Genistein in 1:1 Inclusion Complexes with Ramified Cyclodextrins: Theoretical, Physicochemical and Biological Evaluation



 ,
,  ,
, 
Abstract
:1. Introduction
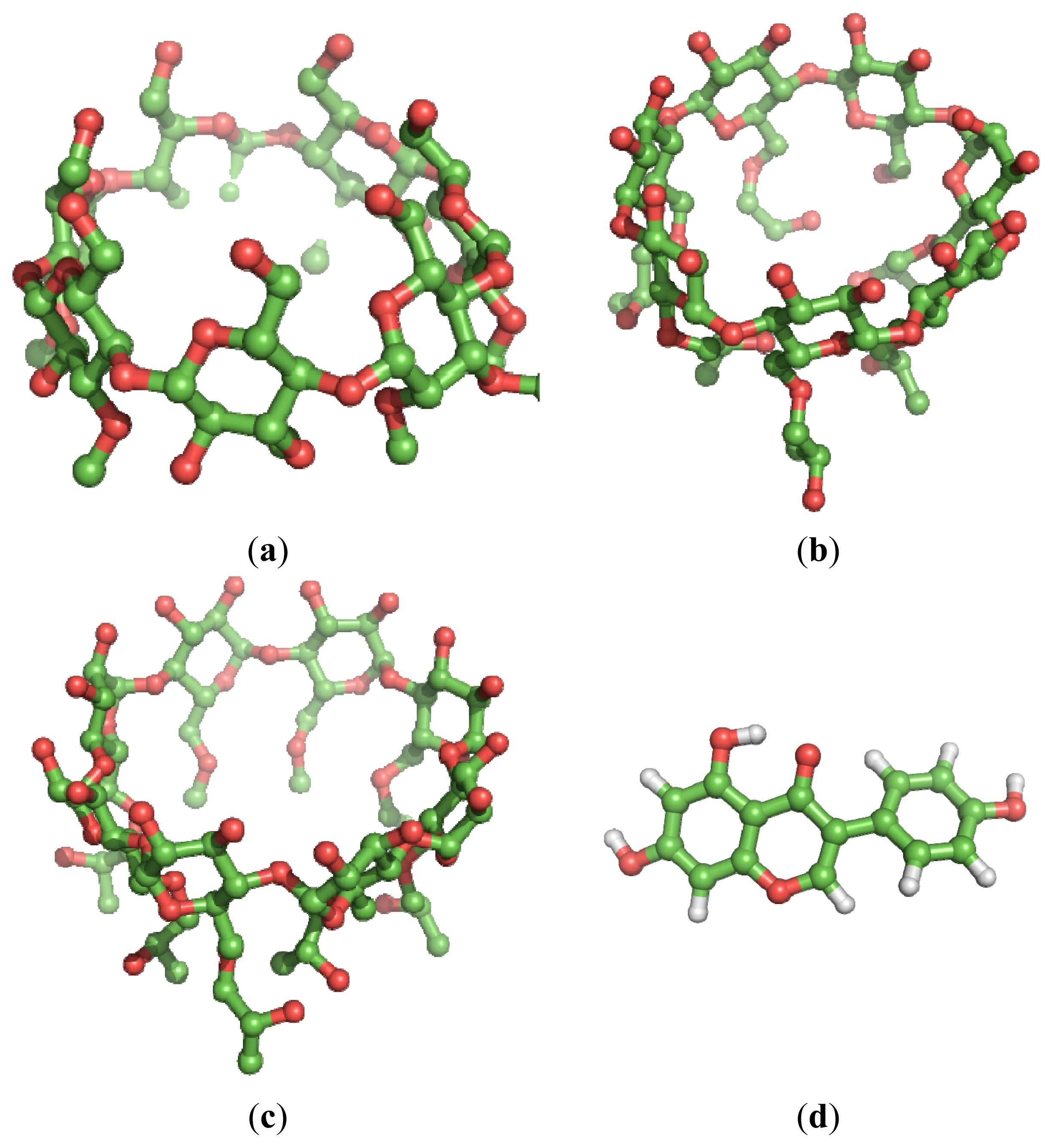
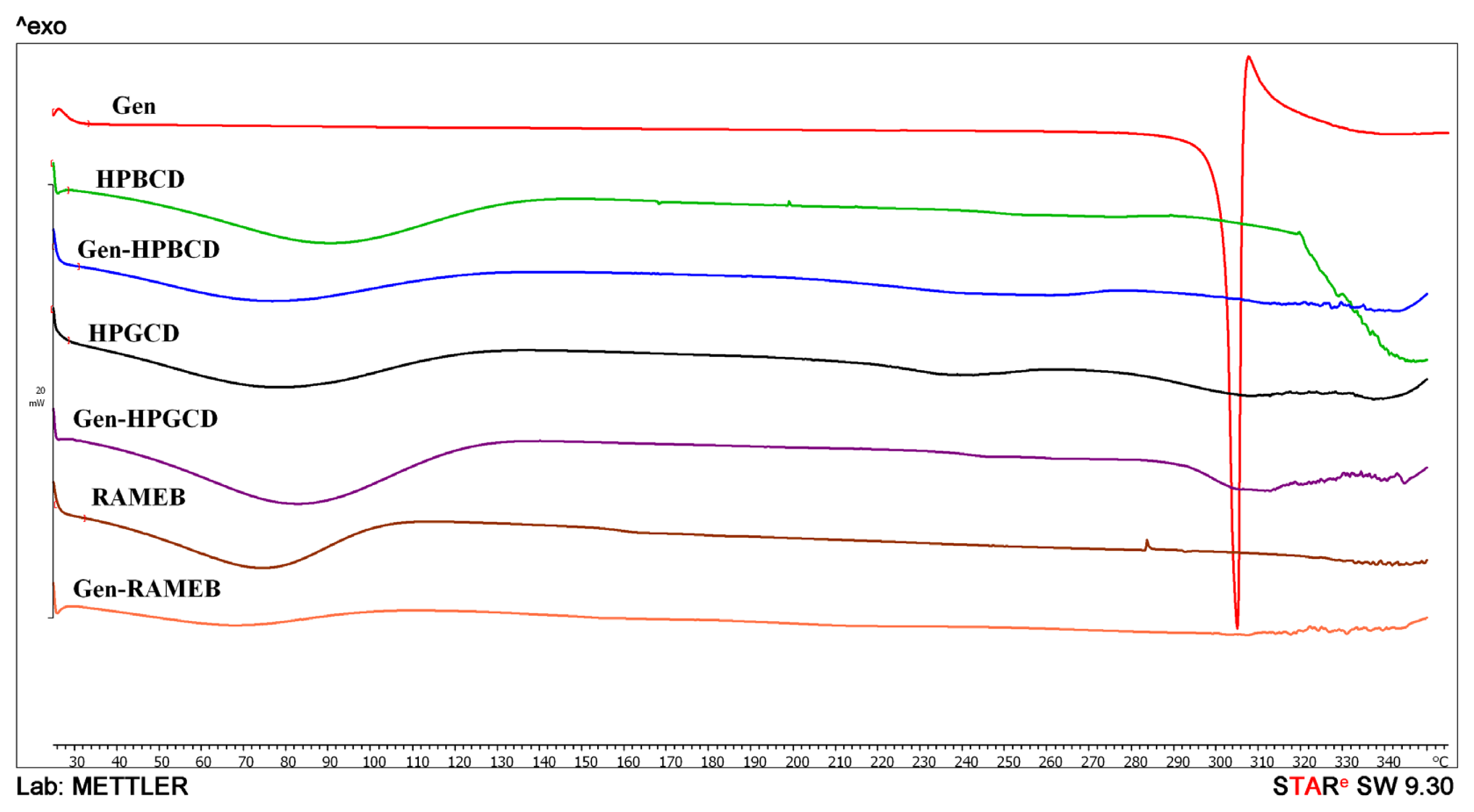
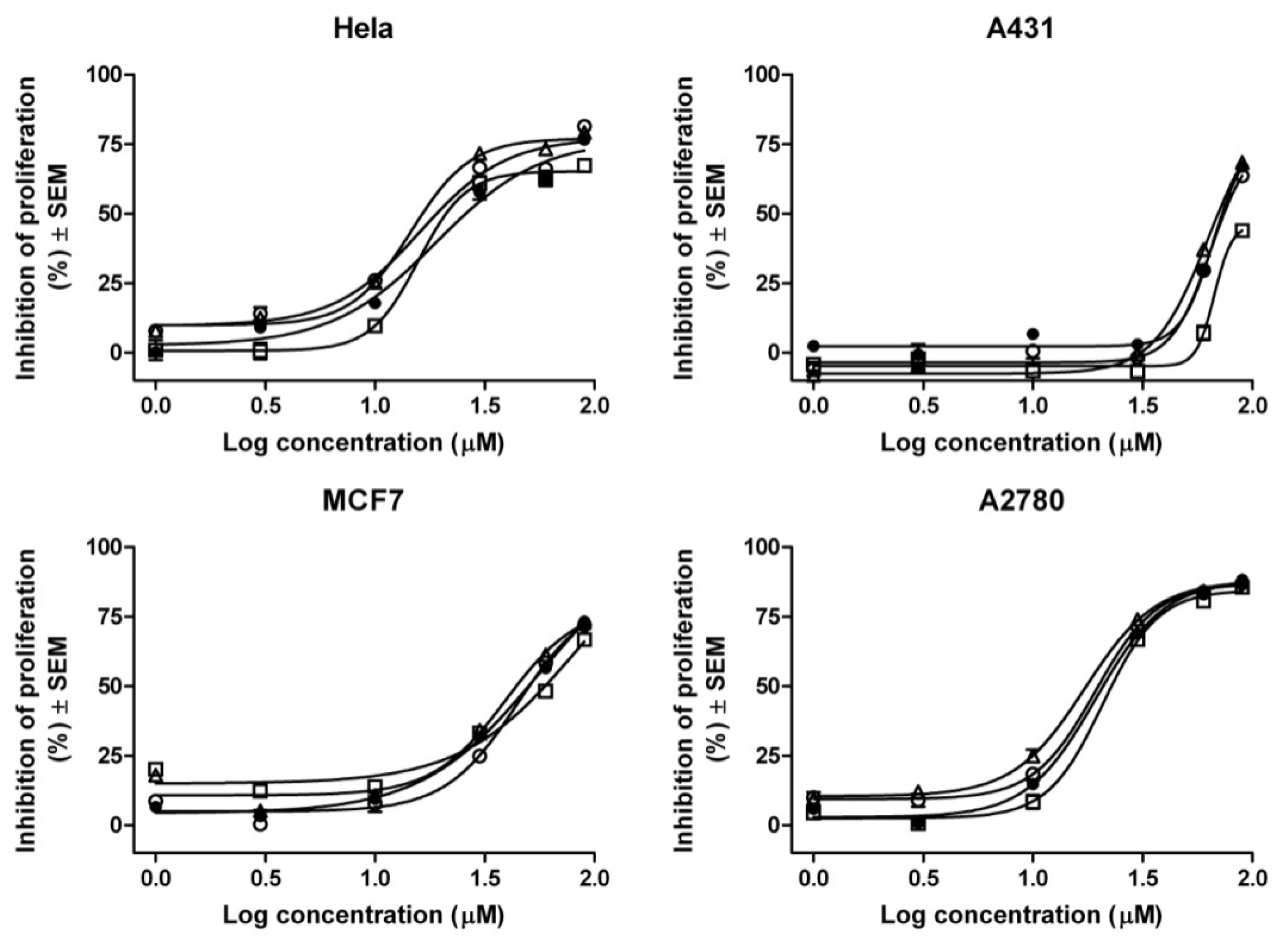
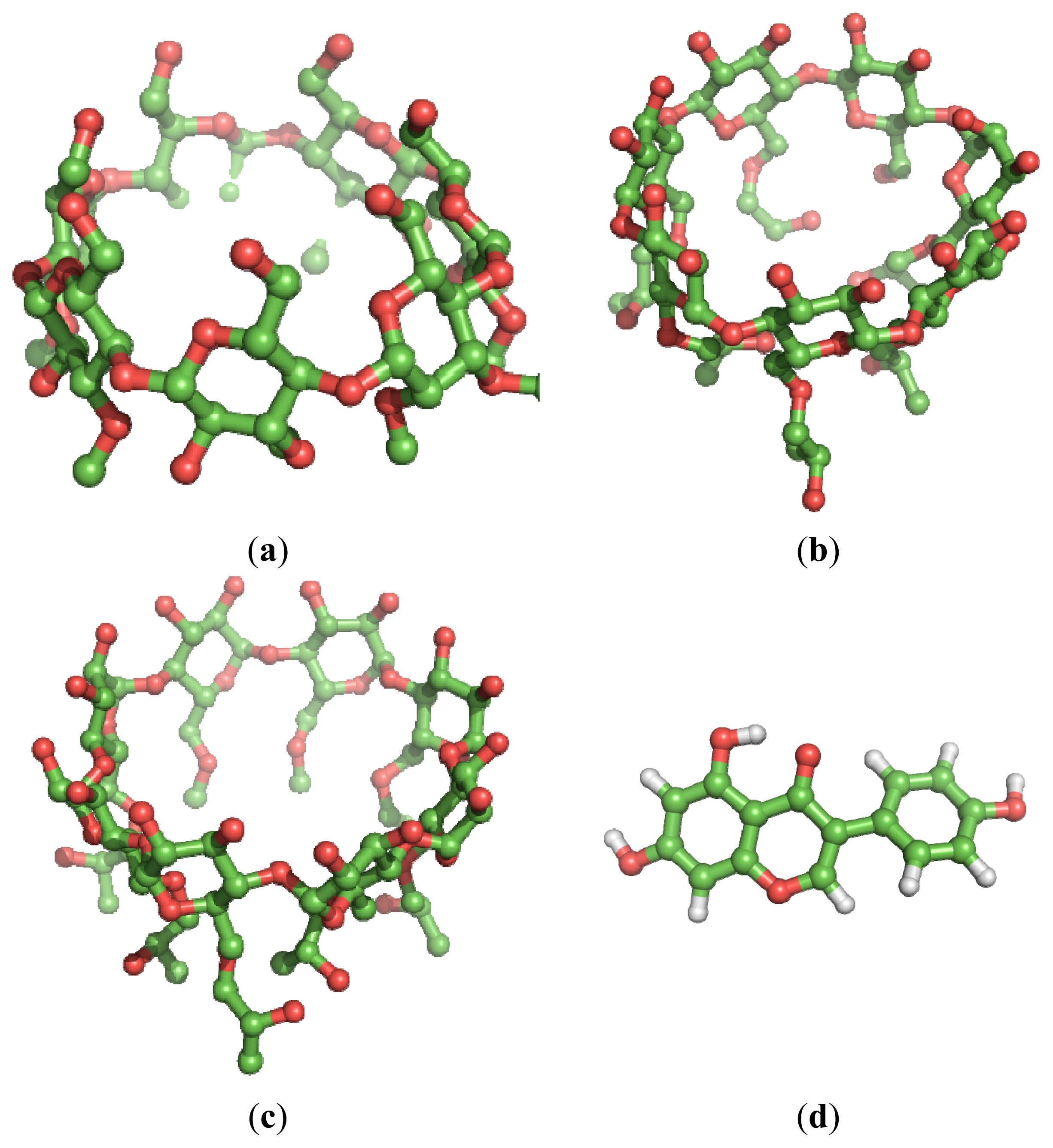
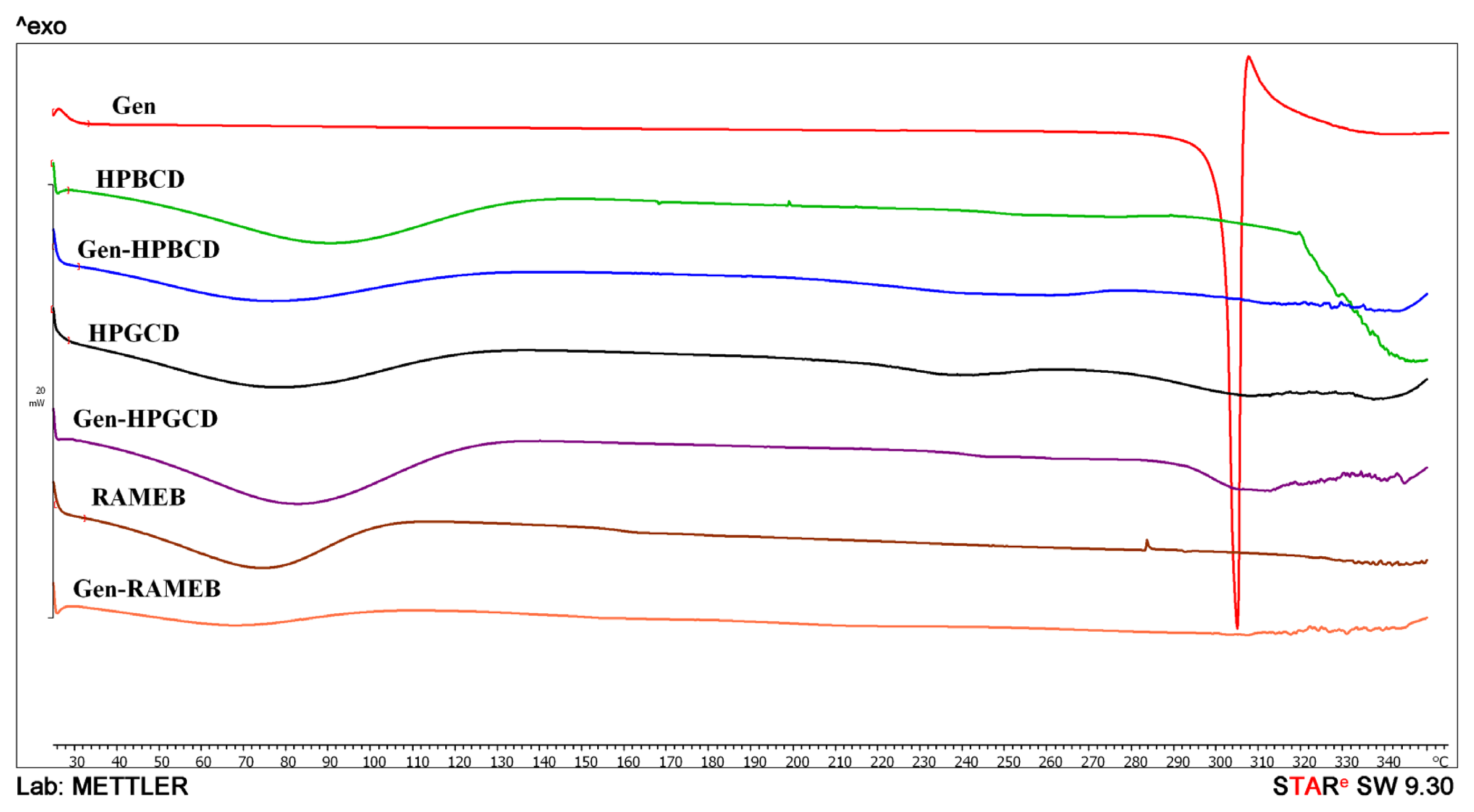
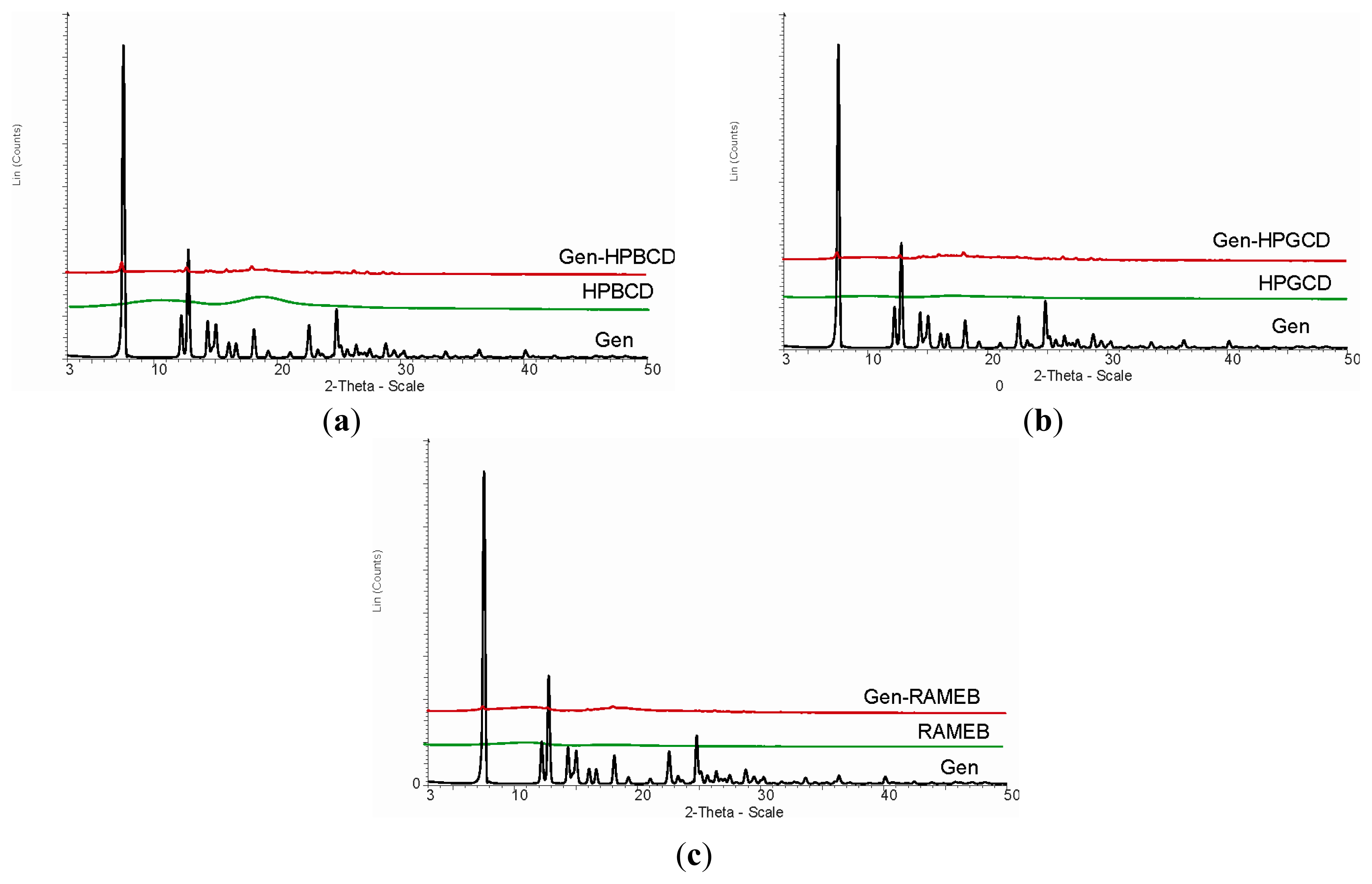
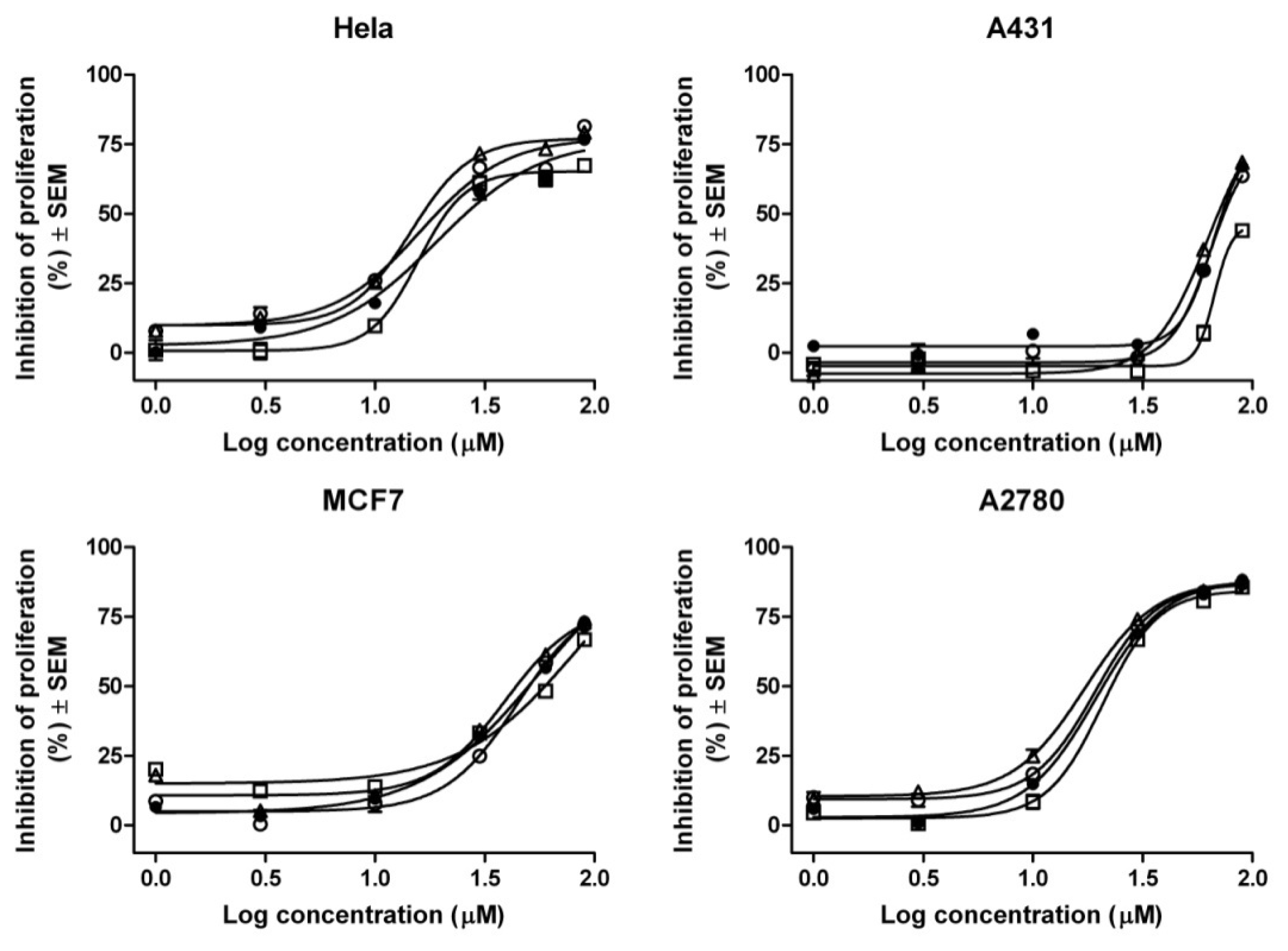
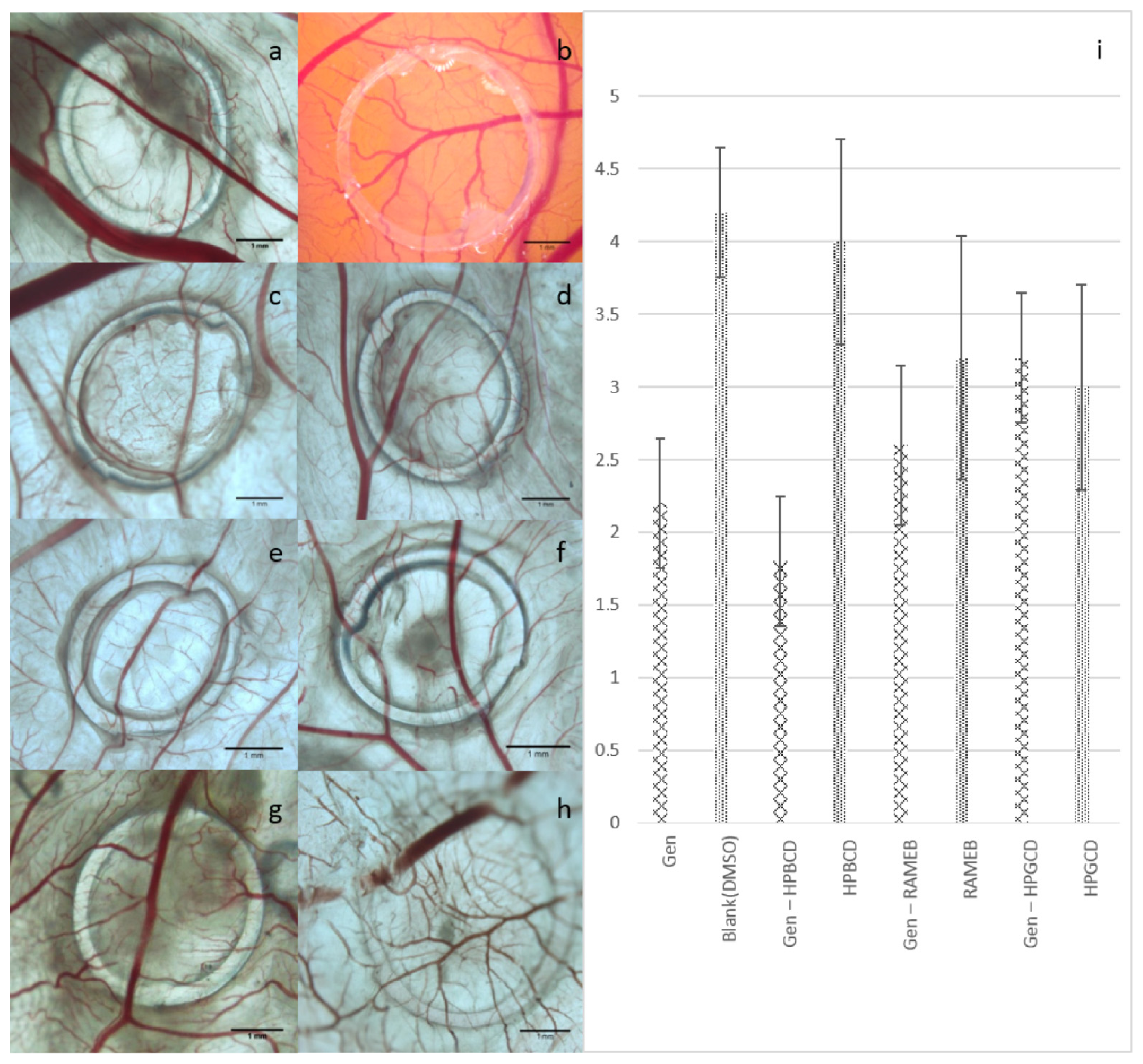
2. Results and Discussion
3. Experimental Section
3.1. Reagents
3.2. Computational Details
3.3. Complexes Preparation
3.4. Phase Solubility Studies
3.5. Differential Scanning Calorimetry (DSC)
3.6. X-ray Diffraction
3.7. Scanning Electron Microscopy (SEM) Assay
3.8. MTT in Vitro Analysis
3.9. In Vitro Antibacterial Activity
3.10. Disk Diffusion Method
3.11. Dilution Assay for Minimal Inhibitory Concentration (MIC) Determinations
3.12. The Chorioallantoic Membrane Assay (CAM Assay)
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Rahman, S.; Islam, R.; Swaraz, A.; Ansari, A.; Parvez, A.K.; Paul, D.K. An insight on genistein as potential pharmacological and therapeutic agent. Asian Pac. J. Trop. Biomed 2012, 2, S1924–S1937. [Google Scholar]
- Taylor, C.K.; Levy, R.M.; Elliott, J.C.; Burnett, B.P. The effect of genistein aglycone on cancer and cancer risk: A review of in vitro, preclinical, and clinical studies. Nutr. Rev 2009, 67, 398–415. [Google Scholar]
- Polkowski, K.; Mazurek, P.A. Biological properties of genistein. A review of in vitro and in vivo data. Acta Pol. Pharm 2000, 57, 135–155. [Google Scholar]
- Pavese, J.M.; Farmer, R.L.; Bergan, R.C. Inhibition of cancer cell invasion and metastasis by genistein. Cancer Metastasis Rev 2010, 29, 465–482. [Google Scholar]
- Danciu, C.; Soica, C.; Csanyi, E.; Ambrus, R.; Feflea, S.; Peev, C.; Dehelean, C. Changes in the anti-inflammatory activity of soy isoflavonoid genistein versus genistein incorporated in two types of cyclodextrin derivatives. Chem. Cent. J 2012, 6, 58. [Google Scholar]
- Hoffmann, D. Medical herbalism: The Science and Practice of Herbal Medicine; Inner Traditions, Bear & Co: Rochester, VT, USA, 2003. [Google Scholar]
- Barrett, J.R. The science of soy: What do we really know? Environ. Health Perspect 2006, 114, A352–A358. [Google Scholar]
- Qin, W.; Zhu, W.; Shi, H.; Hewett, J.E.; Ruhlen, R.L.; MacDonald, R.S.; Rottinghaus, G.E.; Chen, Y.C.; Sauter, E.R. Soy isoflavones have an antiestrogenic effect and alter mammary promoter hypermethylation in healthy premenopausal women. Nutr. Cancer 2009, 61, 238–244. [Google Scholar]
- Gercel-Taylor, C.; Feitelson, A.K.; Taylor, D.D. Inhibitory effect of genistein and daidzein on ovarian cancer cell growth. Anticancer Res 2004, 24, 795–800. [Google Scholar]
- Kronenberg, F. Menopausal hot flashes: A review of physiology and biosociocultural perspective on methods of assessment. J. Nutr 2010, 140, 1380S–1385S. [Google Scholar]
- Evans, M.; Elliott, J.G.; Sharma, P.; Berman, R.; Guthrie, N. The effect of synthetic genistein on menopause symptom management in healthy postmenopausal women: A multi-center, randomized, placebo-controlled study. Maturitas 2011, 68, 189–196. [Google Scholar]
- Altavilla, D.; Crisafulli, A.; Marini, H.; Esposito, M.; D’Anna, R.; Corrado, F.; Bitto, A.; Squadrito, F. Cardiovascular effects of the phytoestrogen genistein. Curr. Med. Chem. Cardiovasc. Hematol. Agents 2004, 2, 179–186. [Google Scholar]
- Wei, H.; Saladi, R.; Lu, Y.; Wang, Y.; Palep, S.R.; Moore, J.; Phelps, R.; Shyong, E.; Lebwohl, M.G. Isoflavone genistein: Photoprotection and clinical implications in dermatology. J. Nutr 2003, 133, 3811S–3819S. [Google Scholar]
- Foti, P.; Erba, D.; Riso, P.; Spadafranca, A.; Criscuoli, F.; Testolin, G. Comparison between daidzein and genistein antioxidant activity in primary and cancer lymphocytes. Arch. Biochem. Biophys 2005, 433, 421–427. [Google Scholar]
- Gao, F.; Wei, D.; Bian, T.; Xie, P.; Zou, J.; Mu, H.; Zhang, B.; Zhou, X. Genistein attenuated allergic airway inflammation by modulating the transcription factors T-bet, GATA-3 and STAT-6 in a murine model of asthma. Pharmacology 2012, 89, 229–236. [Google Scholar]
- Ruijter, J.; Valstar, M.J.; Narajczyk, M.; Wegrzyn, G.; Kulik, W.; Ijist, L.; Wagemans, T.; van der Wal, W.M.; Wijburg, F.A. Genistein in Sanfilippo disease: A randomized controlled crossover trial. Ann. Neurol 2012, 71, 110–120. [Google Scholar]
- Uckun, F.M.; Evans, W.E.; Forsyth, C.J.; Waddick, K.G.; Ahlgren, L.T.; Chelstrom, L.M.; Burkhardt, A.; Bolen, J.; Myers, D.E. Biotherapy of B-cell precursor leukemia by targeting genistein to CD19-associated tyrosine kinases. Science 1995, 267, 886–891. [Google Scholar]
- Rusin, A.; Krawczyk, Z.; Grynkiewicz, G.; Gogler, A.; Zawisza-Puchałka, J.; Szeja, W. Synthetic derivatives of genistein, their properties and possible applications. Acta Biochim. Pol 2010, 57, 23–34. [Google Scholar]
- Del Valle, E.M.M. Cyclodextrins and their uses: A review. Process Biochem 2004, 39, 1033–1046. [Google Scholar]
- Faustino, C.M.C.; Calado, A.R.T.; Garcia-Rio, L. Interactions between β-cyclodextrin and an amino acid-based anionic gemini surfactant derived from cysteine. J. Colloid Interface Sci 2012, 367, 286–292. [Google Scholar]
- Loftsson, T.; Duchêne, D. Cyclodextrins and their pharmaceutical applications. Int. J. Pharm 2007, 329, 1–11. [Google Scholar]
- Szejtli, J. Medicinal applications of cyclodextrins. Med. Res. Rev 1994, 14, 353–386. [Google Scholar]
- Daruházi, A.E.; Szente, L.; Balogh, B.; Mátyus, P.; Béni, S.; Takács, M.; Gergely, A.; Horváth, P.; Szoke, E.; Lemberkovics, E. Utility of cyclodextrins in the formulation of genistein part 1. Preparation and physicochemical properties of genistein complexes with native cyclodextrins. J. Pharm. Biomed. Anal 2008, 48, 636–640. [Google Scholar]
- Lee, S.-H.; Kim, Y.H.; Yu, H.-J.; Cho, N.-S.; Kim, T.-H.; Kim, D.C.; Chung, C.B.; Hwang, Y.I.; Kim, K.H. Enhanced bioavailability of soy isoflavones by complexation with beta-cyclodextrin in rats. Biosci. Biotechnol. Biochem 2007, 71, 2927–2933. [Google Scholar]
- Boys, S.F.; Bernardi, F. Calculation of small molecular interactions by differences of separate total energies—Some procedures with reduced errors. Mol. Phys 1970, 19, 553–566. [Google Scholar]
- Dua, K.; Pabreja, K.; Ramana, M.V.; Lather, V. Dissolution behavior of β-cyclodextrin molecular inclusion complexes of aceclofenac. J. Pharm. Bioallied Sci 2011, 3, 417–425. [Google Scholar]
- Ficarra, R.; Ficarra, P.; di Bella, M.R.; Raneri, D.; Tommasini, S.; Calabrò, M.L.; Gamberini, M.C.; Rustichelli, C. Study of beta-blockers/beta-cyclodextrins inclusion complex by NMR, DSC, X-ray and SEM investigation. J. Pharm. Biomed. Anal 2000, 23, 33–40. [Google Scholar]
- Pralhad, T.; Rajendrakumar, K. Study of freeze-dried quercetin-cyclodextrin binary systems by DSC, FT-IR, X-ray diffraction and SEM analysis. J. Pharm. Biomed. Anal 2004, 34, 333–339. [Google Scholar]
- Liu, Y.; Chen, G.-S.; Li, L.; Zhang, H.-Y.; Cao, D.-X.; Yuan, Y.-J. Inclusion complexation and solubilization of paclitaxel by bridged bis(β-cyclodextrin)s containing a tetraethylenepentaamino spacer. J. Med. Chem 2003, 46, 4634–4637. [Google Scholar]
- Shim, H.-Y.; Park, J.-H.; Paik, H.-D.; Nah, S.-Y.; Kim, D.S.; Han, Y.S. Genistein-induced apoptosis of human breast cancer MCF-7 cells involves calpain-caspase and apoptosis signaling kinase 1-p38 mitogen-activated protein kinase activation cascades. Anticancer Drugs 2007, 18, 649–657. [Google Scholar]
- Singletary, K.W.; Frey, R.S.; Li, J.Y. Differential effects of genistein on cell proliferation, cyclin B1, and p34cdc2 in transformed and nontransformed human breast cells. Pharm. Biol 2002, 40, 35–42. [Google Scholar]
- Solomon, L.A.; Ali, S.; Banerjee, S.; Munkarah, A.R.; Morris, R.T.; Sarkar, F.H. Sensitization of ovarian cancer cells to cisplatin by genistein: The role of NF-kappaB. J. Ovarian Res 2008, 1, 9. [Google Scholar]
- Gossner, G.; Choi, M.; Tan, L.; Fogoros, S.; Griffith, K.A.; Kuenker, M.; Liu, J.R. Genistein-induced apoptosis and autophagocytosis in ovarian cancer cells. Gynecol. Oncol 2007, 105, 23–30. [Google Scholar]
- Zhang, B.; Liu, J.Y.; Pan, J.S.; Han, S.P.; Yin, X.X.; Wang, B.; Hu, G. Combined treatment of ionizing radiation with genistein on cervical cancer HeLa cells. J. Pharmacol. Sci 2006, 102, 129–135. [Google Scholar]
- Sahin, K.; Tuzcu, M.; Basak, N.; Caglayan, B.; Kilic, U.; Sahin, F.; Kucuk, O. Sensitization of cervical cancer cells to cisplatin by genistein: The role of nfb and akt/mtor signaling pathways. J. Oncol 2012, 2012. [Google Scholar] [CrossRef]
- Hipler, U.C.; Schönfelder, U.; Hipler, C.; Elsner, P. Influence of cyclodextrins on the proliferation of HaCaT keratinocytes in vitro. J. Biomed. Mat. Res. Part A 2007, 83, 70–79. [Google Scholar]
- Yadav, V.R.; Prasad, S.; Kannappan, R.; Ravindran, J.; Chaturvedi, M.M.; Vaahtera, L.; Parkkinen, J.; Aggarwal, B.B. Cyclodextrin-complexed curcumin exhibits anti-inflammatory and antiproliferative activities superior to those of curcumin through higher cellular uptake. Biochem. Pharmacol 2010, 80, 1021–1032. [Google Scholar]
- Mendonça, E.A.M.; Lira, M.C.B.; Rabello, M.M.; Cavalcanti, I.M.F.; Galdino, S.L.; Pitta, I.R.; Lima Mdo, C.; Pitta, M.G.; Hernandes, M.Z.; Santos-Magalhães, N.S. Enhanced antiproliferative activity of the new anticancer candidate LPSF/AC04 in cyclodextrin inclusion complexes encapsulated into liposomes. AAPS 2012, 13, 1355–1366. [Google Scholar]
- Pourgholami, M.H.; Wangoo, K.T.; Morris, D.L. Albendazole-cyclodextrin complex: Enhanced cytotoxicity in ovarian cancer cells. Anticancer Res 2008, 28, 2775–2779. [Google Scholar]
- Li, W.; Chen, L.; Huang, Z.; Wu, X.; Zhang, Y.; Hu, Q.; Wang, Y. The influence of cyclodextrin modification on cellular uptake and transfection efficiency of polyplexes. Org. Biomol. Chem 2011, 9, 7799–7806. [Google Scholar]
- Dhule, S.S.; Penfornis, P.; Frazier, T.; Walker, R.; Feldman, J.; Tan, G.; He, J.; Alb, A.; John, V.; Pochampally, R. Curcumin-loaded γ-cyclodextrin liposomal nanoparticles as delivery vehicles for osteosarcoma. Nanomedicine 2012, 8, 440–451. [Google Scholar]
- Arun, R.; Ashok Kumar, C.K.; Sravanthi, V.V.N.S.S. Cyclodextrins as drug carrier molecule: A review. Sci. Pharm 2008, 76, 567–598. [Google Scholar]
- Weber, P.; Wagner, M.; Schneckenburger, H. Cholesterol dependent uptake and interaction of doxorubicin in MCF-7 breast cancer cells. Int. J. Mol. Sci 2013, 14, 8358–8366. [Google Scholar]
- Ulanowska, K.; Majchrzyk, A.; Moskot, M.; Jakóbkiewicz-Banecka, J.; Węgrzyn, G. Assessment of antibacterial effects of flavonoids by estimation of generation times in liquid bacterial cultures. Biologia 2007, 62, 132–135. [Google Scholar]
- Hong, H.; Landauer, M.R.; Foriska, M.A.; Ledney, G.D. Antibacterial activity of the soy isoflavone genistein. J. Basic Microbiol 2006, 46, 329–335. [Google Scholar]
- Zaki, M.; Zaid, A.; Magd, M.; AbouZid, S. Antibacterial effects of isoflavones isolated from Cicer arietinum. Int. J. Nat. Prod. Res 2013, 2, 1–5. [Google Scholar]
- Verdrengh, M.; Collins, L.V.; Bergin, P.; Tarkowski, A. Phytoestrogen genistein as an anti-staphylococcal agent. Microbes Infect 2004, 6, 86–92. [Google Scholar]
- Sun, H-K.; Seshadri, M.; Lingard, S.; Monaghan, W.; Faoagali, J.; Chan, E.; McDonald, H.; Houston, T.; King, M.; Peak, I.; et al. Antibacterial activity of beta-cyclodextrin and 2-hydroxypropyl-beta-cyclodextrin trimethoprim complexes. Am. J. Microbiol 2011, 2, 1–8. [Google Scholar]
- Denadai, A.M.; Teixeira, K.I.; Santoro, M.M.; Pimenta, A.M.; Cortés, M.E.; Sinisterra, R.D. Supramolecular self-assembly of β-cyclodextrin: An effective carrier of the antimicrobial agent chlorhexidine. Carbohydr. Res 2007, 342, 2286–2296. [Google Scholar]
- Wang, J.; Cai, Z. Incorporation of the antibacterial agent, miconazole nitrate into a cellulosic fabric grafted with β-cyclodextrin. Carbohydr. Polym 2008, 72, 695–700. [Google Scholar]
- Farina, H.G.; Pomies, M.; Alonso, D.F.; Gomez, D.E. Antitumor and antiangiogenic activity of soy isoflavone genistein in mouse models of melanoma and breast cancer. Oncol. Rep 2006, 16, 885–891. [Google Scholar]
- Kiriakidis, S.; Högemeier, O.; Starcke, S.; Dombrowski, F.; Hahne, J.C.; Pepper, M.; Jha, H.C.; Wernert, N. Novel tempeh (fermented soyabean) isoflavones inhibit in vivo angiogenesis in the chicken chorioallantoic membrane assay. Br. J. Nutr 2005, 93, 317–323. [Google Scholar]
- Banerjee, S.; Li, Y.; Wang, Z.; Sarkar, F.H. Multi-targeted therapy of cancer by genistein. Cancer Lett 2008, 269, 226–242. [Google Scholar]
- Dehelean, C.A.; Soica, C.; Peev, C.; Ciurlea, S.; Feflea, S.; Kasa, P., Jr. A pharmaco-toxicological evaluation of betulinic acid mixed with hydroxipropilgamma cyclodextrin on in vitro and in vivo Models. Farmacia 2011, 59, 51–59. [Google Scholar]
- Gaussian 09, Revision A.02; Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Mennucci, B.; Petersson, G.A.; et al. (Eds.) Gaussian Inc: Wallingford, CT, USA, 2009.
- Grimme, S. Semiempirical GGA-type density functional constructed with a long-range dispersion correction. J. Comput. Chem 2006, 27, 1787–1799. [Google Scholar]
- Cances, E.; Mennucci, B.; Tomasi, J. A new integral equation formalism for the polarizable continuum model: Theoretical background and applications to isotropic and anisotropic dielectrics. J. Chem. Phys 1997, 107, 3032–3043. [Google Scholar]
- Cossi, M.; Barone, V.; Mennucci, B.; Tomasi, J. Ab initio study of ionic solutions by a polarizable continuum dielectric model. Chem. Phys. Lett 1998, 286, 253–260. [Google Scholar]
- Cossi, M.; Scalmani, G.; Rega, N.; Barone, V. New developments in the polarizable continuum model for quantum mechanical and classical calculations on molecules in solution. J. Chem. Phys 2002, 117, 43–54. [Google Scholar]
- Phase-Solubility Techniques; Higuchi, T.; Connors, A. (Eds.) Wiley Interscience: New York, NY, USA, 1965; Volume 4, pp. 117–211.
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar]
- Ribatti, D.; Vacca, A.; Roncali, L.; Dammacco, F. The chick embryo chorioallantoic membrane as a model for in vivo research on angiogenesis. Int. J. Dev. Biol 1996, 40, 1189–1197. [Google Scholar]
- Dehelean, C.A.; Feflea, S.; Gheorgheosu, D.; Ganta, S.; Cimpean, A.M.; Muntean, D.; Amiji, M.M. Anti-angiogenic and anti-cancer evaluation of betulin nanoemulsion in chicken chorioallantoic membrane and skin carcinoma in Balb/c mice. J. Biomed. Nanotechnol 2013, 9, 577–589. [Google Scholar]
- Ribatti, D. The Chick Embryo Chorioallantoic Membrane in the Study of Angiogenesis and Metastasis; Springer Science + Business Media B.V.: Dordrecht, The Netherlands, 2010. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Solvent | Gen-RAMEB | Gen-HPBCD | Gen-HPGCD | |||
|---|---|---|---|---|---|---|
| Eint (Non-CP a) (kcal/mol) | Eint (CP b) (kcal/mol) | Eint (Non-CP a) (kcal/mol) | Eint (CP b) (kcal/mol) | Eint (Non-CP a) (kcal/mol) | Eint (CP b) (kcal/mol) | |
| Gas Phase | −34.68 | −21.32 | −50.14 | −33.15 | −54.25 | −37.08 |
| DMSO | −30.07 | −19.55 | −43.84 | −31.01 | −48.38 | −36.27 |
| Water | −29.83 | −20.29 | −43.79 | −30.98 | −48.02 | −36.18 |
| Cyclodextrin | Stability constant (M−1) |
|---|---|
| RAMEB | 10.850 |
| HPBCD | 10.900 |
| HPGCD | 12.700 |
| Compound | IC50 values (μM) a | |||
|---|---|---|---|---|
| HeLa | A2780 | MCF7 | A431 | |
| Gen | 25.90 | 20.86 | 50.08 | 72.79 |
| Gen-RAMEB (1:1) | 21.07 | 22.80 | 59.47 | >90 |
| Gen-HPBCD (1:1) | 16.47 | 17.49 | 44.02 | 69.88 |
| Gen-HPGCD (1:1) | 19.09 | 19.84 | 50.40 | 74.35 |
| Compound | Zone of inhibition (mm). Results are presented as mean values ± SD | ||||||
|---|---|---|---|---|---|---|---|
| B. subtilis | S. aureus | E. faecalis | E. coli | S. typhimurium | S. sonnei | P. aeruginosa | |
| Gen | 15 ± 0.46 | 9 ± 0.12 | 7 ± 0.23 | 6 ± 0.23 | 6 ± 0.15 | 6 ± 0.23 | 6 ± 0.12 |
| Gen-RAMEB (1:1) | 16 ± 0.32 | 9 ± 0.17 | 8 ± 0.19 | 6 ± 0.14 | 6 ± 0.13 | 6 ± 0.09 | 6 ± 0.17 |
| Gen-HPBCD (1:1) | 16 ± 0.20 | 9 ± 0.11 | 8 ± 0.13 | 6 ± 0.21 | 6 ± 0.19 | 6 ± 0.14 | 6 ± 0.22 |
| Gen-HPGCD (1:1) | 17 ± 0.28 | 9 ± 0.09 | 8 ± 0.11 | 6 ± 0.16 | 6 ± 0.17 | 6 ± 0.15 | 6 ± 0.13 |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Danciu, C.; Soica, C.; Oltean, M.; Avram, S.; Borcan, F.; Csanyi, E.; Ambrus, R.; Zupko, I.; Muntean, D.; Dehelean, C.A.; et al. Genistein in 1:1 Inclusion Complexes with Ramified Cyclodextrins: Theoretical, Physicochemical and Biological Evaluation. Int. J. Mol. Sci. 2014, 15, 1962-1982. https://doi.org/10.3390/ijms15021962
Danciu C, Soica C, Oltean M, Avram S, Borcan F, Csanyi E, Ambrus R, Zupko I, Muntean D, Dehelean CA, et al. Genistein in 1:1 Inclusion Complexes with Ramified Cyclodextrins: Theoretical, Physicochemical and Biological Evaluation. International Journal of Molecular Sciences. 2014; 15(2):1962-1982. https://doi.org/10.3390/ijms15021962
Chicago/Turabian StyleDanciu, Corina, Codruta Soica, Mircea Oltean, Stefana Avram, Florin Borcan, Erzsebet Csanyi, Rita Ambrus, Istvan Zupko, Delia Muntean, Cristina A. Dehelean, and et al. 2014. "Genistein in 1:1 Inclusion Complexes with Ramified Cyclodextrins: Theoretical, Physicochemical and Biological Evaluation" International Journal of Molecular Sciences 15, no. 2: 1962-1982. https://doi.org/10.3390/ijms15021962
APA StyleDanciu, C., Soica, C., Oltean, M., Avram, S., Borcan, F., Csanyi, E., Ambrus, R., Zupko, I., Muntean, D., Dehelean, C. A., Craina, M., & Popovici, R. A. (2014). Genistein in 1:1 Inclusion Complexes with Ramified Cyclodextrins: Theoretical, Physicochemical and Biological Evaluation. International Journal of Molecular Sciences, 15(2), 1962-1982. https://doi.org/10.3390/ijms15021962