.J.; Chow, W.H.; Ye, W. A comprehensive analysis of common genetic variation in MUC1, MUC5AC, MUC6 genes and risk of stomach cancer. Cancer Causes Control 2010, 21, 313–321. [Google Scholar]
- Palmer, A.J.; Lochhead, P.; Hold, G.L.; Rabkin, C.S.; Chow, W.H.; Lissowska, J.; Vaughan, T.L.; Berry, S.; Gammon, M.; Risch, H.; et al. Genetic variation in C20orf54, PLCE1 and MUC1 and the risk of upper gastrointestinal cancers in Caucasian populations. Eur. J. Cancer Prev 2013, 21, 541–544. [Google Scholar]
- Zheng, L.; Zhu, C.; Gu, J.; Xi, P.; Du, J.; Jin, G. Functional polymorphism rs4072037 in MUC1 gene contributes to the susceptibility to gastric cancer: evidence from pooled 6580 cases and 10,324 controls. Mol. Biol. Rep 2013, 40, 5791–5796. [Google Scholar]
- Carvalho, F.; Seruca, R.; David, L.; Amorim, A.; Seixas, M.; Bennett, E.; Clausen, H.; Sobrinho-Simões, M. MUC1 gene polymorphism and gastric cancer—An epidemiological study. Glycoconj. J 1997, 14, 107–111. [Google Scholar]
- Ho, S.B.; Niehans, G.A.; Lyftogt, C.; Yan, P.S.; Cherwitz, D.L.; Gum, E.T.; Dahiya, R.; Kim, Y.S. Heterogeneity of mucin gene expression in normal and neoplastic tissues. Cancer Res 1993, 53, 641–651. [Google Scholar]
- Ho, S.B.; Shekels, L.L.; Toribara, N.W.; Kim, Y.S.; Lyftogt, C.; Cherwitz, D.L.; Niehans, G.A. Mucin gene expression in normal, preneoplastic, and neoplastic human gastric epithelium. Cancer Res 1995, 55, 2681–2690. [Google Scholar]
- Utsunomiya, T.; Yonezawa, S.; Sakamoto, H.; Kitamura, H.; Hokita, S.; Aiko, T.; Tanaka, S.; Irimura, T.; Kim, Y.S.; Sato, E. Expression of MUC1 and MUC2 mucins in gastric carcinomas: its relationship with the prognosis of the patients. Clin. Cancer Res 1998, 4, 2605–2614. [Google Scholar]
- Reis, C.A.; David, L.; Seixas, M.; Burchell, J.; Sobrinho-Simões, M. Expression of fully and under-glycosylated forms of MUC1 mucin in gastric carcinoma. Int. J. Cancer 1998, 79, 402–410. [Google Scholar]
- Lee, H.S.; Lee, H.K.; Kim, H.S.; Yang, H.K.; Kim, Y.I.; Kim, W.H. MUC1, MUC2, MUC5AC, and MUC6 expressions in gastric carcinomas: Their roles as prognostic indicators. Cancer 2001, 92, 1427–1434. [Google Scholar]
- Wang, R.Q.; Fang, D.C. Alterations of MUC1 and MUC3 expression in gastric carcinoma: Relevance to patient clinicopathological features. J. Clin. Pathol 2003, 56, 378–384. [Google Scholar]
- Wang, J.Y.; Chang, C.T.; Hsieh, J.S.; Lee, L.W.; Huang, T.J.; Chai, C.Y.; Lin, S.R. Role of MUC1 and MUC5AC expressions as prognostic indicators in gastric carcinomas. J. Surg. Oncol 2003, 83, 253–260. [Google Scholar]
- Barresi, V.; Vitarelli, E.; Grosso, M.; Tuccari, G.; Barresi, G. Relationship between immunoexpression of mucin peptide cores MUC1 and MUC2 and Lauren’s histologic subtypes of gastric carcinomas. Eur. J. Histochem 2006, 50, 301–309. [Google Scholar]
- Kocer, B.; Soran, A.; Kiyak, G.; Erdogan, S.; Eroglu, A.; Bozkurt, B.; Solak, C.; Cengiz, O. Prognostic significance of mucin expression in gastric carcinoma. Dig. Dis. Sci 2004, 49, 954–964. [Google Scholar]
- Terada, T. An immunohistochemical study of primary signet-ring cell carcinoma of the stomach and colorectum: II. Expression of MUC1, MUC2, MUC5AC, and MUC6 in normal mucosa and in 42 cases. Int. J. Clin. Exp. Pathol 2013, 6, 613–621. [Google Scholar]
- Tanaka, M.; Kitajima, Y.; Sato, S.; Miyazaki, K. Combined evaluation of mucin antigen and E-cadherin expression may help select patients with gastric cancer suitable for minimally invasive therapy. Br. J. Surg 2003, 90, 95–101. [Google Scholar]
- Ohno, T.; Aihara, R.; Kamiyama, Y.; Mochiki, E.; Asao, T.; Kuwano, H. Prognostic significance of combined expression of MUC1 and adhesion molecules in advanced gastric cancer. Eur. J. Cancer 2006, 42, 256–263. [Google Scholar]
- Owen, D.A. Stomach. In Histology for Pathologists; Mills, S.E., Ed.; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2007; p. 589. [Google Scholar]
- Reis, C.A.; David, L.; Correa, P.; Carneiro, F.; de Bolós, C.; Garcia, E.; Mandel, U.; Clausen, H.; Sobrinho-Simões, M. Intestinal metaplasia of human stomach displays distinct patterns of mucin (MUC1, MUC2, MUC5AC, and MUC6) expression. Cancer Res 1999, 59, 1003–1007. [Google Scholar]
- Vernygorodskyi, S. Immunohistochemical evaluation of mucin expression in precancerous tissue of stomach. Exp. Oncol 2013, 35, 114–117. [Google Scholar]
- Jonckheere, N.; van Seuningen, I. The membrane-bound mucins: From cell signalling to transcriptional regulation and expression in epithelial cancers. Biochimie 2010, 92, 1–11. [Google Scholar]
- Bafna, S.; Kaur, S.; Batra, S.K. Membrane-bound mucins: the mechanistic basis for alterations in the growth and survival of cancer cells. Oncogene 2010, 29, 2893–2904. [Google Scholar]
- Kufe, D.W. Mucins in cancer: Function, prognosis and therapy. Nat. Rev. Cancer 2009, 9, 874–885. [Google Scholar]
- Gendler, S.J. MUC1, the renaissance molecule. Mammary Gland Biol. Neoplasia 2001, 6, 339–353. [Google Scholar]
- McAuley, J.L.; Linden, S.K.; Png, C.W.; King, R.M.; Pennington, H.L.; Gendler, S.J.; Florin, T.H.; Hill, G.R.; Korolik, V.; McGuckin, M.A. MUC1 cell surface mucin is a critical element of the mucosal barrier to infection. J. Clin. Investig 2007, 117, 2313–2324. [Google Scholar]
- Lindén, S.K.; Sheng, Y.H.; Every, A.L.; Miles, K.M.; Skoog, E.C.; Florin, T.H.; Sutton, P.; McGuckin, M.A. MUC1 limits Helicobacter pylori infection both by steric hindrance and by acting as a releasable decoy. PLoS Pathog 2009, 5, e1000617. [Google Scholar]
- McGuckin, M.A.; Every, A.L.; Skene, C.D.; Linden, S.K.; Chionh, Y.T.; Swierczak, A.; McAuley, J.; Harbour, S.; Kaparakis, M.; Ferrero, R.; et al. Muc1 mucin limits both Helicobacter pylori colonization of the murine gastric mucosa and associated gastritis. Gastroenterology 2007, 133, 1210–1218. [Google Scholar]
- Vinall, L.E.; King, M.; Novelli, M.; Green, C.A.; Daniels, G.; Hilkens, J.; Sarner, M.; Swallow, D.M. Altered expression and allelic association of the hypervariable membrane mucin MUC1 in Helicobacter pylori gastritis. Gastroenterology 2002, 123, 41–49. [Google Scholar]
- Li, M.; Huang, L.; Qiu, H.; Fu, Q.; Li, W.; Yu, Q.; Sun, L.; Zhang, L.; Hu, G.; Hu, J.; et al. Helicobacter pylori infection synergizes with three inflammation-related genetic variants in the GWASs to increase risk of gastric cancer in a Chinese population. PLoS One 2013, 8, e74976. [Google Scholar]
- Carson, D.D. The cytoplasmic tail of MUC1: A very busy place. Sci. Signal 2008, 1. [Google Scholar] [CrossRef]
- Behrens, M.E.; Grandgenett, P.M.; Bailey, J.M.; Singh, P.K.; Yi, C.H.; Yu, F.; Hollingsworth, M.A. The reactive tumor microenvironment: MUC1 signaling directly reprograms transcription of CTGF. Oncogene 2010, 29, 5667–5677. [Google Scholar]
- Masckauchán, T.N.; Shawber, C.J.; Funahashi, Y.; Li, C.M.; Kitajewski, J. Wnt/beta-catenin signaling induces proliferation, survival and interleukin-8 in human endothelial cells. Angiogenesis 2005, 8, 43–51. [Google Scholar]
- Murata-Kamiya, N.; Kurashima, Y.; Teishikata, Y.; Yamahashi, Y.; Saito, Y.; Higashi, H.; Aburatani, H.; Akiyama, T.; Peek, R.M., Jr.; Azuma, T.; et al. Helicobacter pylori CagA interacts with E-cadherin and deregulates the β-catenin signal that promotes intestinal transdifferentiation in gastric epithelial cells. Oncogene 2007, 26, 4617–4626. [Google Scholar]
- Lévy, L.; Neuveut, C.; Renard, C.A.; Charneau, P.; Branchereau, S.; Gauthier, F.; van Nhieu, J.T.; Cherqui, D.; Petit-Bertron, A.F.; Mathieu, D.; et al. Transcriptional activation of interleukin-8 by β-catenin-Tcf4. J. Biol. Chem 2002, 277, 42386–42393. [Google Scholar]
- Guang, W.; Twaddell, W.S.; Lillehoj, E.P. Molecular Interactions between MUC1 Epithelial Mucin, β-Catenin, and CagA Proteins. Front. Immunol 2012, 3. [Google Scholar] [CrossRef]
- Park, Y.S.; Guang, W.; Blanchard, T.G.; Kim, K.C.; Lillehoj, E.P. Suppression of IL-8 production in gastric epithelial cells by MUC1 mucin and peroxisome proliferator-associated receptor-γ. Am. J. Physiol. Gastrointest. Liver Physiol 2012, 303, G765–G774. [Google Scholar]
- Guang, W.; Czinn, S.J.; Blanchard, T.G.; Kim, K.C.; Lillehoj, E.P. Genetic regulation of MUC1 expression by Helicobacter pylori in gastric cancer cells. Biochem. Biophys. Res. Commun 2014, 445, 145–150. [Google Scholar]
- Perrais, M.; Rousseaux, C.; Ducourouble, M.P.; Courcol, R.; Vincent, P.; Jonckheere, N.; van Seuningen, I. Helicobacter pylori urease and flagellin alter mucin gene expression in human gastric cancer cells. Gastric Cancer 2014, 17, 235–246. [Google Scholar]
- Navabi, N.; Johansson, M.E.; Raghavan, S.; Lindén, S.K. Helicobacter pylori infection impairs the mucin production rate and turnover in the murine gastric mucosa. Infect. Immun 2013, 81, 829–837. [Google Scholar]
- Wei, X.; Xu, H.; Kufe, D. Human mucin 1 oncoprotein represses transcription of the p53 tumor suppressor gene. Cancer Res 2007, 67, 1853–1858. [Google Scholar]
- Weinberg, R.A. The Biology of Cancer; Garland Science: New York, NY, USA, 2007. [Google Scholar]
- Raina, D.; Kharbanda, S.; Kufe, D. The MUC1 oncoprotein activates the anti-apoptotic phosphoinositide 3-kinase/Akt and Bcl-xL pathways in rat 3Y1 fibroblasts. J. Biol. Chem 2004, 279, 20607–20612. [Google Scholar]
- Raina, D.; Ahmad, R.; Kumar, S.; Ren, J.; Yoshida, K.; Kharbanda, S.; Kufe, D. MUC1 oncoprotein blocks nuclear targeting of c-Abl in the apoptotic response to DNA damage. EMBO J 2006, 25, 3774–3783. [Google Scholar]
- Rahn, J.J.; Chow, J.W.; Horne, G.J.; Mah, B.K.; Emerman, J.T.; Hoffman, P.; Hugh, J.C. MUC1 mediates transendothelial migration in vitro by ligating endothelial cell ICAM-1. Clin. Exp. Metastasis 2005, 22, 475–483. [Google Scholar]
- Hikita, S.T.; Kosik, K.S.; Clegg, D.O.; Bamdad, C. MUC1* mediates the growth of human pluripotent stem cells. PLoS One 2008, 3, e3312. [Google Scholar]
- Stroopinsky, D.; Rosenblatt, J.; Ito, K.; Mills, H.; Yin, L.; Rajabi, H.; Vasir, B.; Kufe, T.; Luptakova, K.; Arnason, J.; et al. MUC1 is a potential target for the treatment of acute myeloid leukemia stem cells. Cancer Res 2013, 73, 5569–5579. [Google Scholar]
- Mehla, K.; Singh, P.K. MUC1: A novel metabolic master regulator. Biochim. Biophys. Acta 2014, 1845, 126–135. [Google Scholar]
- Saeki, N.; Ono, H.; Sakamoto, H.; Yoshida, T. Genetic factors related to gastric cancer susceptibility identified using a genome-wide association study. Cancer Sci 2013, 104, 1–8. [Google Scholar]
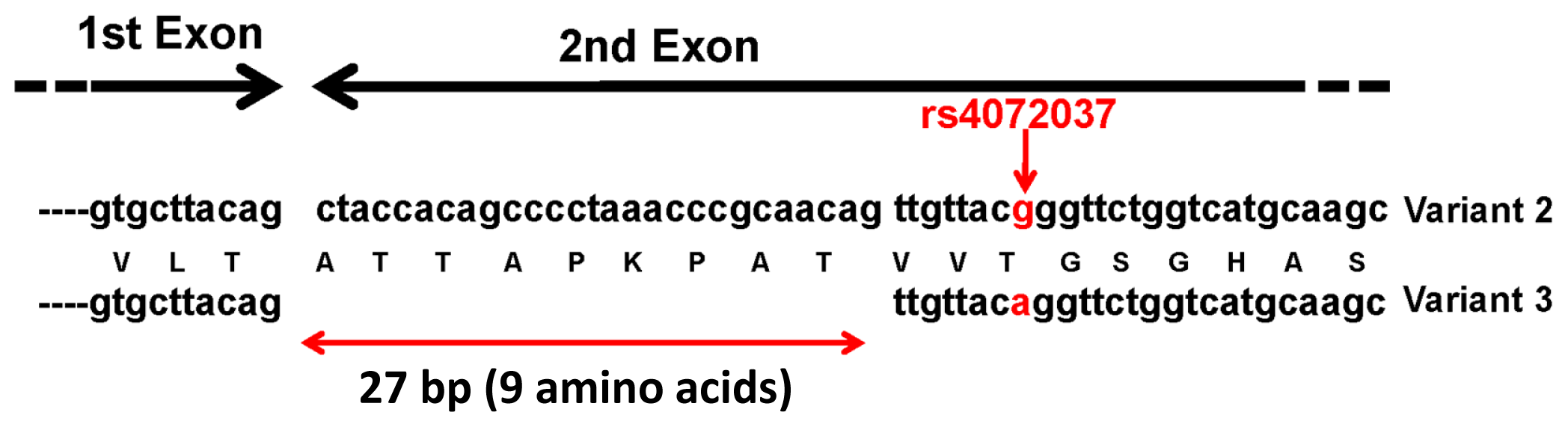


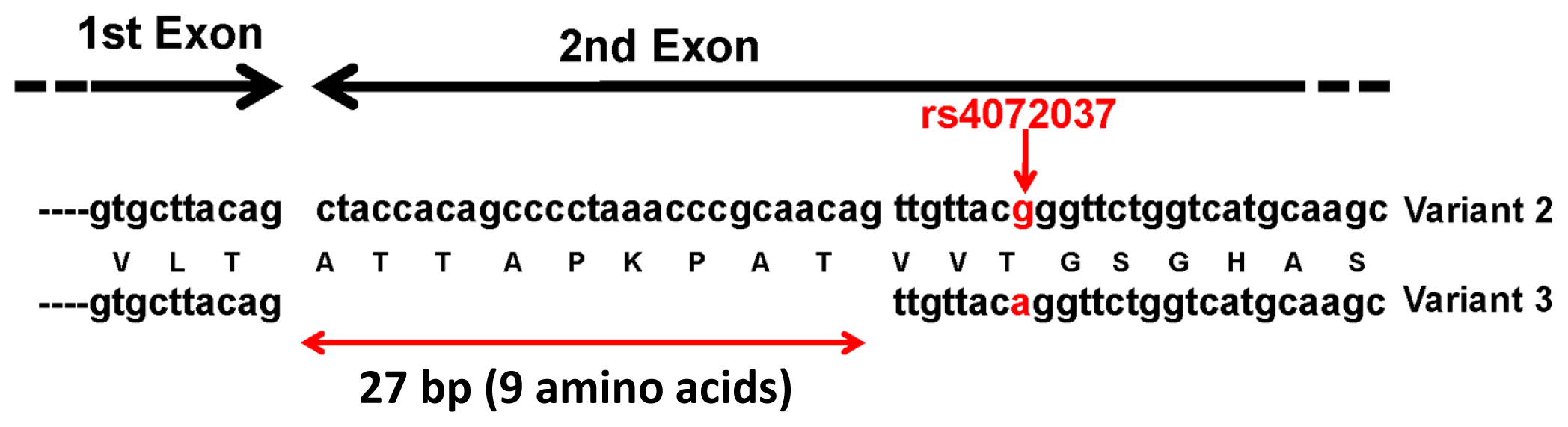{kind=link}
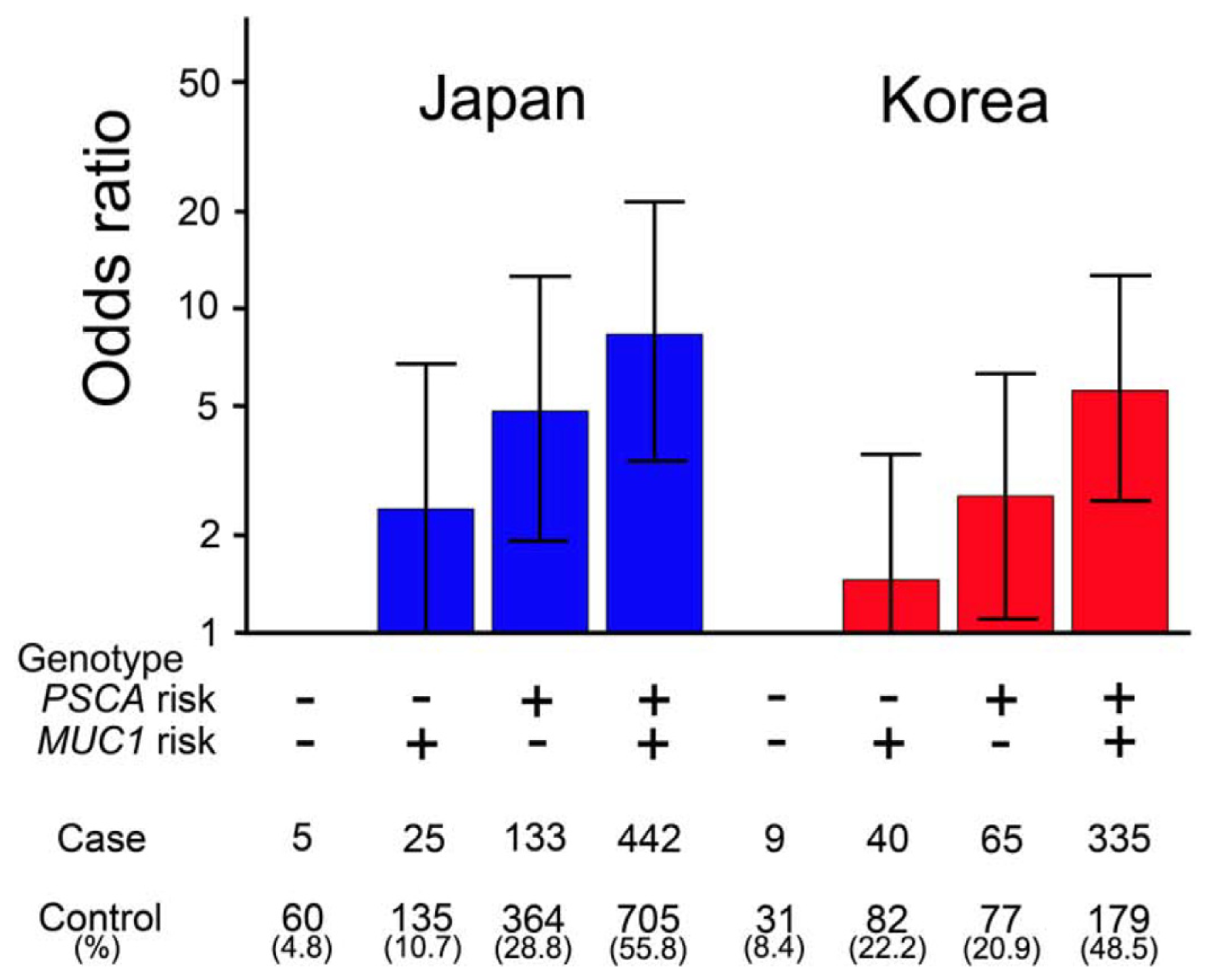{kind=link}
| SNPs (Major/Minor) | Risk Allele | Odds Ratio (95% CI) and Genotype | p Value | Ethnic | Cancer Type | Ref. |
|---|---|---|---|---|---|---|
| rs4072037 (A/G) | A | 1.62 (1.32–1.99) # A to G, allelic | 4.04 × 10−6 | Japanese | DGC | [9] |
| rs4072037 (A/G) | A | 1.74 (1.26–2.39) # A to G, allelic | 7.82 × 10−4 | Korean | DGC | [9] |
| rs4072037 (A/G) | A | 0.78 (0.67–0.91) AG to AA | 0.031 | Korean | All | [30] |
| rs4072037 (A/G) | A | 0.72 (0.62–0.85) G to A, allelic | 5.74 × 10−5 | Chinese | Non-cardia | [11] |
| rs4072037 (A/G) | A | 0.75 (0.65–0.87) G to A, allelic | 9.45 × 10−5 | Chinese | Cardia | [11] |
| rs4072037 (A/G) | A | 0.73 G to A, allelic | 1.0 × 10−4 | Chinese | Non-cardia | [10] |
| rs4072037 (A/G) | A | 1.81 AA to AG + GG | 0.031 | Chinese | All | [31] |
| rs2070803 (G/A) | G | 0.46 (0.32–0.67) AA + AG to GG | <0.001 | Chinese | All | [32] |
| rs4072037 (G/A) | A | 2.20 (1.41–3.44) AA to GG | <0.01 | Caucasian | All | [33] |
| rs4072037 (G/A) | A | 0.5 (0.3–0.9) AG to AA | - | Caucasian | Cardia | [34] |
| rs4072037 (G/A) | A | 0.4 (0.2–0.9) AG to AA | - | Caucasian | Non-cardia, Intestinal | [34] |
| Study | MUC1 Staining | Correlation to Clinical Information | ||||||
|---|---|---|---|---|---|---|---|---|
| Intestinal | Diffuse | Intestinal + Diffuse | Note | |||||
| Case No. | (%) | Case No. | (%) | Case No. | (%) | |||
| Ho, et al. [38] | - | - | - | - | 25/33 | (75.8) | - | - |
| Reis, et al. [40] | 31/31 | (100) | 24/24 | (100) | - | - | fully glycosylated MUC1 | lymphatic invasion *, nodal metastasis *, advanced stage |
| - | 73/90 | (81.1) | 30/49 | (61.2) | - | - | under-glycosylated MUC1 | wall penetration, lymphatic invasion *, nodal metastasis, advanced stage |
| Utsunomiya, et al. [39] | (60/68) | (88) | (45/68) | (66) | - | - | fully glycosylated MUC1 | worse prognosis * |
| Lee, et al. [41] | 37/113 | (32.7) | 28/159 | (17.6) | - | - | - | worse prognosis * |
| Wang, et al. [42] | 13/21 | (61.9) | 11/17 | (64.7) | - | - | - | better prognosis |
| Wang, et al. [43] | 14/26 | (53.8) | 30/44 | (68.2) | - | - | - | worse prognosis * |
| Kocer, et al. [45] | 10/16 | (62.5) | 13/19 | (68.4) | - | - | - | worse prognosis * |
| Barresi, et al. [44] | 23/27 | (85.2) | 3/10 | (30) | - | - | - | - |
| Terada, et al. [46] | - | - | 3/30 | (10) | - | - | signet-ring cell carcinoma | - |
| Factors | GC Risk | ||
|---|---|---|---|
| MUC1 polymorphism | rs4072037 | GG, AG | AA |
| tandem-repeat [59] | LL, LS | SS | |
| splicing variant [9] | 2/2, 2/3 | 3/3 | |
| HP infection | seronegative | 1.00 (reference) | 2.46 (1.42–4.27) # |
| seropositive | 2.30 (1.23–4.31) # | 3.95 (2.29–6.79) # | |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Saeki, N.; Sakamoto, H.; Yoshida, T. Mucin 1 Gene (MUC1) and Gastric-Cancer Susceptibility. Int. J. Mol. Sci. 2014, 15, 7958-7973. https://doi.org/10.3390/ijms15057958
Saeki N, Sakamoto H, Yoshida T. Mucin 1 Gene (MUC1) and Gastric-Cancer Susceptibility. International Journal of Molecular Sciences. 2014; 15(5):7958-7973. https://doi.org/10.3390/ijms15057958
Chicago/Turabian StyleSaeki, Norihisa, Hiromi Sakamoto, and Teruhiko Yoshida. 2014. "Mucin 1 Gene (MUC1) and Gastric-Cancer Susceptibility" International Journal of Molecular Sciences 15, no. 5: 7958-7973. https://doi.org/10.3390/ijms15057958