Syk/JNK/AP-1 Signaling Pathway Mediates Interleukin-6-Promoted Cell Migration in Oral Squamous Cell Carcinoma

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
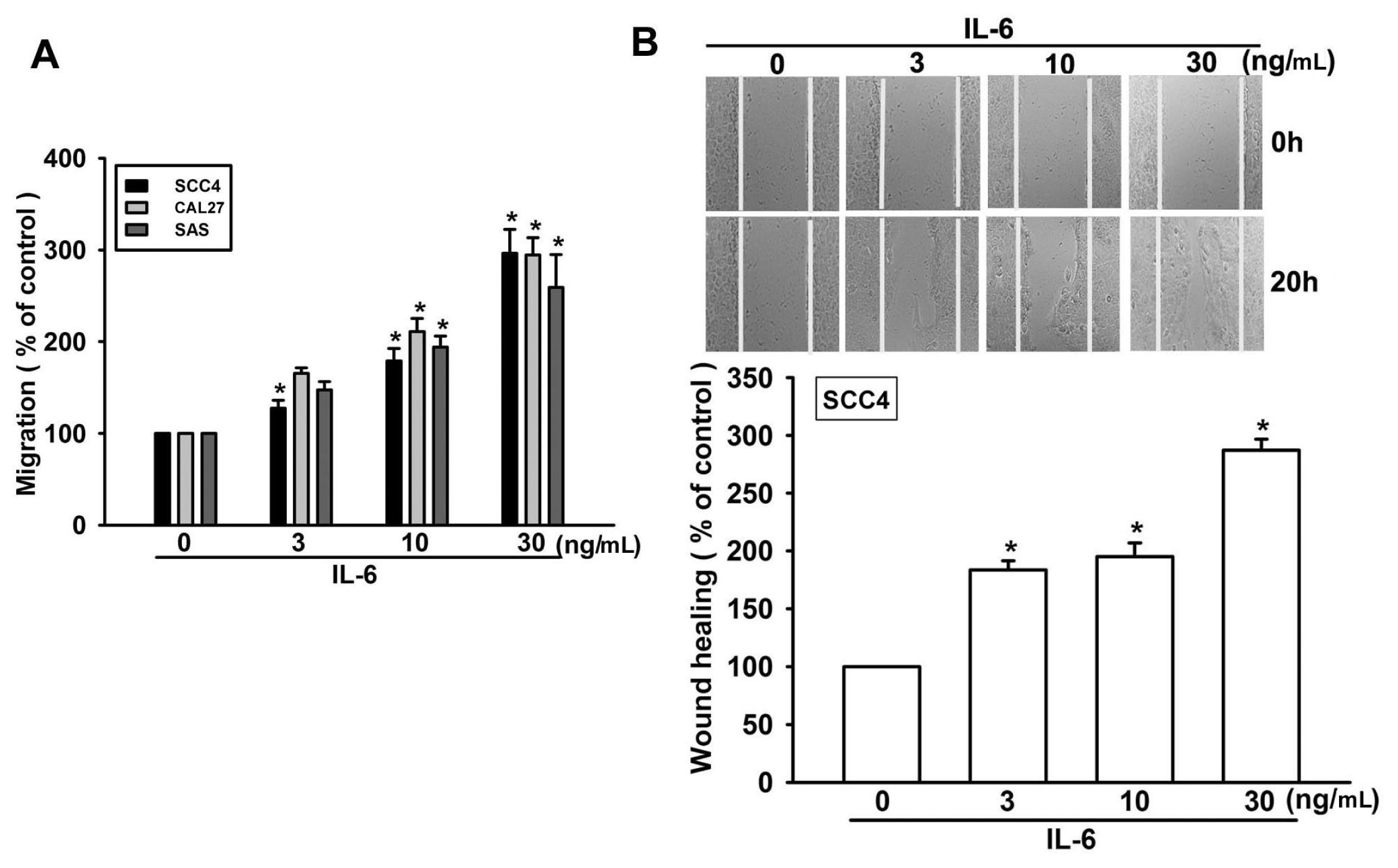
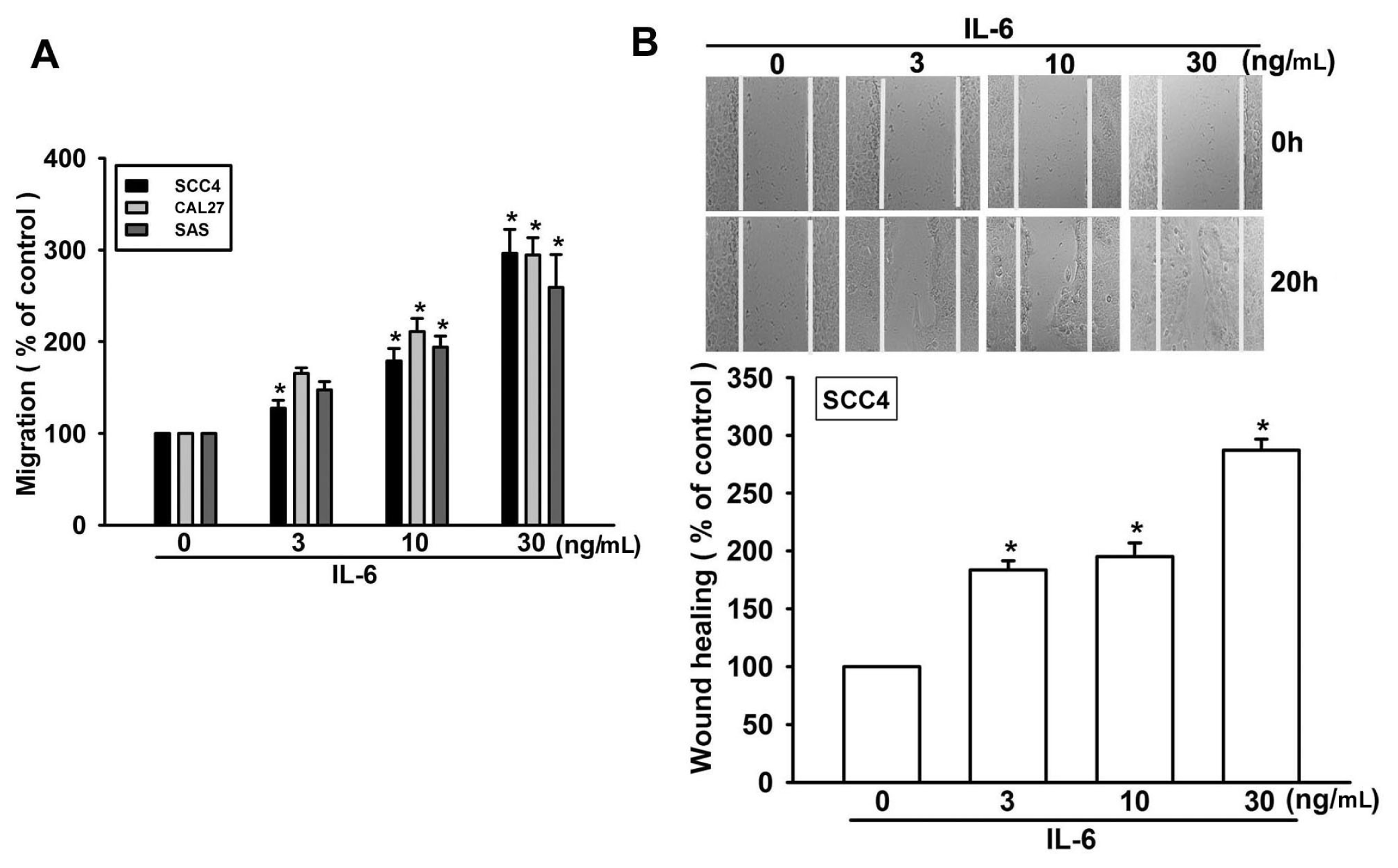
2.1. IL-6 Increases Migration in OSCC (Oral Squamous Cell Carcinoma) Cells
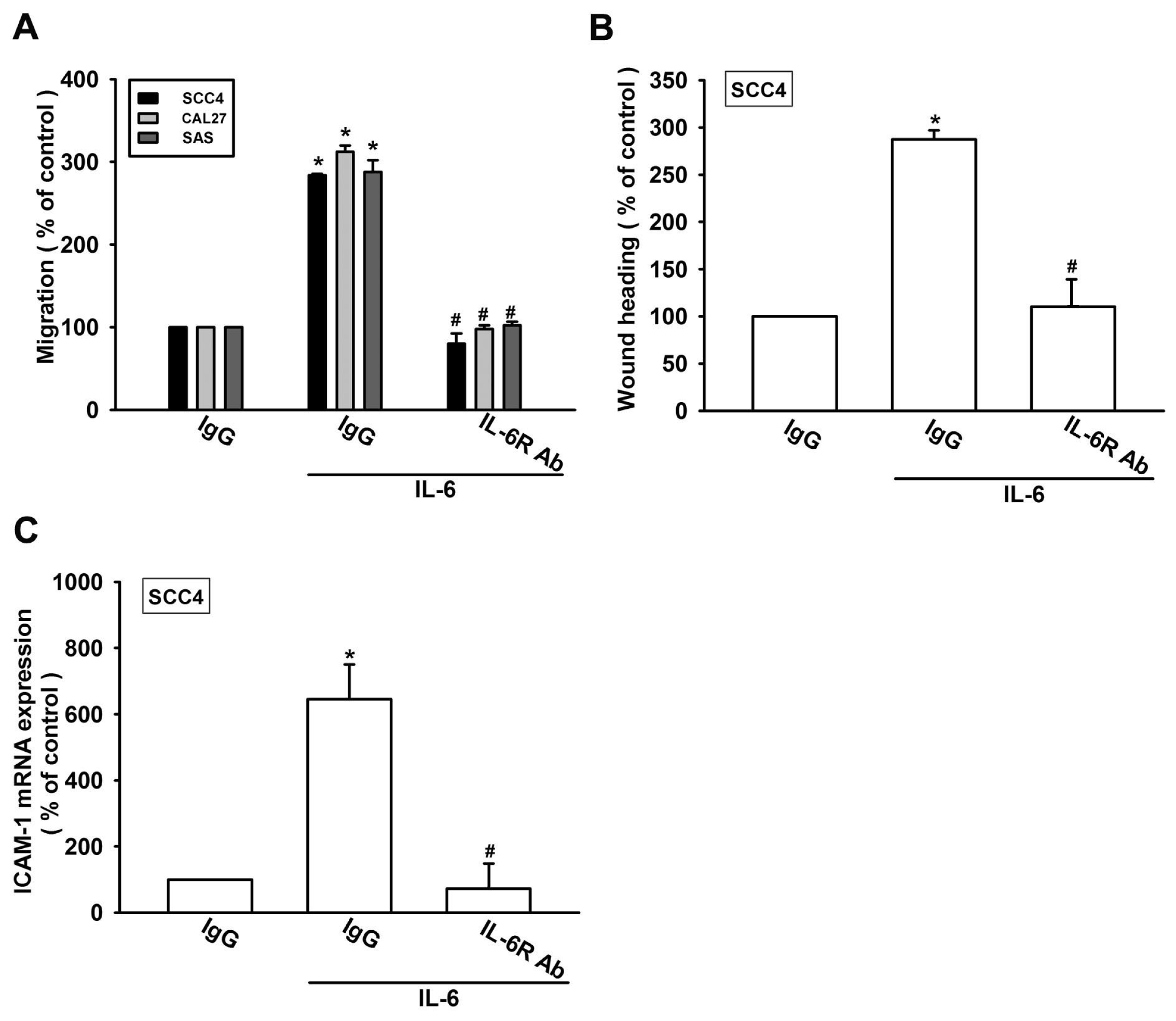
2.2. Involvement of ICAM-1 in IL-6-Directed Cell Migration of OSCC Cells
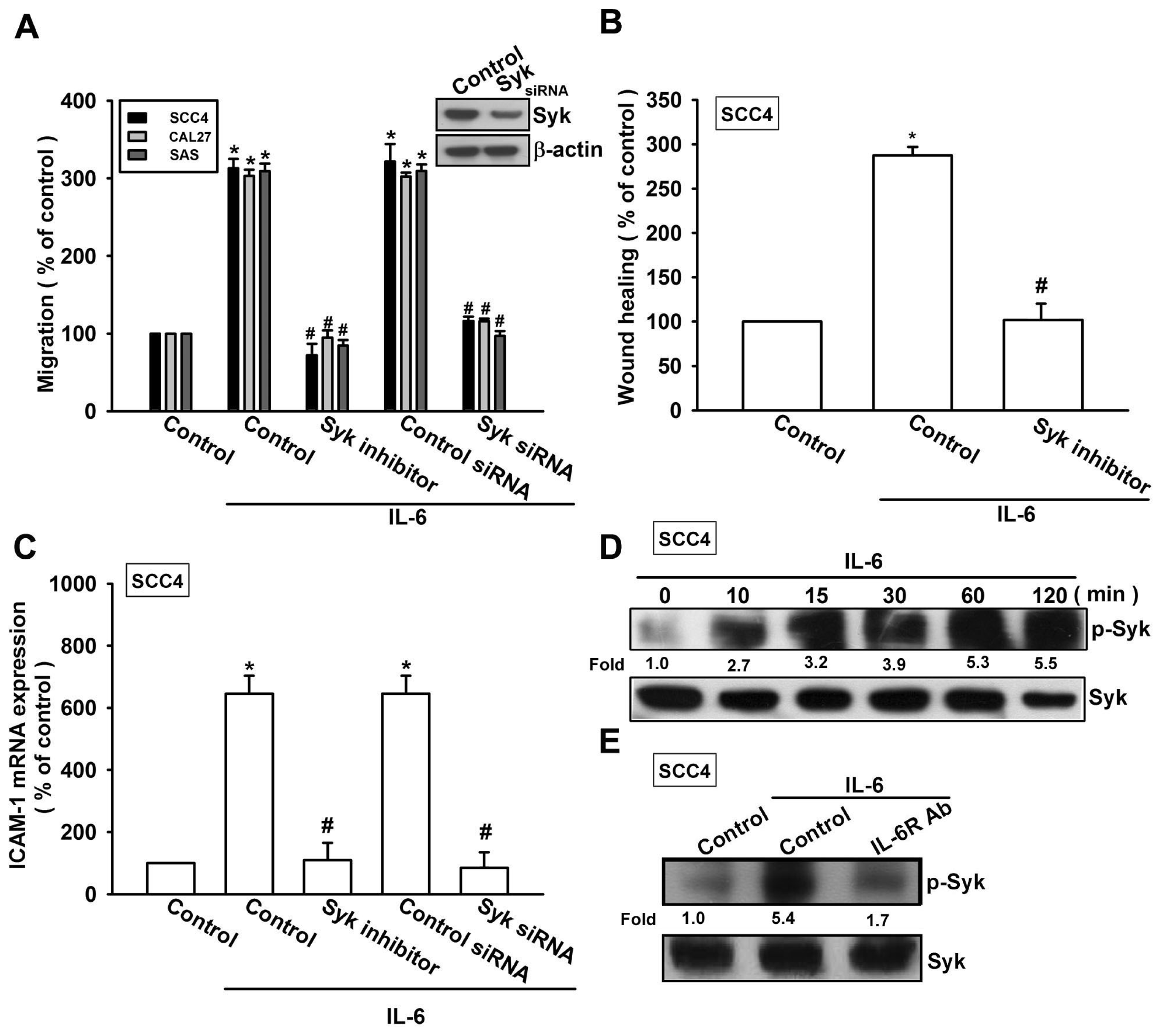
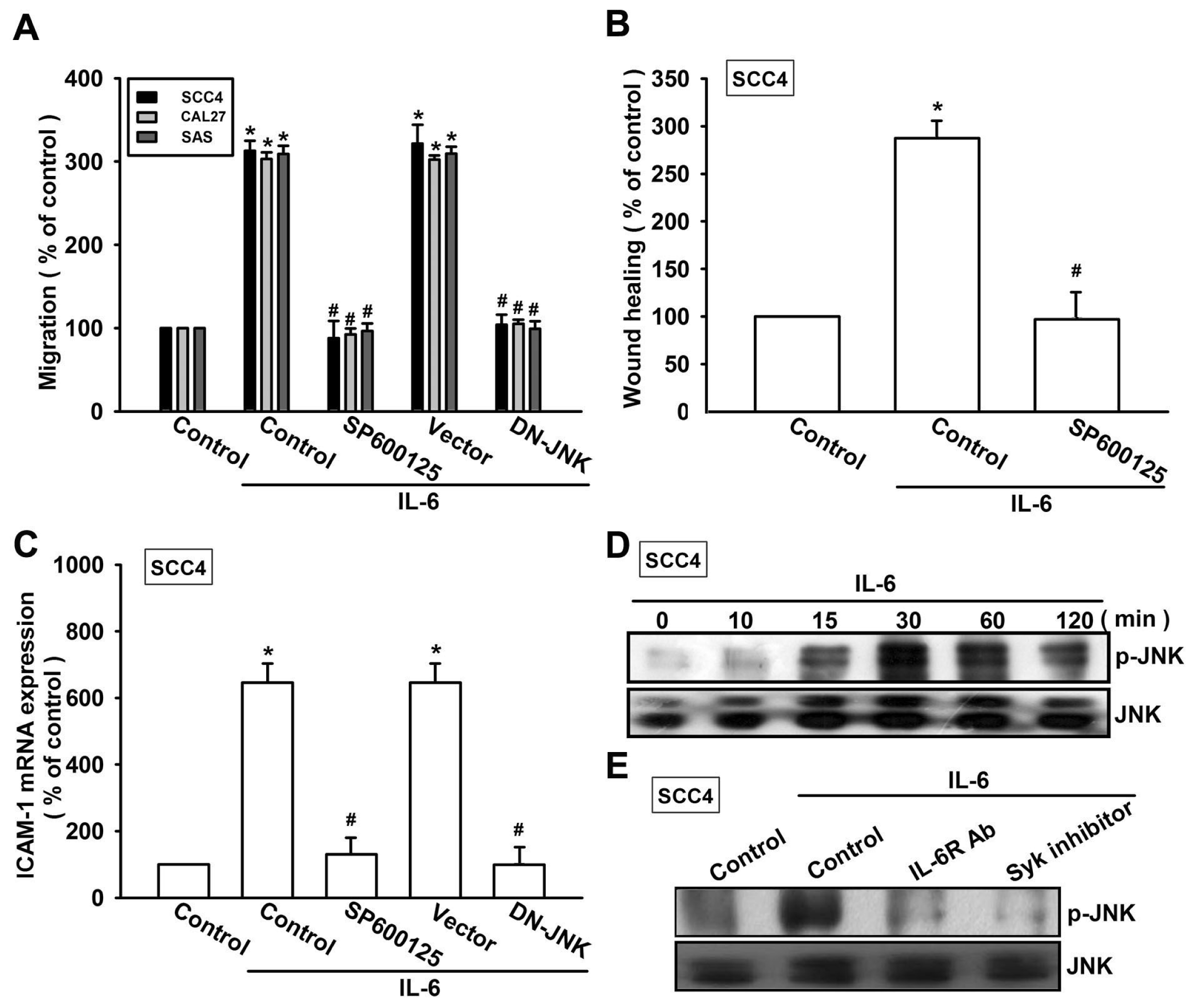
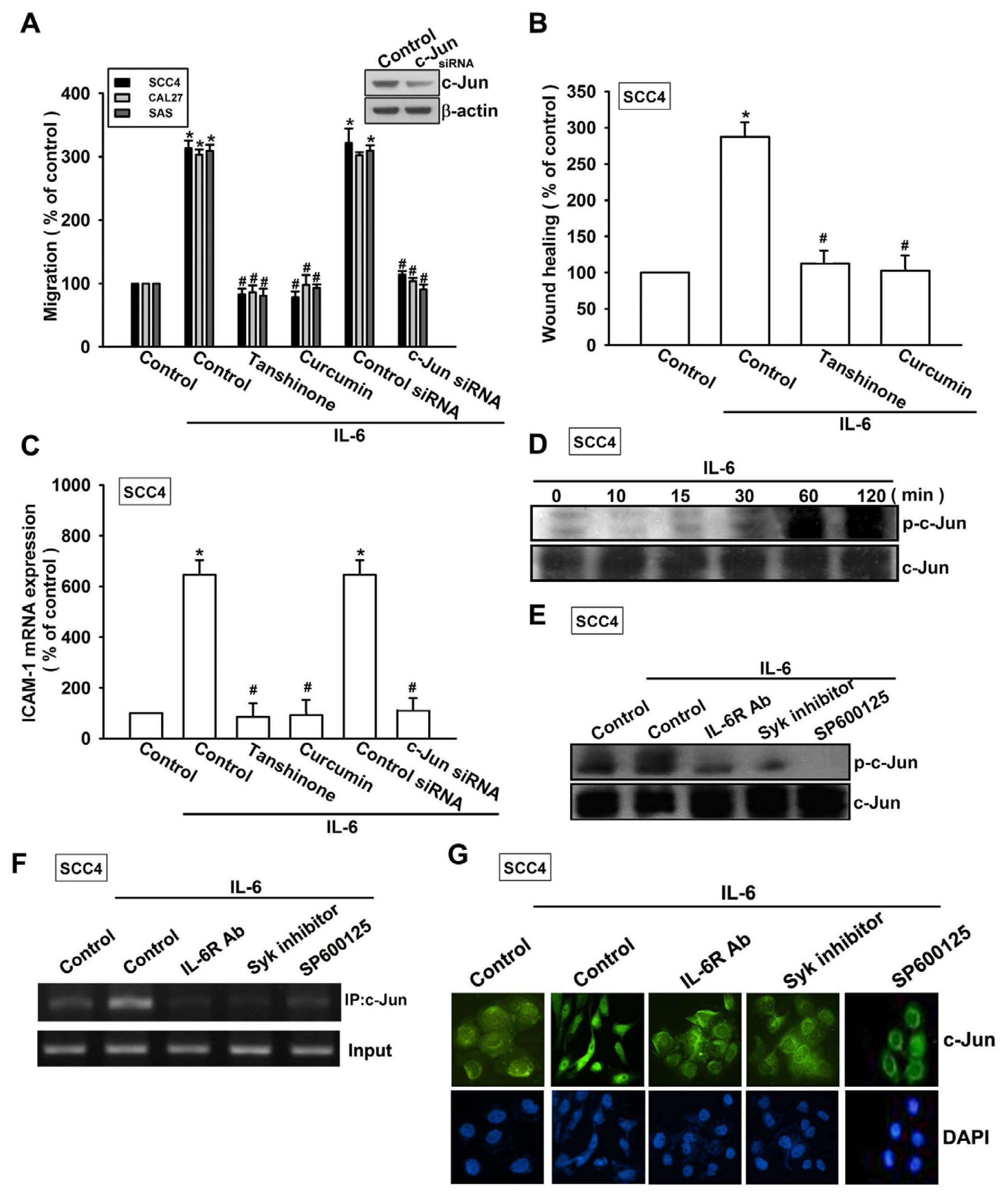
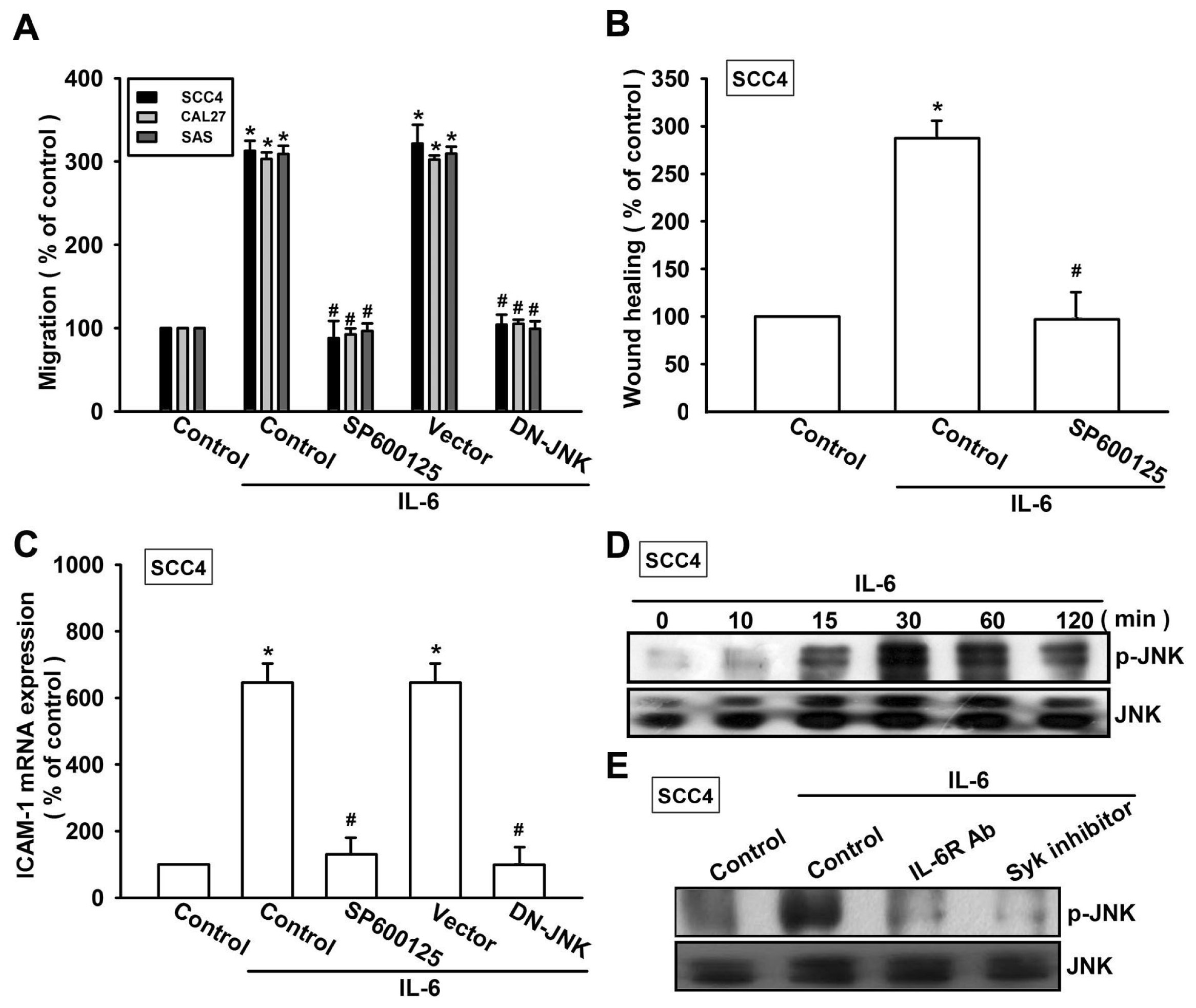
2.3. Syk and c-Jun N-Terminal Kinase (JNK) Signaling Pathways are Involved in IL-6-Mediated ICAM-1 Upregulation and Cell Motility of OSCC Cells
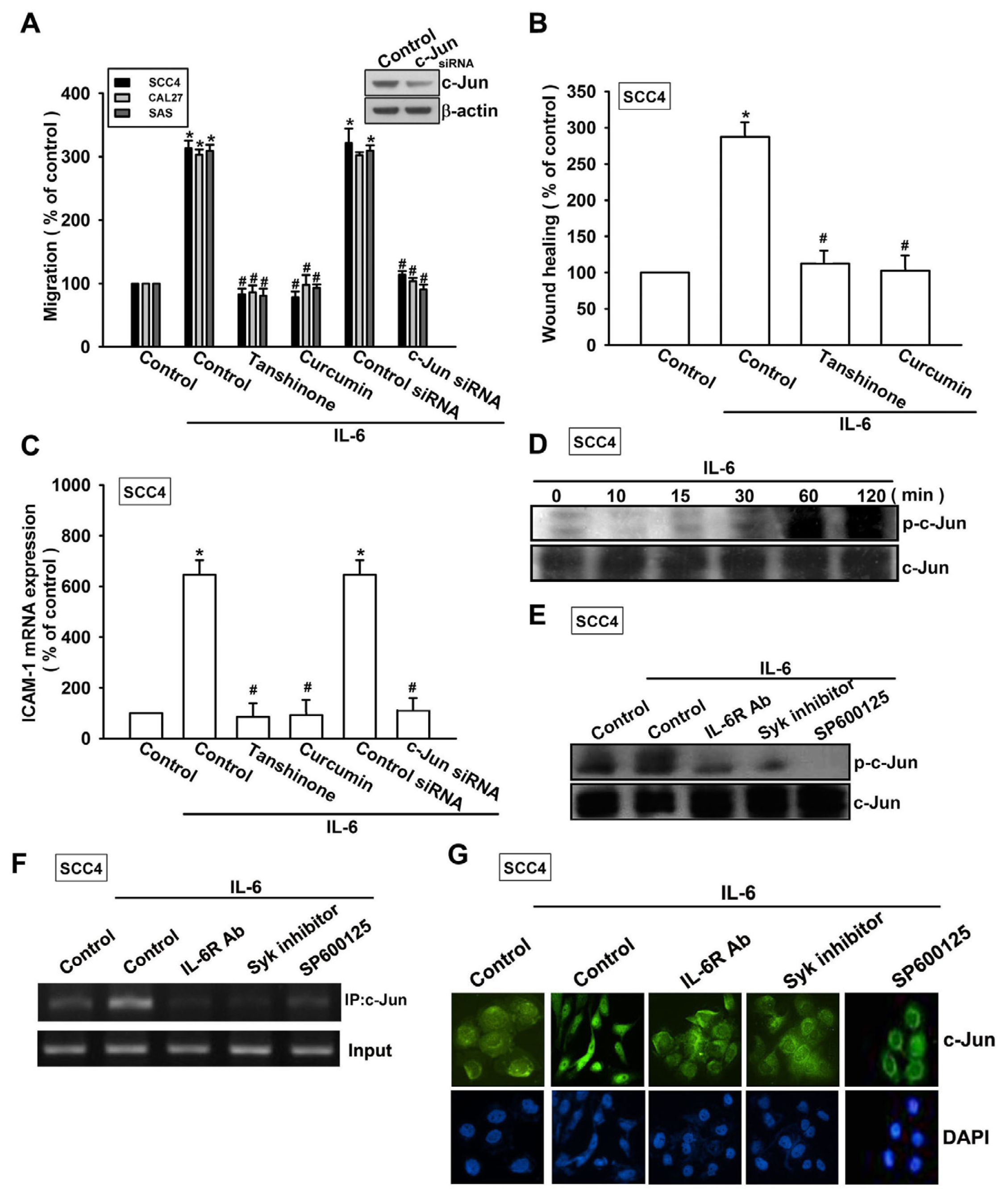
2.4. Involvement of AP-1 in IL-6-Induced Cell Migration and ICAM-1 Expression
3. Discussion
4. Experimental Section
4.1. Materials
4.2. Cell Culture
4.3. Migration Assay
4.4. Wound-Healing Migration Assay
4.5. Quantitative Real-Time Polymerase Chain Reaction (PCR)
4.6. Western Blot Analysis
4.7. Transfection of Small Interfering RNAs (siRNAs) or Mutant
4.8. Flow Cytometric Analysis
4.9. Immunofluorocytochemistry
4.10. Chromatin Immunoprecipitation (ChIP)
4.11. Statistical Analysis
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Lyons, A.J.; Jones, J. Cell adhesion molecules, the extracellular matrix and oral squamous carcinoma. Int. J. Oral Maxillofac. Surg 2007, 36, 671–679. [Google Scholar]
- Bau, D.-T.; Lin, C.-C.; Chiu, C.-F.; Tsai, M.-H. Role of nonhomologous end-joining in oral cancer and personalized pharmacogenomics. BioMedicine 2012, 2, 41–47. [Google Scholar]
- Greenberg, J.S.; El Naggar, A.K.; Mo, V.; Roberts, D.; Myers, J.N. Disparity in pathologic and clinical lymph node staging in oral tongue carcinoma. Cancer 2003, 98, 508–515. [Google Scholar]
- Thomas, G.J.; Speight, P.M. Cell adhesion molecules and oral cancer. Crit. Rev. Oral Biol. Med 2001, 12, 479–498. [Google Scholar]
- Rose-John, S.; Scheller, J.; Elson, G.; Jones, S.A. Interleukin-6 biology is coordinated by membrane-bound and soluble receptors: Role in inflammation and cancer. J. Leukoc. Biol 2006, 80, 227–236. [Google Scholar]
- Okamoto, M.; Lee, C.; Oyasu, R. Interleukin-6 as a paracrine and autocrine growth factor in human prostatic carcinoma cells in vitro. Cancer Res. 1997, 57, 141–146. [Google Scholar]
- Okamoto, M.; Hattori, K.; Oyasu, R. Interleukin-6 functions as an autocrine growth factor in human bladder carcinoma cell lines in vitro. Int. J. Cancer 1997, 72, 149–154. [Google Scholar]
- Miki, S.; Iwano, M.; Miki, Y.; Yamamoto, M.; Tang, B.; Yokokawa, K.; Sonoda, T.; Hirano, T.; Kishimoto, T. Interleukin-6 (IL-6) functions as an in vitro autocrine growth factor in renal cell carcinomas. FEBS Lett 1989, 250, 607–610. [Google Scholar]
- Chen, L.; Mory, Y.; Zilberstein, A.; Revel, M. Growth inhibition of human breast carcinoma and leukemia/lymphoma cell lines by recombinant interferon-beta 2. Proc. Natl. Acad. Sci. USA 1988, 85, 8037–8041. [Google Scholar]
- Okamoto, M.; Lee, C.; Oyasu, R. Autocrine effect of androgen on proliferation of an androgen responsive prostatic carcinoma cell line, LNCAP: Role of interleukin-6. Endocrinology 1997, 138, 5071–5074. [Google Scholar]
- Eustace, D.; Han, X.; Gooding, R.; Rowbottom, A.; Riches, P.; Heyderman, E. Interleukin-6 (IL-6) functions as an autocrine growth factor in cervical carcinomas in vitro. Gynecol. Oncol. 1993, 50, 15–19. [Google Scholar]
- Lu, C.; Kerbel, R.S. Interleukin-6 undergoes transition from paracrine growth inhibitor to autocrine stimulator during human melanoma progression. J. Cell Biol 1993, 120, 1281–1288. [Google Scholar]
- Lin, Y.M.; Chang, Z.L.; Liao, Y.Y.; Chou, M.C.; Tang, C.H. IL-6 promotes ICAM-1 expression and cell motility in human osteosarcoma. Cancer Lett 2013, 328, 135–143. [Google Scholar]
- Tang, C.H.; Chen, C.F.; Chen, W.M.; Fong, Y.C. IL-6 increases MMP-13 expression and motility in human chondrosarcoma cells. J. Biol. Chem 2011, 286, 11056–11066. [Google Scholar]
- Gupta, G.P.; Massague, J. Cancer metastasis: Building a framework. Cell 2006, 127, 679–695. [Google Scholar]
- Tang, C.H. Molecular mechanisms of chondrosarcoma metastasis. BioMedicine 2012, 2, 92–98. [Google Scholar]
- Desgrosellier, J.S.; Cheresh, D.A. Integrins in cancer: Biological implications and therapeutic opportunities. Nat. Rev. Cancer 2010, 10, 9–22. [Google Scholar]
- Makrilia, N.; Kollias, A.; Manolopoulos, L.; Syrigos, K. Cell adhesion molecules: Role and clinical significance in cancer. Cancer Investig 2009, 27, 1023–1037. [Google Scholar]
- Lawson, C.; Wolf, S. ICAM-1 signaling in endothelial cells. Pharmacol. Rep 2009, 61, 22–32. [Google Scholar]
- Zimmerman, T.; Blanco, F.J. Inhibitors targeting the LFA-1/ICAM-1 cell-adhesion interaction: Design and mechanism of action. Curr. Pharm. Des 2008, 14, 2128–2139. [Google Scholar]
- Duperray, A.; Languino, L.R.; Plescia, J.; McDowall, A.; Hogg, N.; Craig, A.G.; Berendt, A.R.; Altieri, D.C. Molecular identification of a novel fibrinogen binding site on the first domain of ICAM-1 regulating leukocyte-endothelium bridging. J. Biol. Chem 1997, 272, 435–441. [Google Scholar]
- Huang, W.C.; Chan, S.T.; Yang, T.L.; Tzeng, C.C.; Chen, C.C. Inhibition of ICAM-1 gene expression, monocyte adhesion and cancer cell invasion by targeting IKK complex: Molecular and functional study of novel alpha-methylene-gamma-butyrolactone derivatives. Carcinogenesis 2004, 25, 1925–1934. [Google Scholar]
- Rosette, C.; Roth, R.B.; Oeth, P.; Braun, A.; Kammerer, S.; Ekblom, J.; Denissenko, M.F. Role of ICAM1 in invasion of human breast cancer cells. Carcinogenesis 2005, 26, 943–950. [Google Scholar]
- Yang, S.F.; Chen, M.K.; Hsieh, Y.S.; Chung, T.T.; Hsieh, Y.H.; Lin, C.W.; Su, J.L.; Tsai, M.H.; Tang, C.H. Prostaglandin E2/EP1 signaling pathway enhances intercellular adhesion molecule 1 (ICAM-1) expression and cell motility in oral cancer cells. J. Biol. Chem 2010, 285, 29808–29016. [Google Scholar]
- Chuang, J.Y.; Chang, A.C.; Chiang, I.P.; Tsai, M.H.; Tang, C.H. Apoptosis signal-regulating kinase 1 is involved in WISP-1-promoted cell motility in human oral squamous cell carcinoma cells. PLoS One 2013, 8, e78022. [Google Scholar]
- Lin, C.W.; Chuang, C.Y.; Tang, C.H.; Chang, J.L.; Lee, L.M.; Lee, W.J.; Chow, J.M.; Yang, S.F.; Chien, M.H. Combined effects of icam-1 single-nucleotide polymorphisms and environmental carcinogens on oral cancer susceptibility and clinicopathologic development. PLoS One 2013, 8, e72940. [Google Scholar]
- Lo, C.W.; Chen, M.W.; Hsiao, M.; Wang, S.; Chen, C.A.; Hsiao, S.M.; Chang, J.S.; Lai, T.C.; Rose-John, S.; Kuo, M.L.; et al. IL-6 trans-signaling in formation and progression of malignant ascites in ovarian cancer. Cancer Res 2011, 71, 424–434. [Google Scholar]
- Liu, J.F.; Hou, S.M.; Tsai, C.H.; Huang, C.Y.; Hsu, C.J.; Tang, C.H. CCN4 induces vascular cell adhesion molecule-1 expression in human synovial fibroblasts and promotes monocyte adhesion. Biochim. Biophys. Acta 2013, 1833, 966–975. [Google Scholar]
- Chen, Z.; Hagler, J.; Palombella, V.J.; Melandri, F.; Scherer, D.; Ballard, D.; Maniatis, T. Signal-induced site-specific phosphorylation targets I kappa B alpha to the ubiquitin-proteasome pathway. Genes Dev 1995, 9, 1586–1597. [Google Scholar]
- Van de Stolpe, A.; van der Saag, P.T. Intercellular adhesion molecule-1. J. Mol. Med 1996, 74, 13–33. [Google Scholar]
- Chen, L.M.; Kuo, C.H.; Lai, T.Y.; Lin, Y.M.; Su, C.C.; Hsu, H.H.; Tsai, F.J.; Tsai, C.H.; Huang, C.Y.; Tang, C.H. RANKL increases migration of human lung cancer cells through intercellular adhesion molecule-1 up-regulation. J. Cell Biochem 2011, 112, 933–941. [Google Scholar]
- Fong, Y.C.; Lin, C.Y.; Su, Y.C.; Chen, W.C.; Tsai, F.J.; Tsai, C.H.; Huang, C.Y.; Tang, C.H. CCN6 enhances ICAM-1 expression and cell motility in human chondrosarcoma cells. J. Cell. Physiol 2012, 227, 223–232. [Google Scholar]
- Coussens, L.M.; Werb, Z. Inflammation and cancer. Nature 2002, 420, 860–867. [Google Scholar]
- Mantovani, A.; Bottazzi, B.; Colotta, F.; Sozzani, S.; Ruco, L. The origin and function of tumor-associated macrophages. Immunol. Today 1992, 13, 265–270. [Google Scholar]
- Efremov, D.G.; Laurenti, L. The Syk kinase as a therapeutic target in leukemia and lymphoma. Expert Opin. Investig. Drugs 2011, 20, 623–636. [Google Scholar]
- Tsou, H.K.; Chen, H.T.; Hung, Y.H.; Chang, C.H.; Li, T.M.; Fong, Y.C.; Tang, C.H. HGF and c-Met interaction promotes migration in human chondrosarcoma cells. PLoS One 2013, 8, e53974. [Google Scholar]
- Wu, M.H.; Lo, J.F.; Kuo, C.H.; Lin, J.A.; Lin, Y.M.; Chen, L.M.; Tsai, F.J.; Tsai, C.H.; Huang, C.Y.; Tang, C.H. Endothelin-1 promotes MMP-13 production and migration in human chondrosarcoma cells through FAK/PI3K/Akt/mTOR pathways. J. Cell. Physiol 2012, 227, 3016–3026. [Google Scholar]
- Tang, C.H.; Hsu, C.J.; Fong, Y.C. The CCL5/CCR5 axis promotes interleukin-6 production in human synovial fibroblasts. Arthritis Rheum 2010, 62, 3615–3624. [Google Scholar]
- Huang, C.Y.; Chen, S.Y.; Tsai, H.C.; Hsu, H.C.; Tang, C.H. Thrombin induces epidermal growth factor receptor transactivation and CCL2 expression in human osteoblasts. Arthritis Rheum 2012, 64, 3344–3354. [Google Scholar]



© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chuang, J.-Y.; Huang, Y.-L.; Yen, W.-L.; Chiang, I.-P.; Tsai, M.-H.; Tang, C.-H. Syk/JNK/AP-1 Signaling Pathway Mediates Interleukin-6-Promoted Cell Migration in Oral Squamous Cell Carcinoma. Int. J. Mol. Sci. 2014, 15, 545-559. https://doi.org/10.3390/ijms15010545
Chuang J-Y, Huang Y-L, Yen W-L, Chiang I-P, Tsai M-H, Tang C-H. Syk/JNK/AP-1 Signaling Pathway Mediates Interleukin-6-Promoted Cell Migration in Oral Squamous Cell Carcinoma. International Journal of Molecular Sciences. 2014; 15(1):545-559. https://doi.org/10.3390/ijms15010545
Chicago/Turabian StyleChuang, Jing-Yuan, Yuan-Li Huang, Wei-Lin Yen, I-Ping Chiang, Ming-Hsui Tsai, and Chih-Hsin Tang. 2014. "Syk/JNK/AP-1 Signaling Pathway Mediates Interleukin-6-Promoted Cell Migration in Oral Squamous Cell Carcinoma" International Journal of Molecular Sciences 15, no. 1: 545-559. https://doi.org/10.3390/ijms15010545