Upregulation of Phosphorylated HSP27, PRDX2, GRP75, GRP78 and GRP94 in Acquired Middle Ear Cholesteatoma Growth


Abstract
:1. Introduction
2. Results
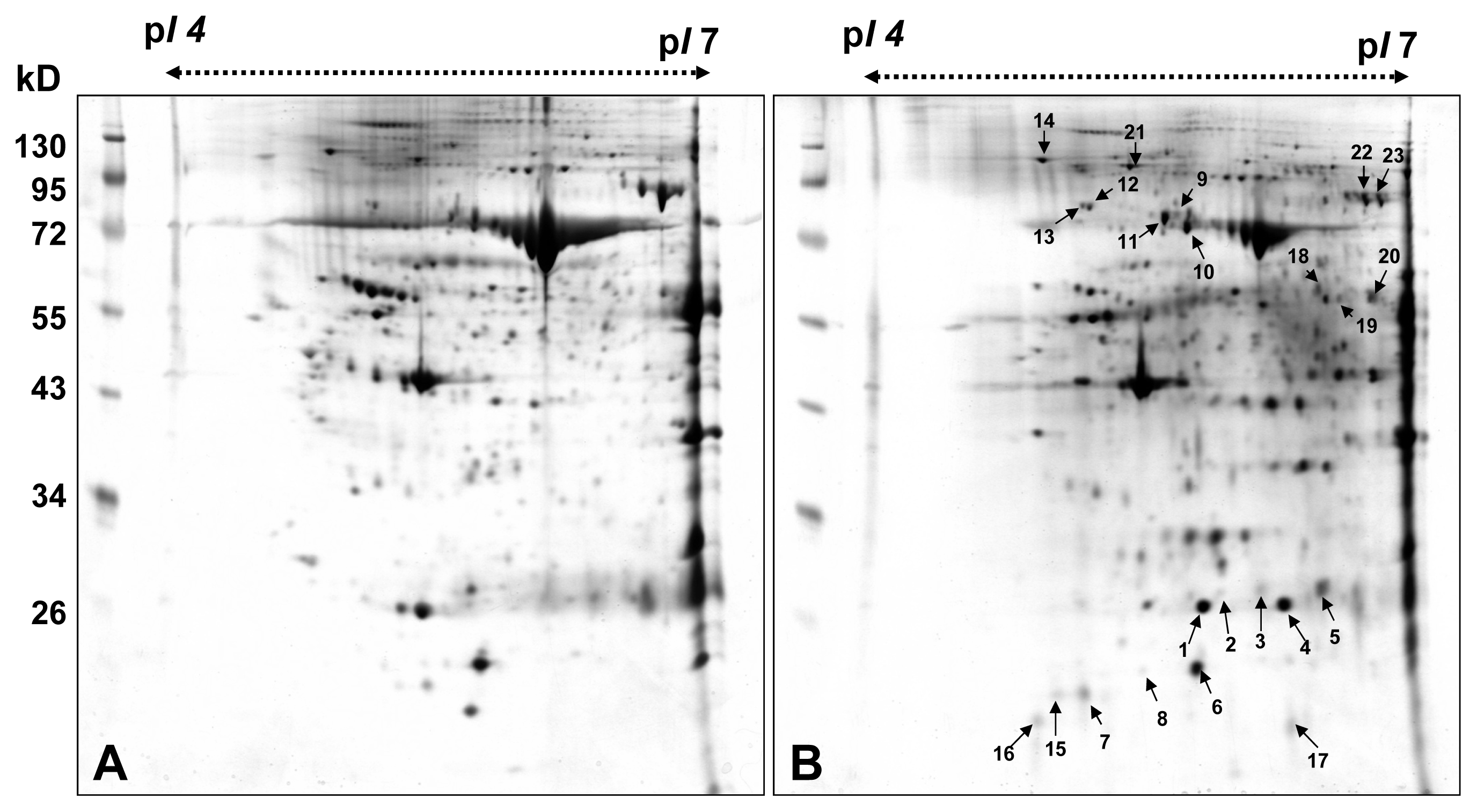
2.1. 2-DE Analysis and Identification of Differential Proteins
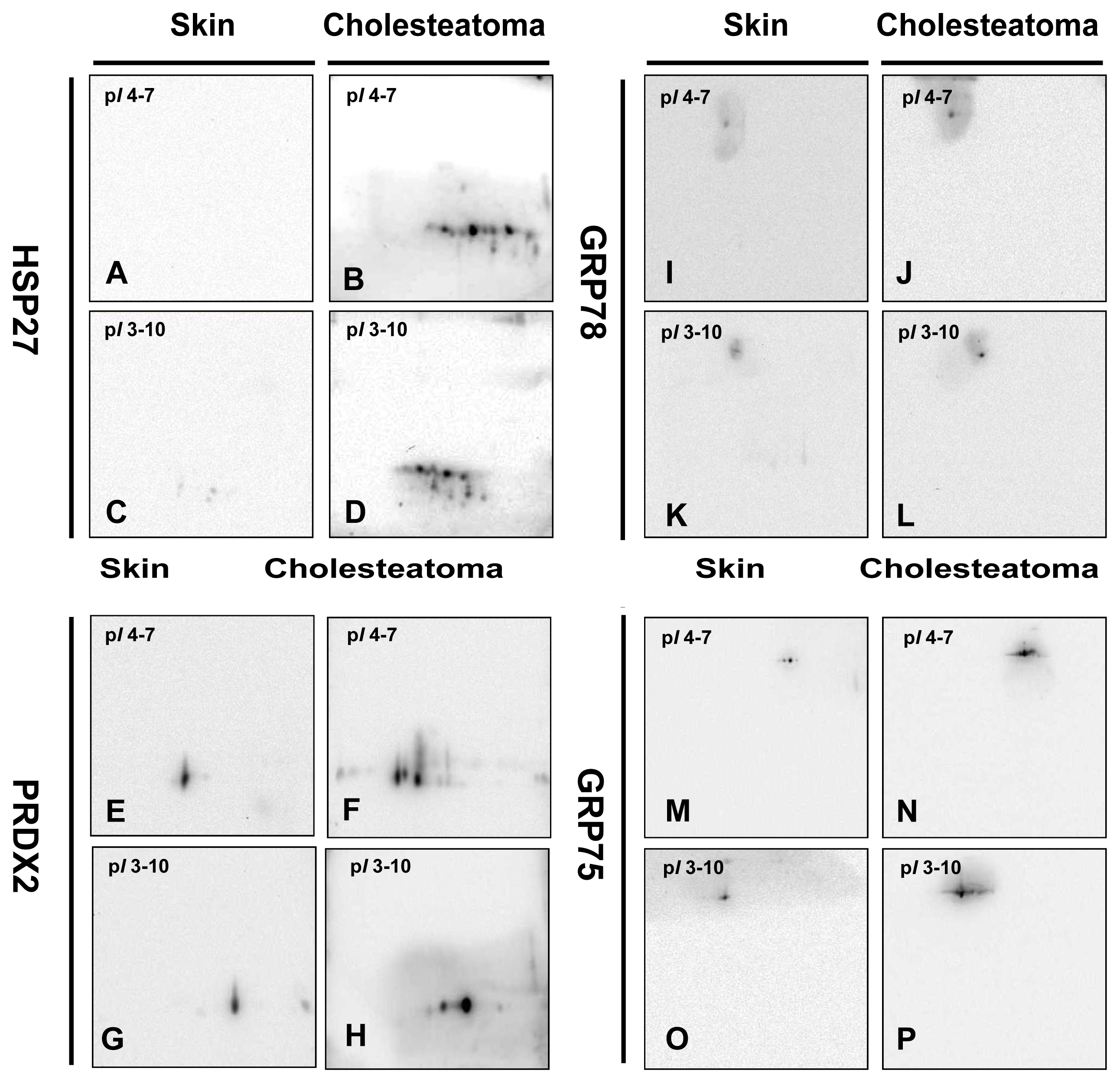
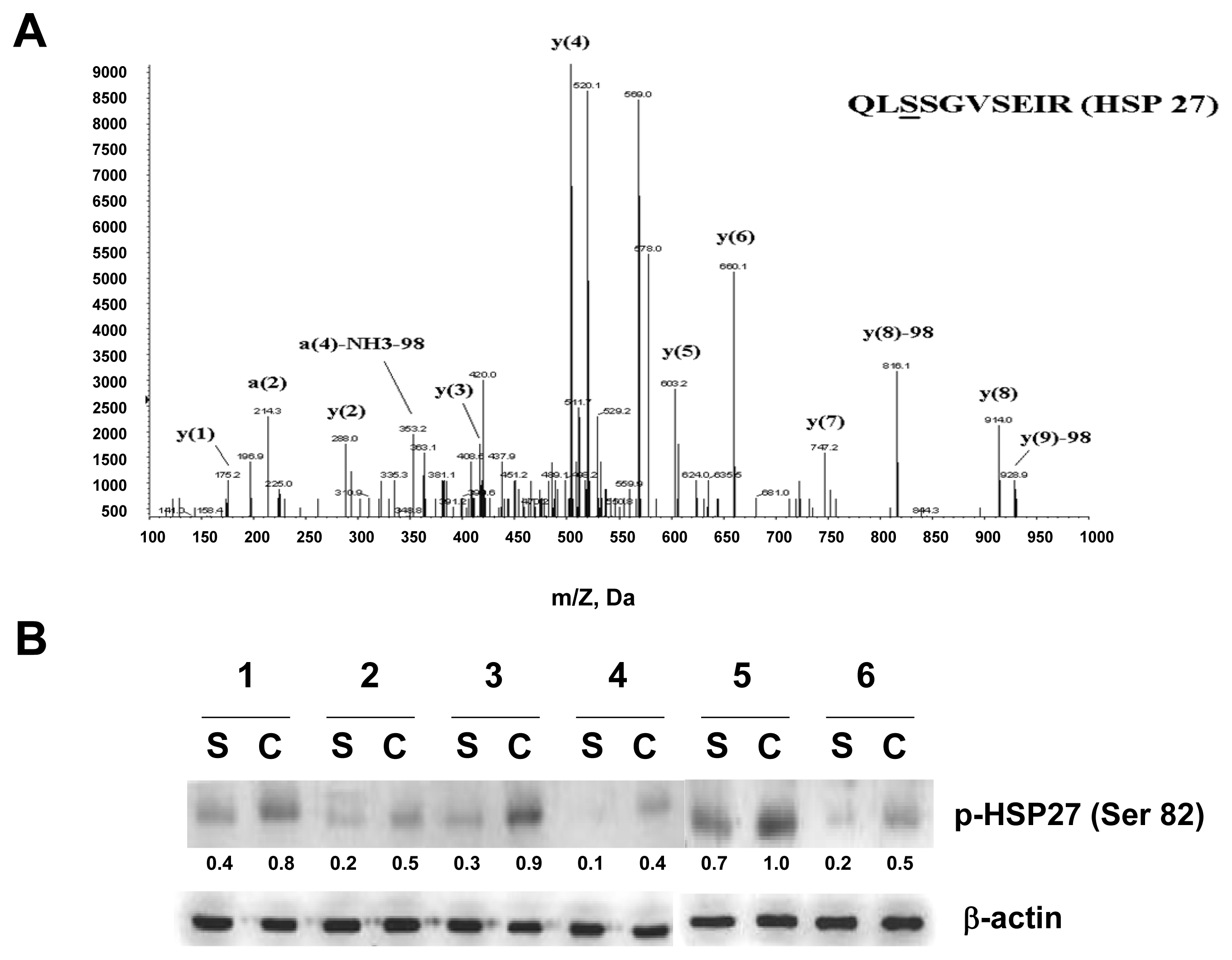
2.2. Phosphorylation of HSP27
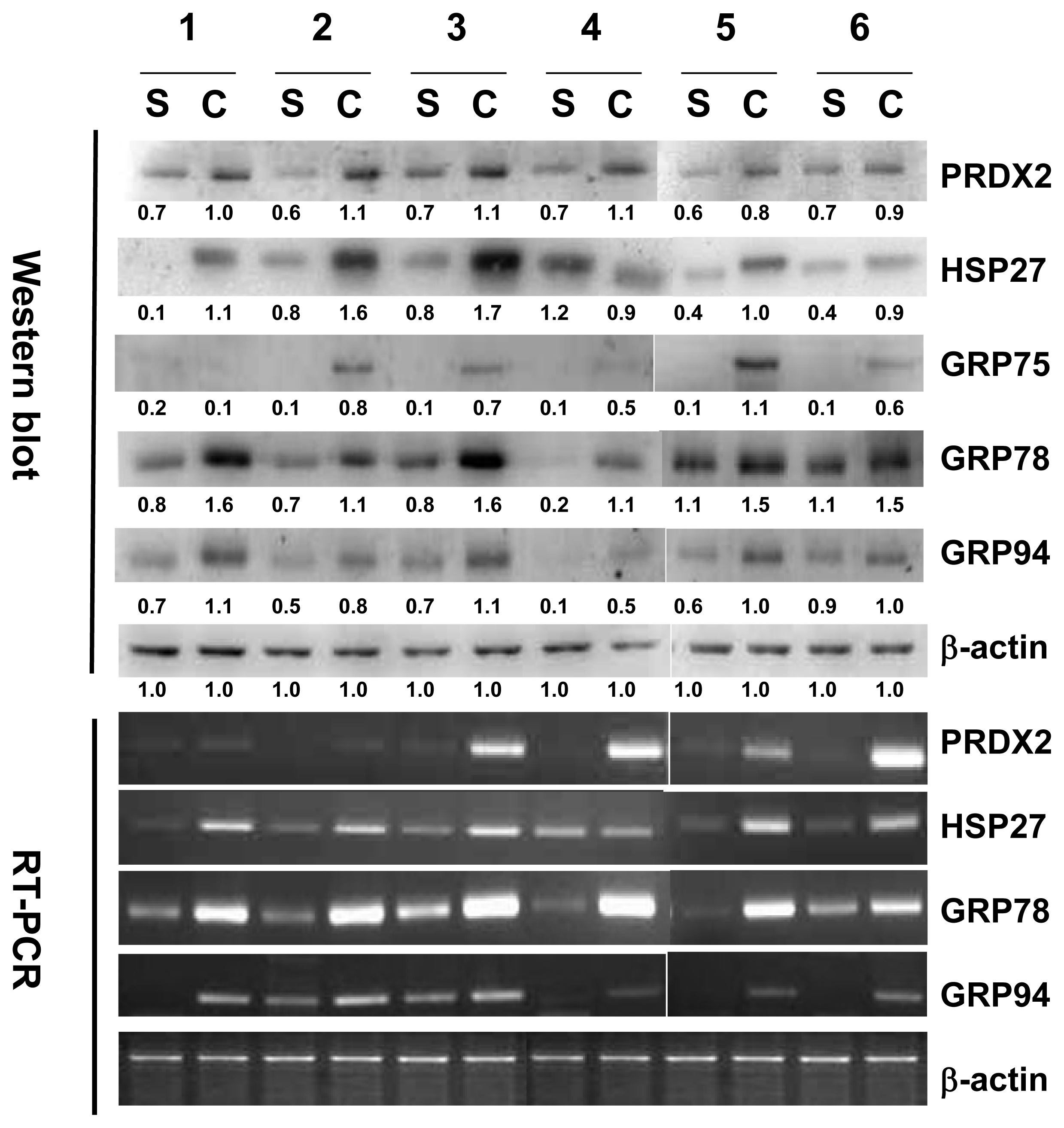
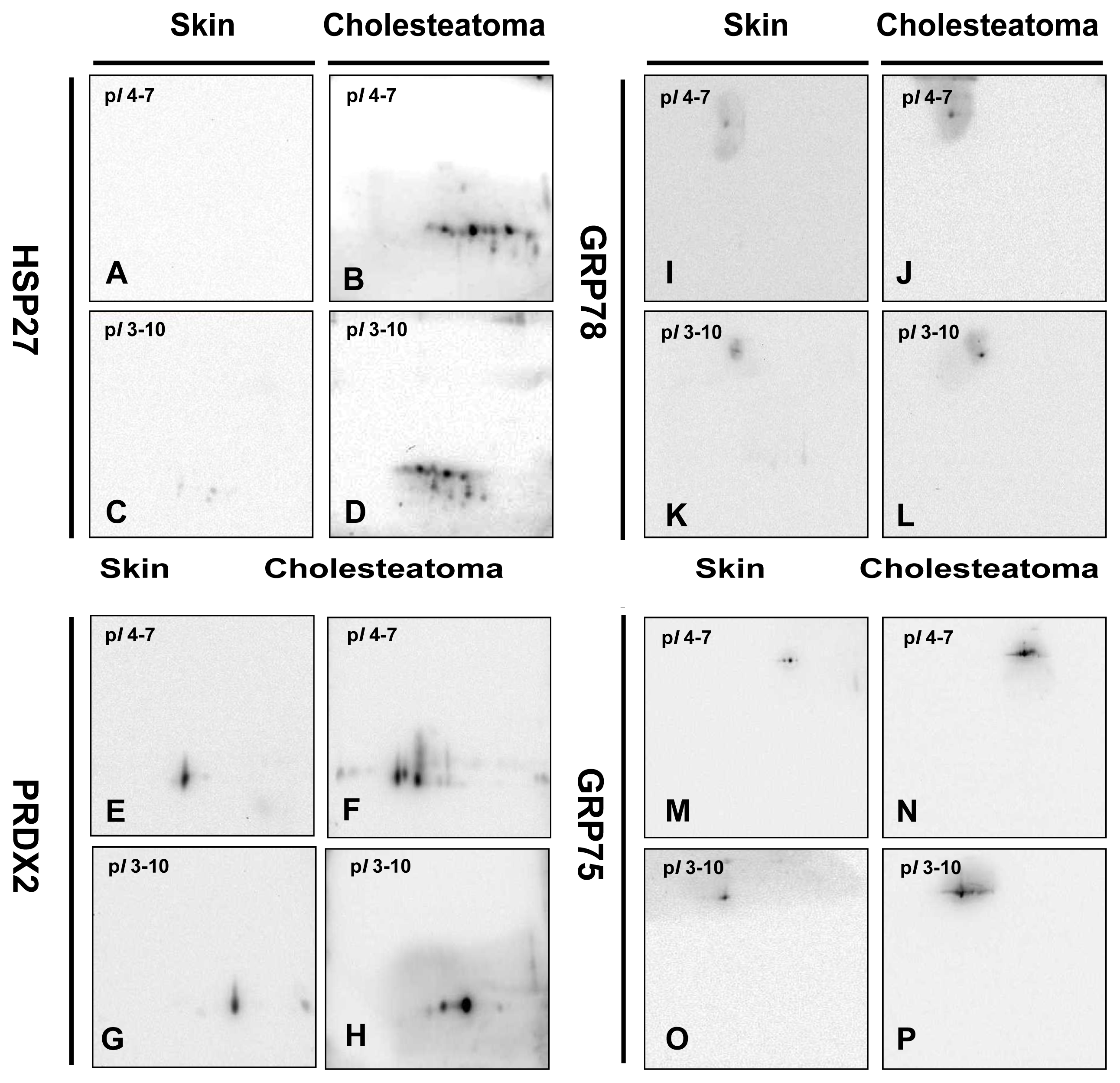
2.3. Validation of HSP27, PRDX2, GRP75, GRP78 and GRP94 by Western Blotting Analysis and RT-PCR
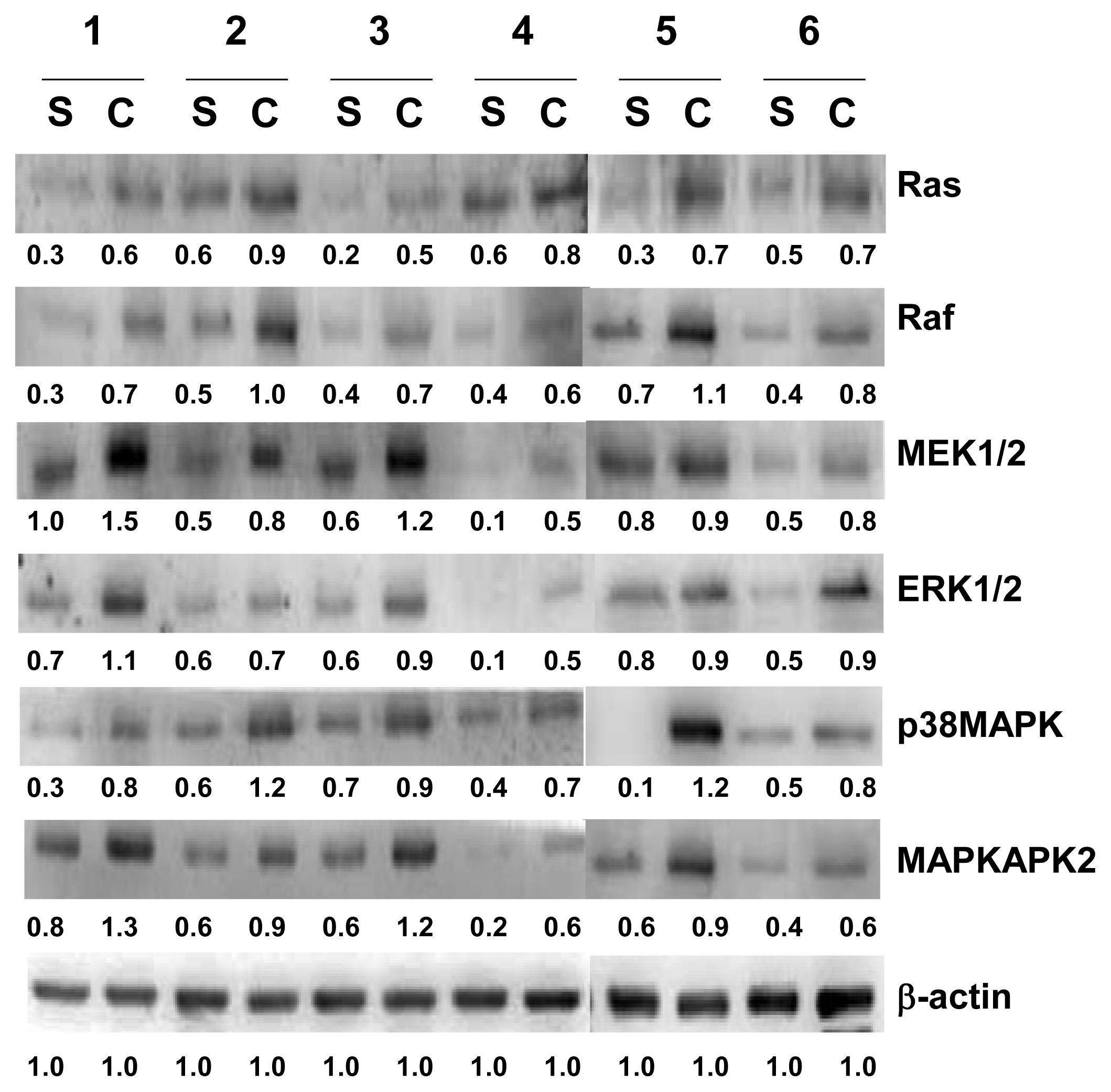
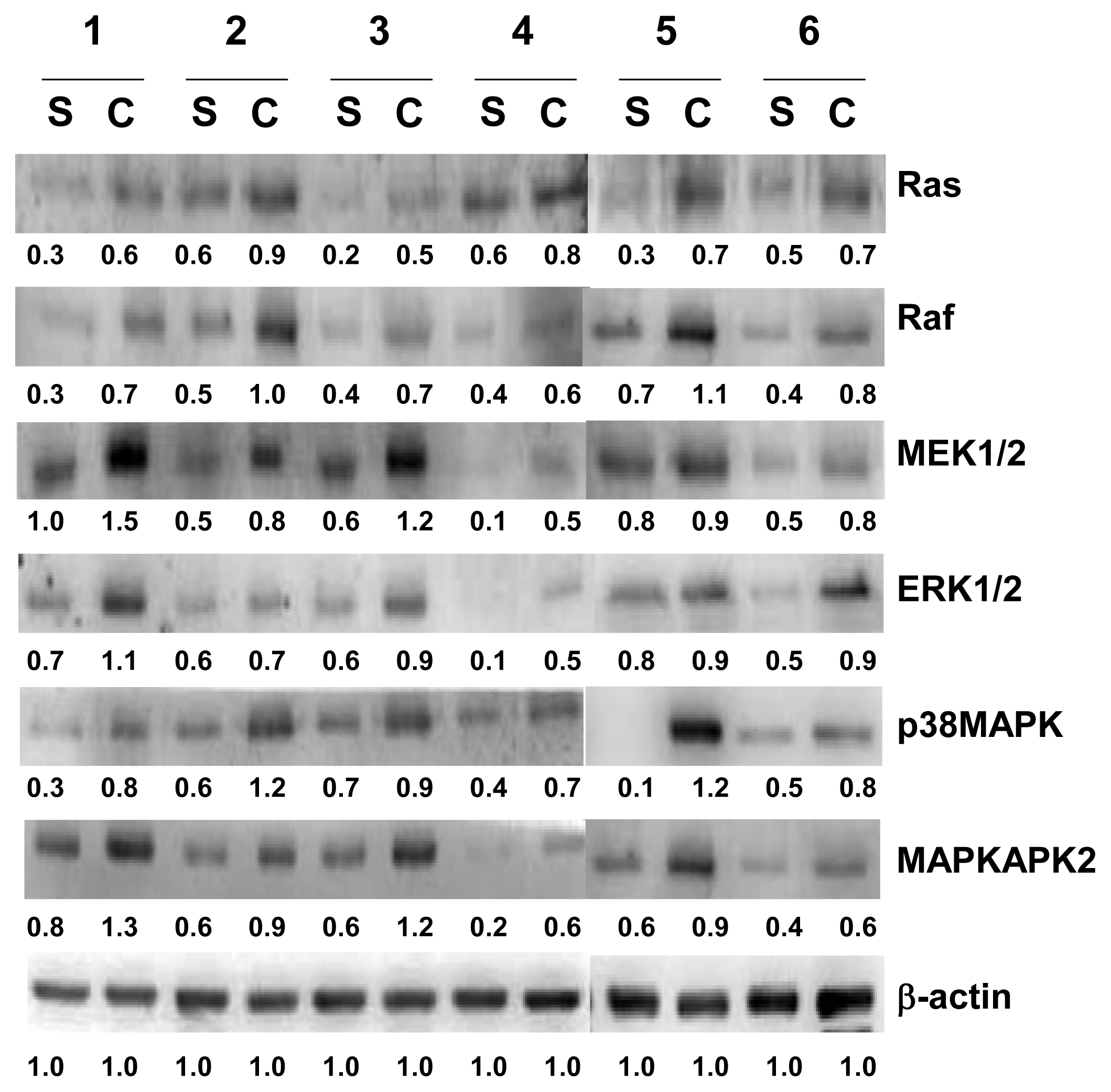
2.4. Verification of Ras, Raf, ERK1/2, MEK1/2, p38MAPK and MAPKAPK2 by Western Blotting Analysis
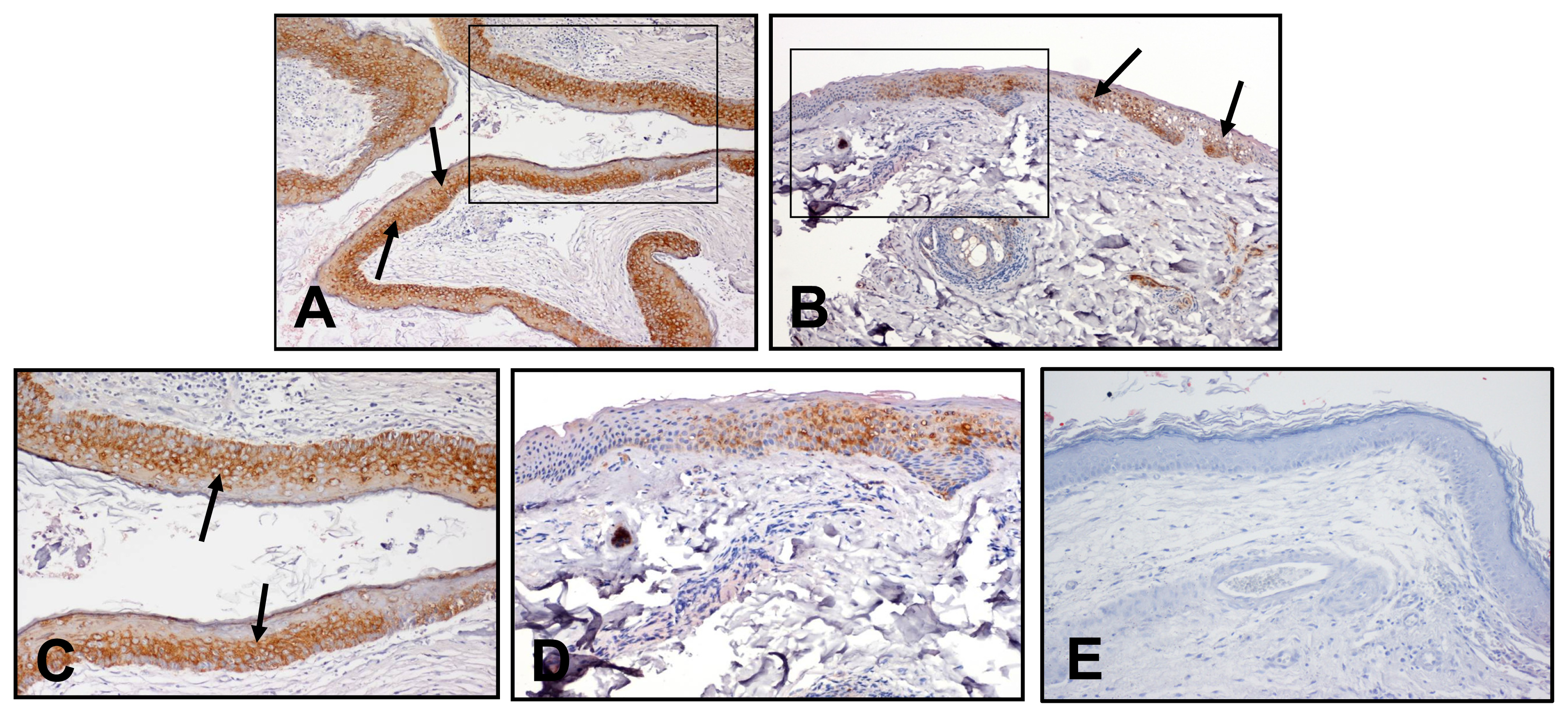
2.5. Immunoreactivity of HSP27
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Sample Preparation
4.3. Two-Dimensional Gel Electrophoresis
4.4. Protein Spot Identification by LC-MS/MS
4.4.1. In-Gel Digestion
4.4.2. LC-MS/MS Analysis and MASCOT Database Searching
4.5. Western Blotting Analysis
4.6. RNA Isolation and RT-PCR
- (1)
- β-actin follows:
- 5′-3′AGAGATGGCCACGGCTGCTT (forward);
- 5′-3′ATTTGCGGTGGACGATGGAG (reverse).
- (2)
- Heat shock protein 27 (HSP27) follows:
- 5′-3′ ACGAGCATGGCTACATCTCC (forward);
- 5′-3′ CTTTACTTGGCGGCAGTCTC (reverse).
- (3)
- Thioredoxin peroxidase 2 (PRDX2) follows:
- 5′-3′GTGTCCTTCGCCAGATCACT (forward);
- 5′-3′ ACGTTGGGCTTAATCGTGTC (reverse).
- (4)
- Glucose-regulated protein 78 (GRP78) follows:
- 5′-3′ TCCTATGTCGCCTTCACT (forward);
- 5′-3′ ACAGACGGGTCATTCCAC (reverse).
- (5)
- Glucose-regulated protein 94 (GRP94) follows:
- 5′-3′ GGGAGGTCACCTTCAAGTCG (forward);
- 5′-3′ GGGTGTAGACGTGGAGCTC (reverse).
4.7. Immunohistochemical Staining of Hsp27
5. Conclusions
Acknowledgements
Conflict of Interest
References
- Kuczkowski, J.; Sakowicz-Burkiewicz, M.; Iżycka-Œwieszewska, E.; Mikaszewski, B.; Pawełczyk, T. Expression of tumor necrosis factor-α, interleukin-1α, interleukin-6 and interleukin-10 in chronic otitis media with bone osteolysis. ORL J. Otorhinolaryngol. Relat. Spec 2011, 73, 93–99. [Google Scholar]
- Haruyam, T.; Furukawa, M.; Kusunoki, T.; Onoda, J.; Ikeda, K. Expression of IL-17 and its role in bone destruction in human middle ear cholesteatoma. ORL J. Otorhinolaryngol. Relat. Spec 2010, 72, 325–331. [Google Scholar]
- Kuczkowski, J.; Sakowicz-Burkiewicz, M.; Iżycka-Œwieszewska, E. Expression of the receptor activator for nuclear factor-κB ligand and osteoprotegerin in chronic otitis media. Am. J. Otolaryngol 2010, 31, 404–409. [Google Scholar]
- Juhn, S.K.; Jung, M.K.; Hoffman, M.D.; Drew, B.R.; Preciado, D.A.; Sausen, N.J.; Jung, T.T.; Kim, B.H.; Park, S.Y.; Lin, J.; et al. The role of inflammatory mediators in the pathogenesis of otitis media and sequelae. Clin. Exp. Otorhinolaryngol 2008, 1, 117–138. [Google Scholar]
- Nason, R.; Jung, J.Y.; Chole, R.A. Lipopolysaccharide-induced osteoclastogenesis from mononuclear precursors: A mechanism for osteolysis in chronic otitis. J. Assoc. Res. Otolaryngol 2009, 10, 151–160. [Google Scholar]
- Hilton, C.W.; Ondrey, F.G.; Wuertz, B.R.; Levine, S.C. Interleukin-8 production in response to tumor necrosis factor-alpha by cholesteatoma keratinocytes in cell culture. Laryngoscope 2011, 121, 372–374. [Google Scholar]
- Helgaland, T.; Engelen, B.; Olsnes, C.; Aarstad, H.J.; Vassbotn, F.S. In vitro cholesteatoma growth and secretion of cytokines. Acta Otolaryngol 2010, 130, 815–819. [Google Scholar]
- Greenberg, J.S.; Manolidis, S. High incidence of complications encountered in chronic otitis media surgery in a U.S. metropolitan public hospital. Otolaryngol. Head Neck Surg 2001, 125, 623–627. [Google Scholar]
- Bagger-Sjoback, D.; Phelps, P.D. Cholesteatoma with extension to the cochlea. Am. J. Otol 1985, 6, 338–343. [Google Scholar]
- Chi, H.P.; Ho, K.Y.; Chai, C.Y.; Ta, C.F.; Wang, L.F.; Lee, K.W.; Kuo, W.R.; Wu, S.C.; Tsai, S.M. Epidermal growth factor expression in middle ear cholesteatoma. Kaohsiung J. Med. Sci 2004, 20, 6–11. [Google Scholar]
- Raynov, A.M.; Choung, Y.H.; Park, H.Y.; Choi, S.J.; Park, K. Establishment and characterization of an in vitro model for cholesteatoma. Clin. Exp. Otorhinolaryngol 2008, 1, 86–91. [Google Scholar]
- Marenda, S.A.; Aufdemorte, T.B. Localization of cytokines in cholesteatoma tissue. Otolaryngol. Head Neck Surg 1995, 112, 359–368. [Google Scholar]
- Yoshikawa, M.; Kojima, H.; Wada, K.; Tsukidate, T.; Okada, N.; Saito, H.; Moriyama, H. Identification of specific gene expression profiles in fibroblasts derived from middle ear cholesteatoma. Arch. Otolaryngol. Head Neck Surg 2006, 132, 734–742. [Google Scholar]
- Raffa, S.; Leone, L.; Scrofani, C.; Monini, S.; Torrisi, M.R.; Barbara, M. Cholesteatoma-associated fibroblasts modulate epithelial growth and differentiation through KGF/FGF7 secretion. Histochem. Cell. Biol 2012, 138, 251–269. [Google Scholar]
- Yamamoto-Fukuda, T.; Aoki, D.; Hishikawa, Y.; Kobayashi, T.; Takahashi, H.; Koji, T. Possible involvement of keratinocyte growth factor and its receptor in enhanced epithelial-cell proliferation and acquired recurrence of middle-ear cholesteatoma. Lab. Invest 2003, 83, 123–136. [Google Scholar]
- Huisman, M.A.; De Heer, E.; Grote, J.J. Survival signaling and terminal differentiation in cholesteatoma epithelium. Acta OtoLaryngol 2007, 127, 424–429. [Google Scholar]
- Liu, C.I.; Chen, C.C.; Chen, J.C.; Su, J.H.; Huang, H.H.; Chen, J.Y.; Wu, Y.J. Proteomic analysis of anti-tumor effects of 11-dehydrosinulariolide on CAL-27 cells. Mar. Drugs 2011, 9, 1254–1272. [Google Scholar]
- Su, T.R.; Lin, J.J.; Chiu, C.C.; Chen, J.Y.F.; Su, J.H.; Cheng, Z.J.; Hwang, W.I.; Huang, H.H.; Wu, Y.J. Proteomic investigation of anti-tumor activities exerted by sinularin against A2058 melanoma cells. Electrophoresis 2012, 33, 1139–1152. [Google Scholar]
- Kim, J.L.; Jung, H.H. Proteomic analysis of cholesteatoma. Acta OtoLaryngol 2004, 124, 783–788. [Google Scholar]
- Shieh, T.J.; Ho, K.Y.; Kuo, W.R.; Chai, C.Y.; Lin, C.S.; Juan, K.H. Evaluation of proliferative activity in middle ear cholesteatoma using proliferating cell nuclear antigen. Kaohsiung J. Med. Sci 1999, 15, 468–474. [Google Scholar]
- Yasuda, E.; Kumada, T.; Takai, S.; Ishisaki, A.; Noda, T.; Matsushima-Nishiwaki, R.; Yoshimi, N.; Kato, K.; Toyoda, H.; Kaneoka, Y.; et al. Attenuated phosphorylation of heat shock protein 27 correlates with tumor progression in patients with hepatocellular carcinoma. Biochem. Biophys. Res. Commun 2005, 337, 337–342. [Google Scholar]
- Yuan, J.; Rozengurt, E. PKD, PKD2, and p38 MAPK mediate Hsp27 serine-82 phosphorylation induced by neurotensin in pancreatic cancer PANC-1 cells. J. Cell. Biochem 2008, 103, 648–662. [Google Scholar]
- Huisman, M.A.; De Heer, E.; Grote, J.J. Sustained extracellular signal-regulated kinase1/2 mitogen-activated protein kinase signalling is related to increased p21 expression in cholesteatoma epithelium. Acta Otolaryngol 2005, 125, 134–140. [Google Scholar]
- Tavaria, M.; Gabriele, T.; Kola, I.; Anderson, R.L. A hitchhiker’s guide to the human Hsp70 family. Cell. Stress Chaperones 1996, 1, 23–28. [Google Scholar]
- Morano, K.A. New tricks for an old dog: The evolving world of Hsp70. Ann. N. Y. Acad. Sci 2007, 1113, 1–14. [Google Scholar]
- Shinoda, H.; Huang, C.C. Heat shock proteins in middle ear cholesteatoma. Otolaryngol. Head Neck Surg 1996, 114, 77–83. [Google Scholar]
- Lee, A.S. The ER chaperone and signaling regulator GRP78/BiP as a monitor of endoplasmic reticulum stress. Methods 2005, 35, 373–381. [Google Scholar]
- Lee, A.S. The glucose-regulated proteins: Stress induction and clinical applications. Trends Biochem. Sci 2001, 26, 504–510. [Google Scholar]
- Hughes, C.S.; Shen, J.W.; Subjeck, J.R. Resistance to etoposide induced by three glucose-regulated stresses in Chinese hamster ovary cells. Cancer Res 1989, 49, 4452–4454. [Google Scholar]
- Jamora, C.; Dennert, G.; Lee, A.S. Inhibition of tumor progression by suppression of stress protein GRP78/BiP induction in fibrosarcoma B/C10ME. Proc. Natl. Acad. Sci. USA 1996, 93, 7690–7694. [Google Scholar]
- Song, M.S.; Park, Y.K.; Lee, J.H.; Park, K. Induction of Glucose-regulated Protein 78 by Chronic Hypoxia in Human Gastric Tumor Cells through a Protein Kinase C-ɛ/ERK/AP-1 Signaling Cascade. Cancer Res 2001, 61, 8322–8330. [Google Scholar]
- Voisine, C.; Craig, E.A.; Zufall, N.; von Ahsen, O.; Pfanner, N.; Voos, W. The protein import motor of mitochondria: Unfolding and trapping of preproteins are distinct and separable functions of matrix Hsp70. Cell 1999, 97, 565–574. [Google Scholar]
- Ma, Z.; Izumi, H.; Kanai, M.; Kabuyama, Y.; Ahn, N.G.; Fukasawa, K. Mortalin controls centrosome duplication via modulating centrosomal localization of p53. Oncogene 2006, 25, 5377–5390. [Google Scholar]
- Lim, J.H.; Martin, F.; Guiard, B.; Pfanner, N.; Voos, W. The mitochondrial Hsp70-dependent import system actively unfolds preproteins and shortens the lag phase of translocation. EMBO J 2001, 20, 941–950. [Google Scholar]
- Sacht, G.; Brigelius-Flohe, R.; Kiess, M.; Sztajer, H.; Flohe, L. ATP-sensitive association of mortalin with the IL-1 receptor type I. Biofactors 1999, 9, 49–60. [Google Scholar]
- Takano, S.; Wadhwa, R.; Mitsui, Y.; Kaul, S.C. Identification and characterization of molecular interactions between glucose-regulated proteins (GRPs) mortalin/GRP75/peptide-binding protein 74 (PBP74) and GRP94. Biochem. J 2001, 357, 393–398. [Google Scholar]
- Wadhwa, R.; Yaguchi, T.; Hasan, M.K.; Taira, K.; Kaul, S.C. Mortalin-MPD (mevalonate pyrophosphate decarboxylase) interactions and their role in control of cellular proliferation. Biochem. Biophys. Res. Commun 2003, 302, 735–742. [Google Scholar]
- Mizukoshi, E.; Suzuki, M.; Misono, T.; Loupatov, A.; Munekata, E.; Kaul, S.C.; Wadhwa, R.; Imamura, T. Cell-cycle dependent tyrosine phosphorylation on mortalin regulates its interaction with fibroblast growth factor-1. Biochem. Biophys. Res. Commun 2001, 280, 1203–1209. [Google Scholar]
- Yaguchi, T.; Aida, S.; Kaul, S.C.; Wadhwa, R. Involvement of mortalin in cellular senescence from the perspective of its mitochondrial import, chaperone, and oxidative stress management functions. Ann. N. Y. Acad. Sci 2007, 1100, 306–311. [Google Scholar]
- Wadhwa, R.; Takano, S.; Taira, K.; Kaul, S.C. Reduction in mortalin level by its antisense expression causes senescence-like growth arrest in human immortalized cells. J. Gene Med 2004, 6, 439–444. [Google Scholar]
- Langer, R.; Feith, M.; Siewert, J.R.; Wester, H.J.; Hoefler, H. Expression and clinical significance of glucose regulated proteins GRP78 (BiP) and GRP94 (GP96) in human adenocarcinomas of the esophagus. BMC Cancer 2008, 8. [Google Scholar] [CrossRef]
- Zheng, H.C.; Takahashi, H.; Li, X.H.; Hara, T.; Masuda, S.; Guan, Y.F.; Takano, Y. Overexpression of GRP78 and GRP94 are markers for aggressive behavior and poor prognosis in gastric carcinomas. Hum. Pathol 2008, 39, 1042–1049. [Google Scholar]
- Fu, Y.; Lee, A.S. Glucose regulated proteins in cancer progression, drug resistance and immunotherapy. Cancer Biol. Ther 2006, 5, 741–744. [Google Scholar]
- Kubota, K.; Lee, D.H.; Tsuchiya, M.; Young, C.S.; Everett, E.T.; Martinez-Mier, E.A.; Snead, M.L.; Nguyen, L.; Urano, F.; Bartlett, J.D. Fluoride induces endoplasmic reticulum stress in ameloblasts responsible for dental enamel formation. J. Biol. Chem 2005, 280, 23194–23202. [Google Scholar]
- Garrido, C.; Brunet, M.; Didelot, C.; Zermati, Y.; Schmitt, E.; Kroemer, G. Heat shock proteins 27 and 70: Anti-apoptotic proteins with tumorigenic properties. Cell. Cycle 2006, 5, 2592–2601. [Google Scholar]
- Bruey, J.M.; Paul, C.; Fromentin, A.; Hilpert, S.; Arrigo, A.P.; Solary, E.; Garrido, C. Differential regulation of HSP27 oligomerization in tumor cells grown in vitro and in vivo. Oncogene 2000, 19, 4855–4863. [Google Scholar]
- Wang, A.; Liu, X.; Sheng, S.; Ye, H.; Peng, T.; Shi, F.; Crowe, D.L.; Zhou, X. Dysregulation of heat shock protein 27 expression in oral tongue squamous cell carcinoma. BMC Cancer 2009, 9. [Google Scholar] [CrossRef]
- Xu, L.; Chen, S.; Bergan, R.C. MAPKAPK2 and HSP27 are downstream effectors of p38 MAP kinase-mediated matrix metalloproteinase type 2 activation and cell invasion in human prostate cancer. Oncogene 2006, 25, 2987–2998. [Google Scholar]
- Kostenko, S.; Moens, U. Heat shock protein 27 phosphorylation: Kinases, phosphatases, functions and pathology. Cell. Mol. Life Sci 2009, 66, 3289–3307. [Google Scholar]
- Gibbs, S.; Silva Pinto, A.N.; Murli, S.; Huber, M.; Hohl, D.; Ponec, M. Epidermal growth factor and keratinocyte growth factor differentially regulate epidermal migration, growth, and differentiation. Wound Repair Regen 2000, 8, 192–203. [Google Scholar]
- Shirakata, Y. Regulation of epidermal keratinocytes by growth factors. J. Dermatol. Sci 2010, 59, 73–80. [Google Scholar]
- Niwa, M.; Hotta, K.; Hara, A.; Hirade, K.; Ito, H.; Kato, K.; Kozawa, O. TNF-α decreases hsp 27 in human blood mononuclear cells: Involvement of protein kinase c. Life Sci 2006, 80, 181–186. [Google Scholar]
- Sudhoff, H.; Dazert, S.; Gonzales, A.M.; Borkowski, G.; Park, S.Y.; Baird, A.; Hildmann, H.; Ryan, A.F. Angiogenesis and angiogenic growth factors in middle ear cholesteatoma. Otol. Neurotol 2000, 21, 793–798. [Google Scholar]
- Evans, I.M.; Britton, G.; Zachary, I.C. Vascular endothelial growth factor induces heat shock protein (HSP) 27 serine 82 phosphorylation and endothelial tubulogenesis via protein kinase D and independent of p38 kinase. Cell Signal 2008, 20, 1375–1384. [Google Scholar]
- Schluter, H.; Apweiler, R.; Holzhutter, H.G.; Jungblut, P.R. Finding one’s way in proteomics: A protein species nomenclature. Chem. Cent. J. 2009, 3. [Google Scholar] [CrossRef]
- Huang, C.C.; Chen, C.T.; Huang, T.S.; Shinoda, H. Mediation of signal transduction in keratinocytes of human middle ear cholesteatoma by ras protein. Eur. Arch. Otorhinolaryngol 1996, 253, 385–389. [Google Scholar]
- Wong, J.W.; Shi, B.; Farboud, B.; McClaren, M.; Shibamoto, T.; Cross, C.E.; Isseroff, R.R. Ultraviolet B-mediated phosphorylation of the small heat shock protein HSP27 in human keratinocytes. J. Investig. Dermatol 2000, 115, 427–434. [Google Scholar]
- Robitaille, H.; Simard-Bisson, C.; Larouche, D.; Tanguay, R.M.; Blouin, R.; Germain, L. The small heat-shock protein Hsp27 undergoes ERK-dependent phosphorylation and redistribution to the cytoskeleton in response to dual leucine zipper-bearing kinase expression. J. Invest. Dermatol 2010, 130, 74–85. [Google Scholar]
- Mitev, V.; Miteva, L. Signal transduction in keratinocytes. Exp. Dermatol 1999, 8, 96–108. [Google Scholar]
- Rozengurt, E. Protein kinase D signaling: Multiple biological functions in health and disease. Physiology (Bethesda) 2011, 26, 23–33. [Google Scholar]
- Valyi-Nagy, I.; Jensen, P.J.; Albelda, S.M.; Rodeck, U. Cytokine-induced expression of transforming growth factor-alpha and the epidermal growth factor receptor in neonatal skin explants. J. Invest. Dermatol 1992, 99, 350–356. [Google Scholar]
- Saunders, N.A.; Jetten, A.M. Control of growth regulatory and differentiation-specific genes in human epidermal keratinocytes by interferon gamma. Antagonism by retinoic acid and transforming growth factor beta 1. J. Biol. Chem 1994, 269, 2016–2022. [Google Scholar]
- Yun, S.J.; Seo, J.J.; Chae, J.Y.; Lee, S.C. Peroxiredoxin I and II are up-regulated during differentiation of epidermal keratinocytes. Arch. Dermatol. Res 2005, 296, 555–559. [Google Scholar]
- Myers, E.N.; Park, K.; Chun, Y.; Lee, D.; Hwang, S. Signal transduction pathway in human middle ear cholesteatoma. Otolaryngol. Head Neck Surg 1999, 120, 899–904. [Google Scholar]
- Ryu, J.; Park, S.G.; Park, B.C.; Choe, M.; Lee, K.S.; Cho, J.W. Proteomic analysis of psoriatic skin tissue for identification of differentially expressed proteins: Up-regulation of GSTP1, SFN and PRDX2 in psoriatic skin. Int. J. Mol. Med 2011, 28, 785–792. [Google Scholar]
- Wu, Y.J.; Chen, H.M.; Wu, T.T.; Wu, J.S.; Chu, R.M.; Juang, R.H. Preparation of monoclonal antibody bank against whole water-soluble proteins from rapid-growing bamboo shoots. Proteomics 2006, 6, 5898–5902. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot No. | Protein name | Accession No. | Calculate Mw/pI | Peptide matched | Sequence covered % | MASCO T score | Biological process |
|---|---|---|---|---|---|---|---|
| 1 | Heat-shock protein 27 | P04792 | 22.7/5.98 | 41 | 61 | 547 | negative regulation of apoptotic process positive regulation of angiogenesis positive regulation of endothelial cell chemotaxis y positive regulation of interleukin-1 beta production positive regulation of tumor necrosis factor biosynthetic process regulation of I-kappaB kinase/NF-kappaB cascade regulation of translational initiation response to unfolded protein |
| 2 | Heat-shock protein 27 | P04792 | 22.7/5.98 | 10 | 30 | 190 | |
| 3 | Heat-shock protein 27 | P04792 | 22.7/5.98 | 9 | 25 | 140 | |
| 4 | Heat-shock protein 27 | P04792 | 22.7/5.98 | 36 | 48 | 385 | |
| 5 | Heat-shock protein 27 | P04792 | 22.7/5.98 | 10 | 37 | 150 | |
| 6 | Peroxiredoxin-2 | P32119 | 21.8/5.66 | 19 | 43 | 174 | hydrogen peroxide catabolic process |
| 7 | Peroxiredoxin-2 | P32119 | 21.8/5.66 | 40 | 44 | 641 | negative regulation of apoptotic process |
| 8 | Peroxiredoxin-2 | P32119 | 21.8/5.66 | 26 | 42 | 336 | removal of superoxide radicals |
| 9 | 75 kDa glucose-regulated protein | P38646 | 73.6/5.87 | 13 | 15 | 157 | negative regulation of apoptotic process protein export from nucleus protein folding protein targeting to mitochondrion |
| 10 | Heat shock cognate 71 kDa protein | P11142 | 70.8/5.37 | 39 | 34 | 339 | mRNA metabolic process negative regulation of transcription protein folding regulation of cell cycle response to unfolded protein |
| 11 | Heat shock cognate 71 kDa protein | P11142 | 70.8/5.37 | 59 | 46 | 624 | |
| 12 | 78 kDa glucose-regulated protein | P11021 | 72.2/5.07 | 15 | 16 | 123 | ER overload response ER-associated protein catabolic process negative regulation of apoptotic process positive regulation of protein ubiquitination regulation of protein folding in endoplasmic reticulum |
| 13 | 78 kDa glucose-regulated protein | P11021 | 72.2/5.07 | 55 | 42 | 719 | |
| 14 | 94 kDa glucose-regulated protein | P14625 | 92.4/4.76 | 75 | 37 | 799 | ER-associated protein catabolic process actin rod assembly activation of signaling protein activity involved in unfolded protein response negative regulation of apoptotic process protein folding |
| 15 | Uncharacterize d protein C7orf24 | O75223 | 20.9/5.07 | 13 | 26 | 158 | glutathione biosynthetic process release of cytochrome c from mitochondria |
| 16 | Uncharacterize d protein C7orf24 | O75223 | 20.9/5.07 | 7 | 18 | 104 | |
| 17 | NEDD8-conjugating enzyme Ubc12 | P61081 | 20.8/7.57 | 3 | 12 | 35 | protein neddylation |
| 18 | Serum albumin precursor | P02768 | 69.3/5.92 | 41 | 25 | 388 | lipoprotein metabolic process maintenance of mitochondrion location negative regulation of apoptotic process response to nutrient |
| 19 | Glial fibrillary acidic protein | P14136 | 49.8/5.42 | 11 | 6 | 141 | extracellular matrix organization intermediate filament organization response to wounding |
| 20 | Ig alpha-1 chain C region | P01876 | 37.6/6.08 | 6 | 9 | 81 | immune response protein-chromophore linkage |
| 21 | Transitional endoplasmic reticulum ATPase | P55072 | 89.2/5.14 | 27 | 21 | 176 | ER-associated protein catabolic process activity involved in apoptotic process double-strand break repair endoplasmic reticulum unfolded protein response protein ubiquitination |
| 22 | Serotransferrin precursor | P02787 | 77.0/6.81 | 31 | 23 | 306 | cellular iron ion homeostasis transferrin transport transmembrane transport |
| 23 | Serotransferrin precursor | P02787 | 77.0/6.81 | 20 | 18 | 181 | |
| Patient No. | Sex | Age | Hsp27 staining score | |
|---|---|---|---|---|
| Cholesteatoma a | Retroauricular skin b | |||
| 1 | Male | 22 | 12 | 4 |
| 2 | Female | 13 | 12 | 6 |
| 3 | Female | 56 | 12 | 6 |
| 4 | Female | 68 | 6 | 3 |
| 5 | Female | 31 | 12 | 6 |
| 6 | Male | 35 | 9 | 2 |
| 7 | Female | 58 | 9 | 2 |
| 8 | Male | 50 | 8 | 3 |
| 9 | Female | 57 | 8 | 2 |
| 10 | Female | 26 | 9 | 4 |
| 11 | Female | 14 | 12 | 6 |
| 12 | Female | 57 | 8 | 3 |
| Mean | 40.5 | 9.7 | 4.0 | |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ho, K.Y.; Yeh, T.S.; Huang, H.H.; Hung, K.F.; Chai, C.Y.; Chen, W.T.; Tsai, S.M.; Chang, N.C.; Chien, C.Y.; Wang, H.M.; et al. Upregulation of Phosphorylated HSP27, PRDX2, GRP75, GRP78 and GRP94 in Acquired Middle Ear Cholesteatoma Growth. Int. J. Mol. Sci. 2013, 14, 14439-14459. https://doi.org/10.3390/ijms140714439
Ho KY, Yeh TS, Huang HH, Hung KF, Chai CY, Chen WT, Tsai SM, Chang NC, Chien CY, Wang HM, et al. Upregulation of Phosphorylated HSP27, PRDX2, GRP75, GRP78 and GRP94 in Acquired Middle Ear Cholesteatoma Growth. International Journal of Molecular Sciences. 2013; 14(7):14439-14459. https://doi.org/10.3390/ijms140714439
Chicago/Turabian StyleHo, Kuen Yao, Tai Sheng Yeh, Han Hsiang Huang, Kuo Feng Hung, Chee Yin Chai, Wan Tzu Chen, Shih Meng Tsai, Ning Chia Chang, Chen Yu Chien, Hsun Mo Wang, and et al. 2013. "Upregulation of Phosphorylated HSP27, PRDX2, GRP75, GRP78 and GRP94 in Acquired Middle Ear Cholesteatoma Growth" International Journal of Molecular Sciences 14, no. 7: 14439-14459. https://doi.org/10.3390/ijms140714439
APA StyleHo, K. Y., Yeh, T. S., Huang, H. H., Hung, K. F., Chai, C. Y., Chen, W. T., Tsai, S. M., Chang, N. C., Chien, C. Y., Wang, H. M., & Wu, Y. J. (2013). Upregulation of Phosphorylated HSP27, PRDX2, GRP75, GRP78 and GRP94 in Acquired Middle Ear Cholesteatoma Growth. International Journal of Molecular Sciences, 14(7), 14439-14459. https://doi.org/10.3390/ijms140714439