Insight into Differential Responses of Upland and Paddy Rice to Drought Stress by Comparative Expression Profiling Analysis

Abstract
:1. Introduction
2. Results and Discussion
2.1. Performance of Upland and Paddy Rice under Normal and Drought Conditions
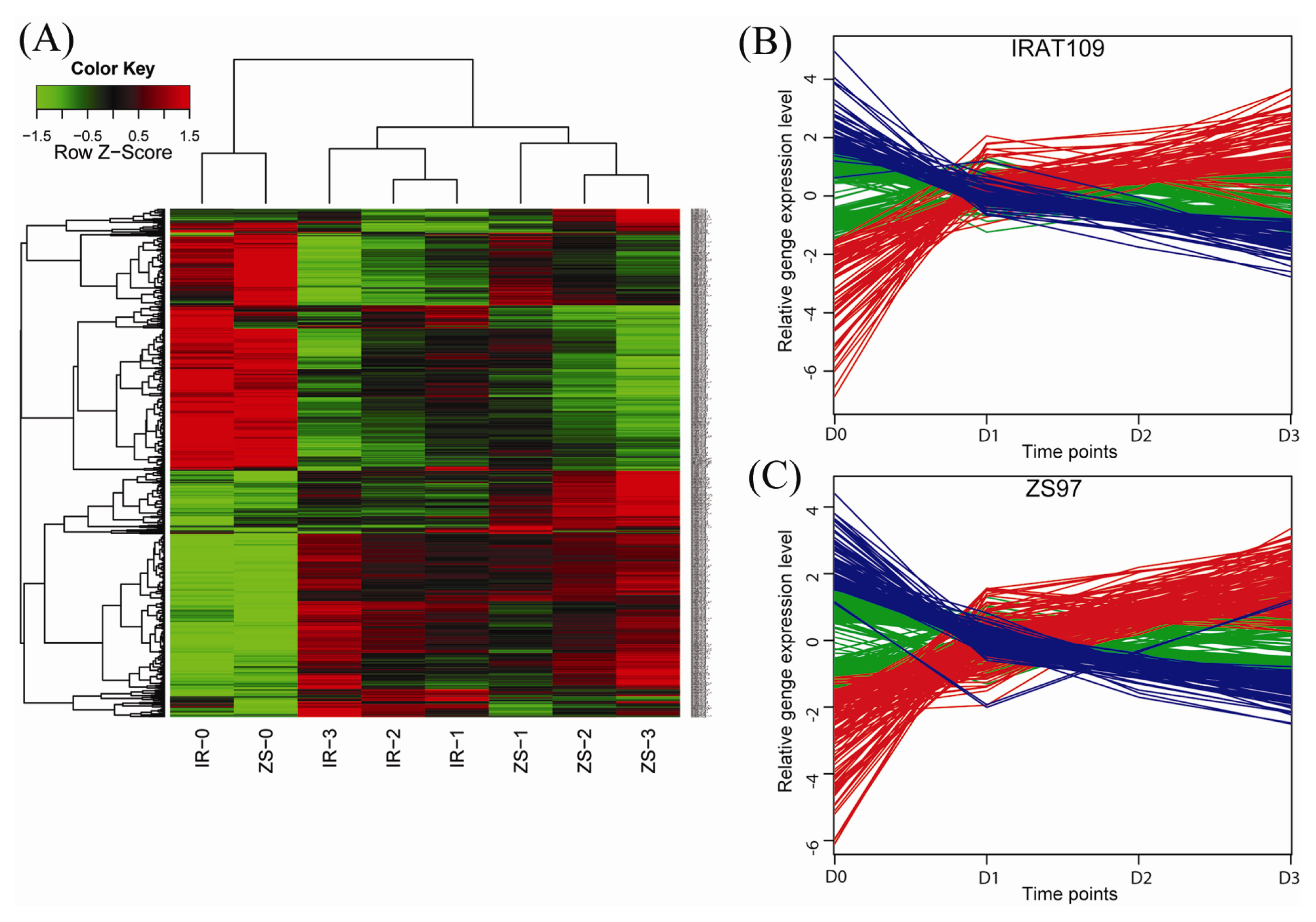
2.2. Different Drought-Responsive Profiles of the Two Rice Genotypes
2.3. Dynamic Patterns of the Drought-Responsive Genes
2.4. Differentially Expressed Genes in the Intervals of Drought Resistance-Related QTL
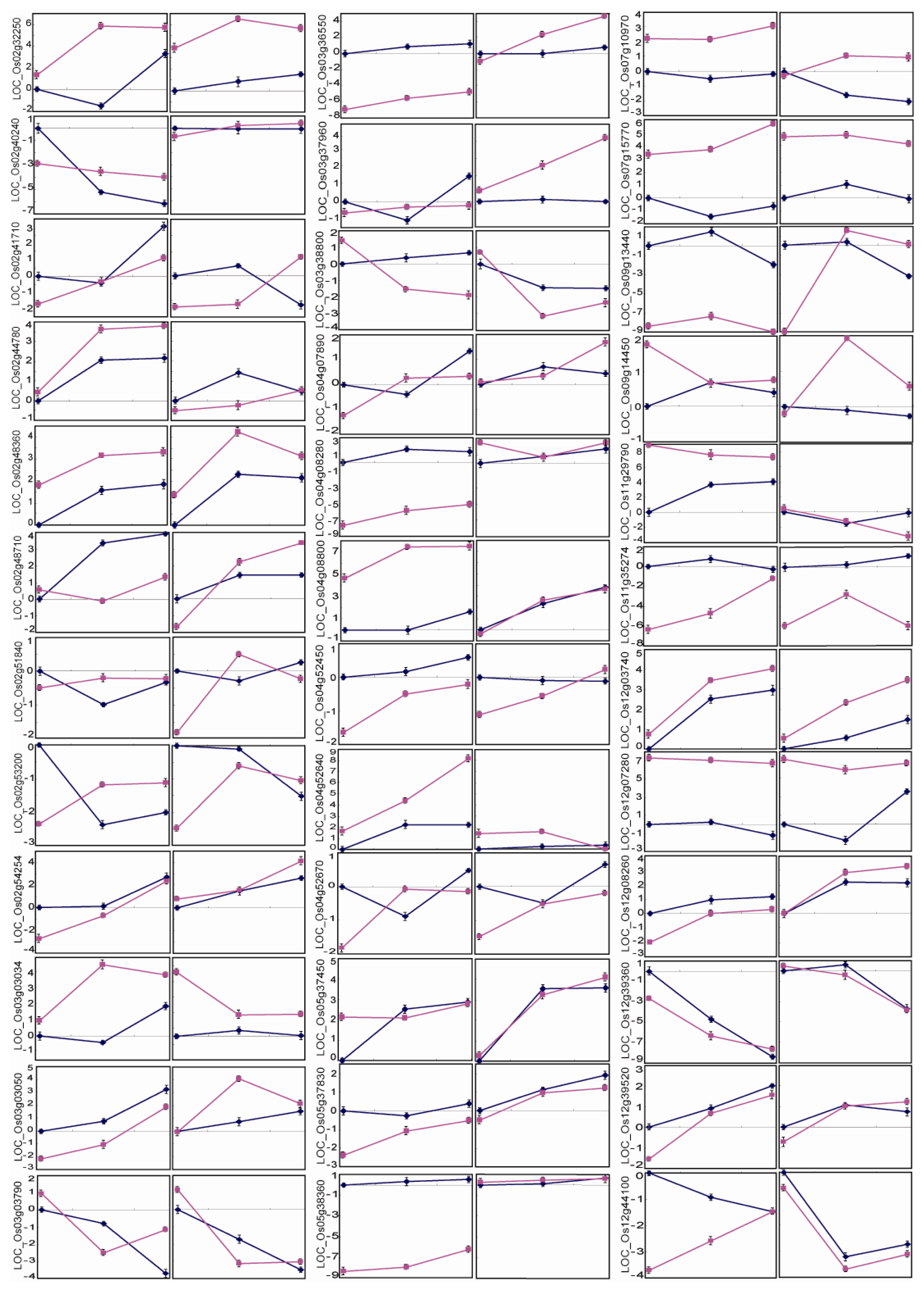
2.5. Differential Expression Patterns of Drought-Responsive Genes in the QTL Intervals
2.6. Discussion
2.6.1. Mechanism of Drought Resistance at the Reproductive Stage
2.6.2. Candidate Genes for QTL Related to Drought Resistance
3. Experimental Methods
3.1. Plant Materials, Cultivation and Stress Treatment
3.2. Microarray and Initial Data Analysis
3.3. Physiological Analysis
3.4. Quantitative Gene Expression Level by Real-Time PCR Analysis
3.5. Accession Numbers
4. Conclusions
Supplementary Information
ijms-14-05214-s001.docAcknowledgments
Conflict of Interest
References
- Zhang, Q. Strategies for developing Green Super Rice. Proc. Natl. Acad. Sci. USA 2007, 104, 16402–16409. [Google Scholar]
- Boonjung, H.; Fukai, S. Effects of soil water deficit at different growth stages on rice growth and yield under upland conditions. 2. Phenology, biomass production and yield. Field Crops Res 1996, 48, 47–55. [Google Scholar]
- Pantuwan, G.; Fukai, S.; Cooper, M.; Rajatasereekul, S.; O’Toole, J.C. Yield response of rice (Oryza sativa L.) genotypes to drought under rainfed lowlands. 2. Selection of drought-resistant genotypes. Field Crops Res 2002, 73, 169–180. [Google Scholar]
- Nguyen, H.N.; Babu, R.C.; Blum, A. Breeding for drought resistance in rice: physiology and molecular genetics consideration. Crop Sci 1997, 17, 1426–1434. [Google Scholar]
- Luo, L.J. Breeding for water-saving and drought-resistance rice (WDR) in China. J. Exp. Bot 2010, 61, 3509–3517. [Google Scholar]
- Yue, B.; Xiong, L.; Xue, W.; Xing, Y.; Luo, L.; Xu, C. Genetic analysis for drought resistance of rice at reproductive stage in field with different types of soil. Theor. Appl. Genet 2005, 111, 1127–1136. [Google Scholar]
- Seki, M.; Narusaka, M.; Abe, H.; Kasuga, M.; Yamaguchi-Shinozaki, K.; Carninci, P.; Hayashizaki, Y.; Shinozaki, K. Monitoring the expression pattern of 1300 Arabidopsis genes under drought and cold stresses by using a full-length cDNA microarray. Plant Cell 2001, 13, 61–72. [Google Scholar]
- Rabbani, M.A.; Maruyama, K.; Abe, H.; Khan, M.A.; Katsura, K.; Ito, Y.; Yoshiwara, K.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Monitoring expression profiles of rice genes under cold, drought, and high-salinity stresses and abscisic acid application using cDNA microarray and RNA gel-blot analyses. Plant Physiol 2003, 133, 1755–1767. [Google Scholar]
- Zhang, W.; Ruan, J.; Ho, T.H.; You, Y.; Yu, T.; Quatrano, R.S. Cis-regulatory element based targeted gene finding: genome-wide identification of abscisic acid- and abiotic stress-responsive genes in Arabidopsis thaliana. Bioinformatics 2005, 21, 3074–3081. [Google Scholar]
- Ueda, A.; Kathiresan, A.; Bennett, J.; Takabe, T. Comparative transcriptome analyses of barley and rice under salt stress. Theor. Appl. Genet 2006, 112, 1286–1294. [Google Scholar]
- Walia, H.; Wilson, C.; Ismail, A.M.; Close, T.J.; Cui, X. Comparing genomic expression patterns across plant species reveals highly diverged transcriptional dynamics in response to salt stress. BMC Genomics 2009, 10, 398. [Google Scholar]
- Wang, H.; Zhang, H.; Gao, F.; Li, J.; Li, Z. Comparison of gene expression between upland and lowland rice cultivars under water stress using cDNA microarray. Theor. Appl. Genet 2007, 115, 1109–1126. [Google Scholar]
- Degenkolbe, T.; Do, P.T.; Zuther, E.; Repsilber, D.; Walther, D.; Hincha, D.K.; Kohl, K.I. Expression profiling of rice cultivars differing in their tolerance to long-term drought stress. Plant Mol. Biol 2009, 69, 133–153. [Google Scholar]
- Lenka, S.K.; Katiyar, A.; Chinnusamy, V.; Bansal, K.C. Comparative analysis of drought-responsive transcriptome in Indica rice genotypes with contrasting drought tolerance. Plant Biotechnol. J. 2010, voume number?. 1–13. [Google Scholar]
- Wang, D.; Pan, Y.; Zhao, X.; Zhu, L.; Fu, B.; Li, Z. Genome-wide temporal-spatial gene expression profiling of drought responsiveness in rice. BMC Genomics 2011, 12, 149–163. [Google Scholar]
- Zou, G.H.; Mei, H.W.; Liu, H.Y.; Liu, G.L.; Hu, S.P.; Yu, X.Q.; Li, M.S.; Wu, J.H.; Luo, L.J. Grain yield responses to moisture regimes in a rice population: association among traits and genetic markers. Theor. Appl. Genet 2005, 112, 106–113. [Google Scholar]
- Yue, B.; Xue, W.; Xiong, L.; Yu, X.; Luo, L.; Cui, K.; Jin, D.; Xing, Y.; Zhang, Q. Genetic basis of drought resistance at reproductive stage in rice: separation of drought tolerance from drought avoidance. Genetics 2006, 172, 1213–1228. [Google Scholar]
- Ding, X.; Li, X.; Xiong, L. Evaluation of near-isogenic lines for drought resistance QTL and fine mapping of a locus affecting flag leaf width, spikelet number, and root volume in rice. Theor. Appl. Genet 2011, 123, 815–826. [Google Scholar]
- Xiang, Y.; Tang, N.; Du, H.; Ye, H.; Xiong, L. Characterization of OsbZIP23 as a key player of the basic leucine zipper transcription factor family for conferring abscisic acid sensitivity and salinity and drought tolerance in rice. Plant Physiol 2008, 148, 1938–1952. [Google Scholar]
- Xiao, B.Z.; Chen, X.; Xiang, C.B.; Tang, N.; Zhang, Q.F.; Xiong, L.Z. Evaluation of seven function-known candidate genes for their effects on improving drought resistance of transgenic rice under field conditions. Mol. Plant 2009, 2, 73–83. [Google Scholar]
- Ding, X.; Hou, X.; Xie, K.; Xiong, L. Genome-wide identification of BURP domain-containing genes in rice reveals a gene family with diverse structures and responses to abiotic stresses. Planta 2009, 230, 149–163. [Google Scholar]
- Xiao, B.; Huang, Y.; Tang, N.; Xiong, L. Over-expression of a LEA gene in rice improves drought resistance under the field conditions. Theor. Appl. Genet 2007, 115, 35–46. [Google Scholar]
- Wang, L.; Xie, W.; Chen, Y.; Tang, W.; Yang, J.; Ye, R.; Liu, L.; Lin, Y.; Xu, C.; Xiao, J.; et al. A dynamic gene expression atlas covering the entire life cycle of rice. Plant J. 2010, 61, 752–766. [Google Scholar]
- Jain, M.; Nijhawan, A.; Tyagi, A.K.; Khurana, J.P. Validation of housekeeping genes as internal control for studying gene expression in rice by quantitative real-time PCR. Biochem. Biophys. Res. Commun 2006, 345, 646–651. [Google Scholar]
- Jain, M.; Nijhawan, A.; Arora, R.; Agarwal, P.; Ray, S.; Sharma, P.; Kapoor, S.; Tyagi, A.K.; Khurana, J.P. F-box proteins in rice: genome-wide analysis, classification, temporal and spatial gene expression during panicle and seed development, and regulation by light and abiotic stress. Plant Physiol 2007, 143, 1467–1483. [Google Scholar]
- Ouyang, S.Q.; Liu, Y.F.; Liu, P.; Lei, G.; He, S.J.; Ma, B.; Zhang, W.K.; Zhang, J.S.; Chen, S.Y. Receptor-like kinase OsSIK1 improves drought and salt stress tolerance in rice (Oryza sativa) plants. Plant J 2010, 62, 316–329. [Google Scholar]
- Zhou, J.; Wang, X.; Jiao, Y.; Qin, Y.; Liu, X.; He, K.; Chen, C.; Ma, L.; Wang, J.; Xiong, L.; et al. Global genome expression analysis of rice in response to drought and high-salinity stresses in shoot, flag leaf, and panicle. Plant Mol. Biol 2007, 63, 591–608. [Google Scholar]
- Hazen, S.P.; Pathan, M.S.; Sanchez, A.; Baxter, I.; Dunn, M.; Estes, B.; Chang, H.S.; Zhu, T.; Kreps, J.A.; Nguyen, H.T. Expression profiling of rice segregating for drought tolerance QTLs using a rice genome array. Funct. Integr. Genomics 2005, 5, 104–116. [Google Scholar]
- Robin, S.; Pathan, M.S.; Courtois, B.; Lafitte, R.; Carandang, S.; Lanceras, S.; Amante, M.; Nguyen, H.T.; Li, Z. Mapping osmotic adjustment in an advanced backcross inbred population of rice. Theor. Appl. Genet 2003, 107, 1288–1296. [Google Scholar]
- Xue, W.; Xing, Y.; Weng, X.; Zhao, Y.; Tang, W.; Wang, L.; Zhou, H.; Yu, S.; Xu, C.; Li, X.; et al. Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice. Nat. Genet 2008, 40, 761–767. [Google Scholar]
- Schubert, V. SMC proteins and their multiple functions in higher plants. Cytogenet. Genome Res 2009, 124, 202–214. [Google Scholar]
- Affymetrix standard protocols. Available online: http://www.affymetrix.com/products/arrays/specific/rice.affx accessed on 17 February 2013.
- R platform. Available online: http://www.R-project.org accessed on 17 February 2013.
- Bolstad, B.M.; Irizarry, R.A.; Astrand, M.; Speed, T.P. A comparison of normalization methods for high-density oligonucleotide array data based on variance and bias. Bioinformatics 2003, 19, 185–193. [Google Scholar]
- Irizarry, R.A.; Hobbs, B.; Collin, F.; Beazer-Barclay, Y.D.; Antonellis, K.J.; Scherf, U.; Speed, T.P. Exploration, normalization, and summaries of high-density oligonucleotide array probe level data. Biostatistics 2003, 4, 249–264. [Google Scholar]
- Gautier, L.; Cope, L.; Bolstad, B.M.; Irizarry, R.A. affy: analysis of Affymetrix GeneChip data at the probe level. Bioinformatics 2004, 20, 307–315. [Google Scholar]
- Gentleman, R.C.; Carey, V.J.; Bates, D.M.; Bolstad, B.; Dettling, M.; Dudoit, S.; Ellis, B.; Gautier, L.; Ge, Y.; Gentry, J.; et al. Bioconductor: open software development for computational biology and bioinformatics. Genome Biol 2004, 5, R80. [Google Scholar]
- Breitling, R.; Armengaud, P.; Amtmann, A.; Herzyk, P. Rank products: A simple, yet powerful, new method to detect differentially regulated genes in replicated microarray experiments. FEBS Lett 2004, 573, 83–92. [Google Scholar]
- Alexa, A.; Rahnenfuhrer, J.; Lengauer, T. Improved scoring of functional groups from gene expression data by decorrelating GO graph structure. Bioinformatics 2006, 22, 1600–1607. [Google Scholar]
- Wang, J.; Yu, H.; Xie, W.; Xing, Y.; Yu, S.; Xu, C.; Li, X.; Xiao, J.; Zhang, Q. A global analysis of QTLs for expression variations in rice shoots at the early seedling stage. Plant J 2010, 63, 1063–1074. [Google Scholar]
- Hartweck, L.M.; Olszewski, N.E. Rice GIBBERELLIN INSENSITIVE DWARF1 is a gibberellin receptor that illuminates and raises questions about GA signaling. Plant Cell 2006, 18, 278–282. [Google Scholar]
- Cao, W.H.; Liu, J.; He, X.J.; Mu, R.L.; Zhou, H.L.; Chen, S.Y.; Zhang, J.S. Modulation of ethylene responses affects plant salt-stress responses. Plant Physiol 2007, 143, 707–719. [Google Scholar]
- Liang, D.; Wu, C.; Li, C.; Xu, C.; Zhang, J.; Kilian, A.; Li, X.; Zhang, Q.; Xiong, L. Establishment of a patterned GAL4-VP16 transactivation system for discovering gene function in rice. Plant J 2006, 46, 1059–1072. [Google Scholar]
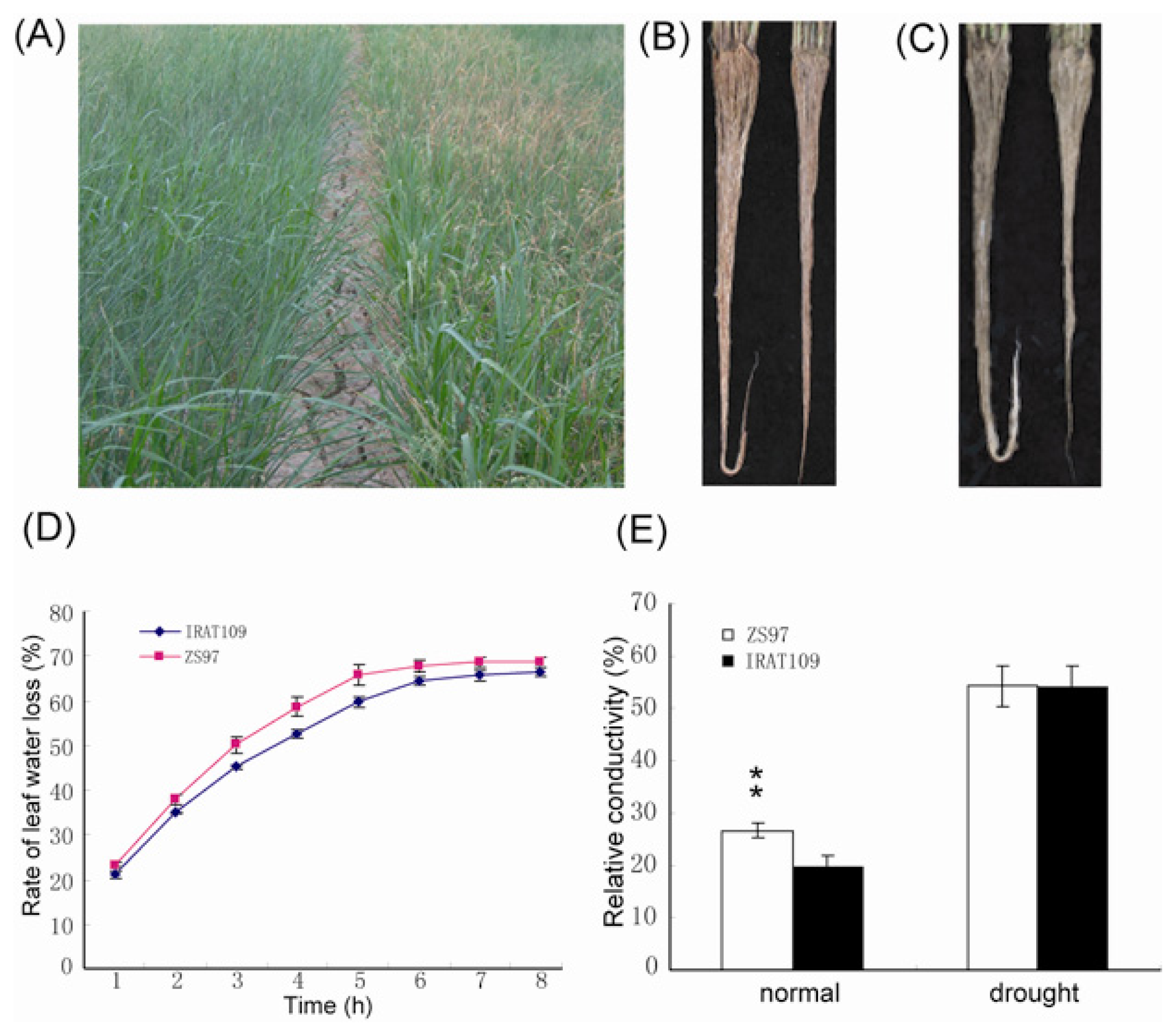
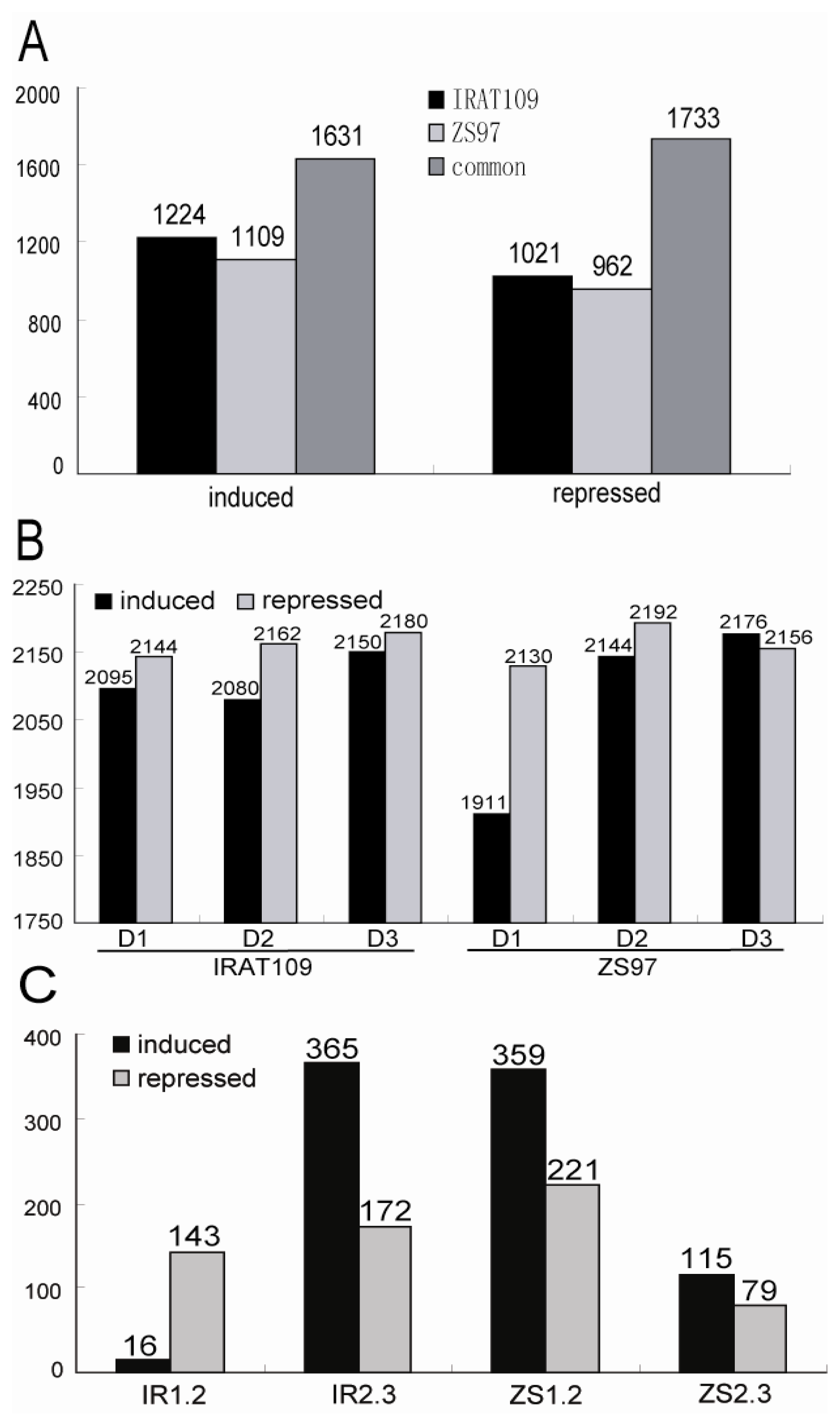

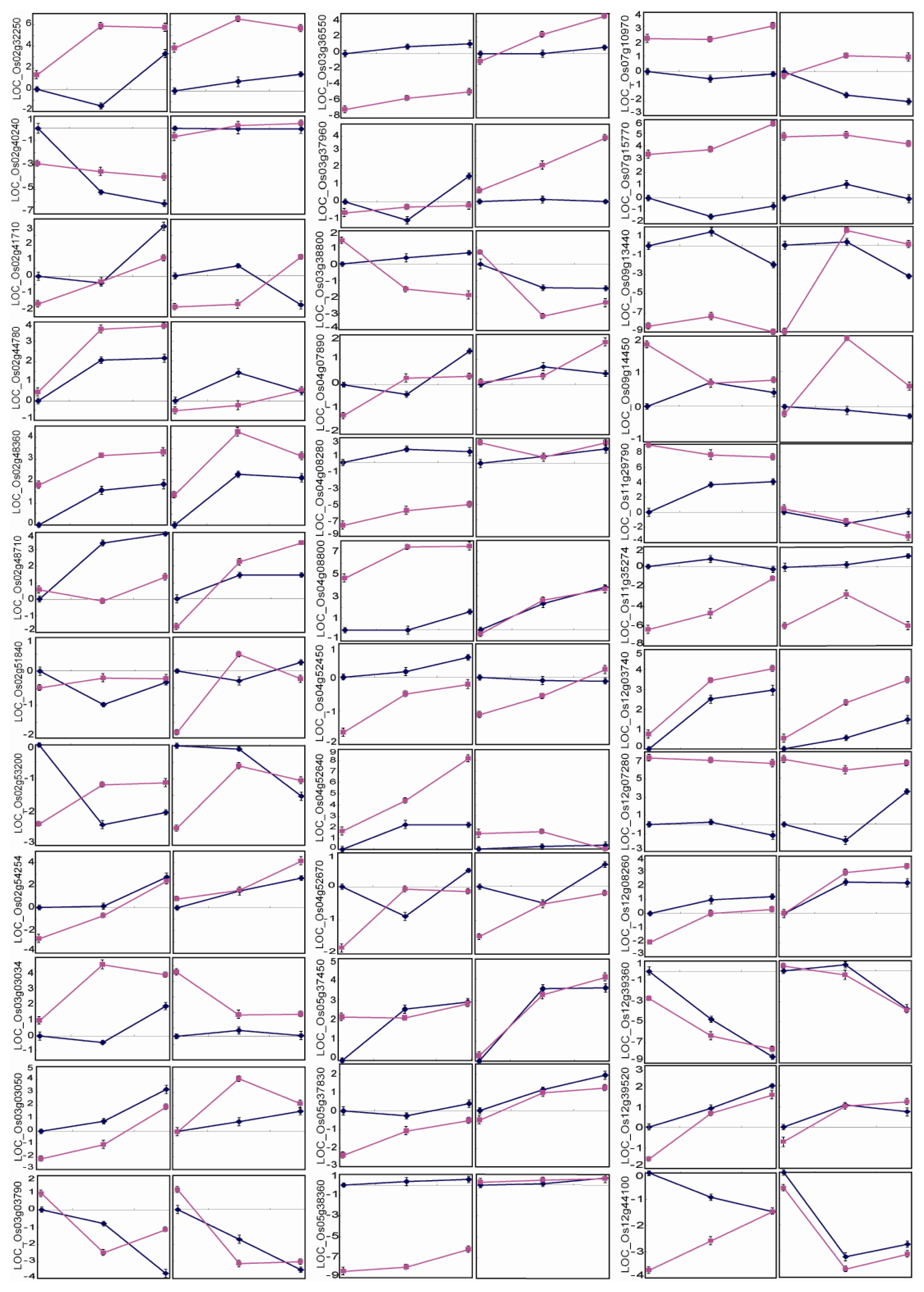

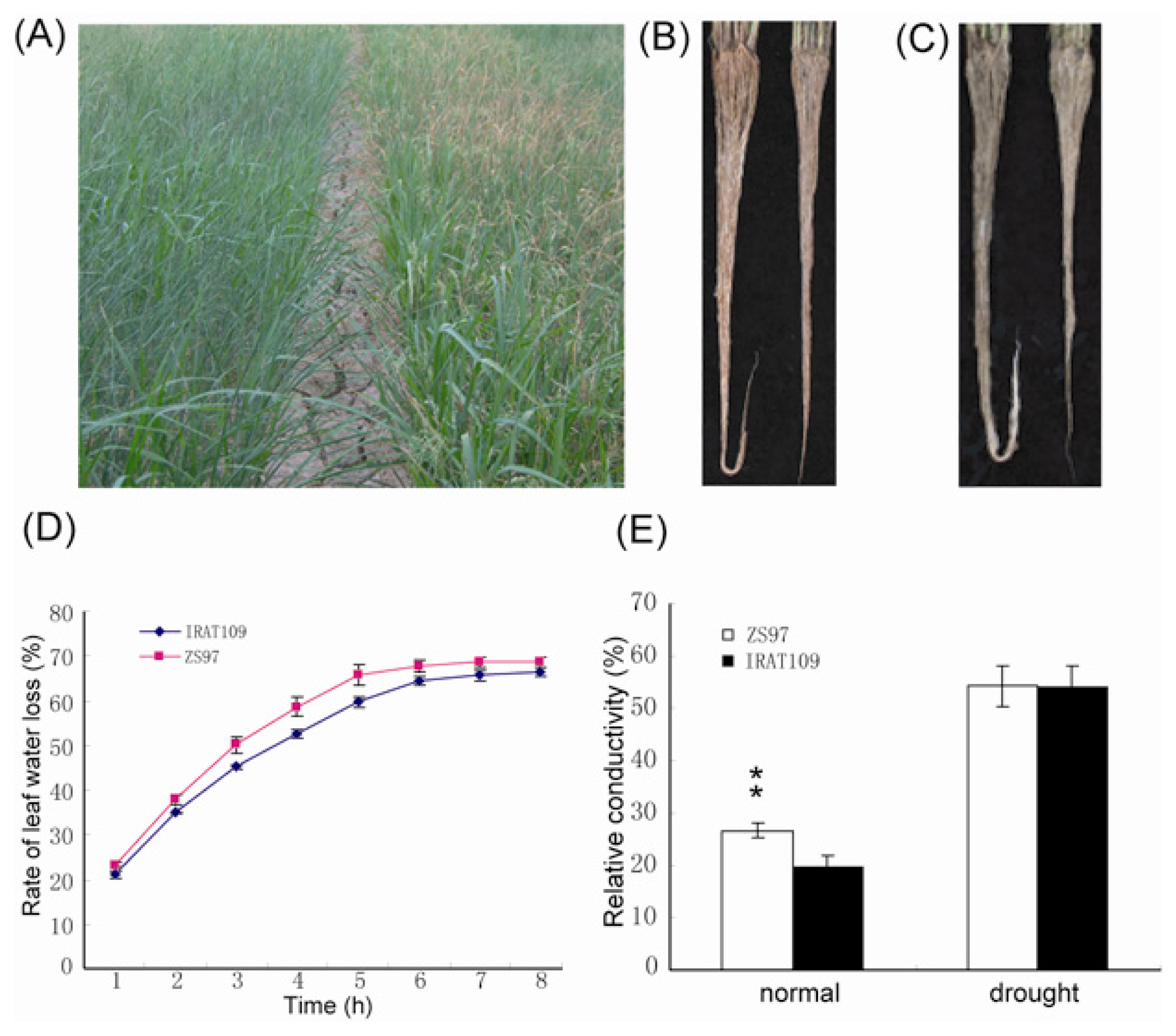{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Normal growth | Drought stress | Relative value b | |||
|---|---|---|---|---|---|---|
| ZS97 | IRAT109 | ZS97 | IRAT109 | ZS97 | IRAT109 | |
| PH | 87.34 ± 6.96 | 103.95 ± 7.58 **a | 80.81 ± 6.63 | 95.33 ± 6.63 ** | 0.926 | 0.917 |
| GY | 41.72 ± 5.41 * | 38.45 ± 7.11 | 19.83 ± 7.4 | 25.55 ± 10.14 ** | 0.475 | 0.662 |
| TPP | 31.57 ± 2.27 ** | 24.85 ± 2.13 | 23.84 ± 8.12 ** | 13.13 ± 5.94 | 0.753 | 0.526 |
| SPP | 93.34 ± 17.81 | 114.18 ± 24.94 ** | 75.44 ± 12.57 ** | 72.14 ± 16.51 | 0.808 | 0.631 |
| SF | 0.91 ± 0.06 ** | 0.67 ± 0.08 | 0.47 ± 0.10 | 0.46 ± 0.20 | 0.516 | 0.687 |
| KGW | 23.48 ± 2.02 | 32.28 ± 2.14 ** | 19.69 ± 2.27 | 31.26 ± 3.84 ** | 0.838 | 0.969 |
| RV | 36.62 ± 9.43 | 60.51 ± 10.27 ** | 20.82 ± 8.43 | 31.56 ± 9.15 ** | 0.568 | 0.522 |
| RVT | 1.16 ± 0.37 | 2.43 ± 0.57 ** | 0.86 ± 0.27 | 2.38 ± 0.50 ** | 0.741 | 0.979 |
| GO.ID | Term | IRAT109 | ZS97 | |||||
|---|---|---|---|---|---|---|---|---|
| Level a | Significant b | Expected c | Classic d | Significant b | Expected c | Classic d | ||
| Up | ||||||||
| GO:0006950 | response to stress | 3 | 233 | 180.79 | 9.60 × 10−6 | 250 | 180.61 | 8.50 × 10−9 |
| GO:0009605 | response to external stimulus | 3 | 69 | 43.5 | 8.10 ×10−5 | 80 | 43.46 | 5.90×10−8 |
| GO:0005975 | carbohydrate metabolic process | 4 | 111 | 77.88 | 7.00 ×10−5 | 131 | 77.8 | 1.00 × 10−9 |
| GO:0009991 | response to extracellular stimulus | 4 | 16 | 7.84 | 0.00475 | 21 | 7.83 | 2.60 × 10−5 |
| GO:0000271 | polysaccharide biosynthetic process | 6 | 10 | 4.3 | 0.00949 | 12 | 4.29 | 0.00093 |
| GO:0042221 | response to chemical stimulus | 3 | - | - | - | 32 | 19.36 | 0.00338 |
| GO:0006725 | cellular aromatic compound metabolic process | 4 | - | - | - | 24 | 12.88 | 0.00225 |
| GO:0017144 | drug metabolic process | 4 | - | - | - | 10 | 3.54 | 0.00224 |
| GO:0016051 | carbohydrate biosynthetic process | 5 | - | - | - | 19 | 9.49 | 0.00275 |
| GO:0016999 | antibiotic metabolic process | 5 | - | - | - | 10 | 3.54 | 0.00224 |
| GO:0044270 | nitrogen compound catabolic process | 5 | - | - | - | 7 | 2.18 | 0.00483 |
| GO:0017000 | antibiotic biosynthetic process | 6 | - | - | - | 8 | 3.01 | 0.00889 |
| GO:0034637 | cellular carbohydrate biosynthetic process | 6 | - | - | - | 13 | 5.65 | 0.00369 |
| GO:0009250 | glucan biosynthetic process | 8 | - | - | - | 10 | 3.84 | 0.0042 |
| GO:0016787 | hydrolase activity | 3 | - | - | - | 290 | 246.07 | 0.00093 |
| GO:0016491 | oxidoreductase activity | 3 | - | - | - | 53 | 37.37 | 0.0065 |
| GO:0006139 | nucleobase, nucleoside, nucleotide and nucleic acid metabolic process | 4 | 286 | 250.98 | 0.00506 | - | - | - |
| GO:0022402 | cell cycle process | 4 | 6 | 1.36 | 0.00153 | - | - | - |
| GO:0044262 | cellular carbohydrate metabolic process | 5 | 24 | 13.95 | 0.00629 | - | - | - |
| GO:0016311 | dephosphorylation | 6 | 13 | 5.81 | 0.0047 | - | - | - |
| GO:0044042 | glucan metabolic process | 6 | 13 | 6.11 | 0.00727 | - | - | - |
| GO:0006073 | cellular glucan metabolic process | 7 | 13 | 6.11 | 0.00727 | - | - | - |
| GO:0006350 | transcription | 7 | 187 | 145.2 | 9.70 × 10−5 | - | - | - |
| GO:0006470 | protein amino acid dephosphorylation | 9 | 13 | 5.81 | 0.0047 | - | - | - |
| GO:0045735 | nutrient reservoir activity | 2 | 11 | 4.78 | 0.00753 | - | - | - |
| GO:0003676 | nucleic acid binding | 3 | 325 | 286.7 | 0.00509 | - | - | - |
| Down
| ||||||||
| GO:0050896 | response to stimulus | 2 | 608 | 465.77 | 2.90 × 10−17 | 583 | 447.76 | 2.80 × 10−16 |
| GO:0065007 | biological regulation | 2 | 441 | 378 | 4.30 × 10−5 | 416 | 363.39 | 0.00042 |
| GO:0022414 | reproductive process | 3 | 82 | 52.03 | 1.90 ×10−5 | 71 | 50.02 | 0.0015 |
| GO:0048856 | anatomical structure development | 3 | 73 | 46.77 | 7.40 × 10−5 | 66 | 44.96 | 0.00092 |
| GO:0006793 | phosphorus metabolic process | 4 | 252 | 158.57 | 3.70 × 10−5 | 231 | 152.44 | 1.10 ×10−11 |
| GO:0009908 | flower development | 6 | 66 | 40.61 | 4.80 × 10−5 | 60 | 39.04 | 0.00049 |
| GO:0016310 | phosphorylation | 6 | 242 | 147.35 | 2.90 × 10−16 | 221 | 141.65 | 1.90 × 10−12 |
| GO:0043687 | post-translational protein modification | 8 | 278 | 194.01 | 3.50 × 10−11 | 255 | 186.51 | 2.70 ×10−8 |
| GO:0006468 | protein amino acid phosphorylation | 9 | 233 | 136.72 | 1.40 × 10−17 | 215 | 131.44 | 3.00 × 10−14 |
| GO:0009653 | anatomical structure morphogenesis | 4 | 50 | 26.71 | 9.20 × 10−6 | 42 | 25.68 | 0.00096 |
| GO:0030154 | cell differentiation | 4 | 83 | 49.94 | 2.10 × 10−6 | 76 | 48.01 | 3.40 ×10−5 |
| GO:0048608 | reproductive structure development | 5 | 71 | 43.49 | 2.10 × 10−5 | 64 | 41.81 | 0.00036 |
| GO:0006464 | protein modification process | 7 | 340 | 266.3 | 1.50 × 10−7 | 307 | 256 | 0.00014 |
| GO:0006725 | cellular aromatic compound metabolic process | 4 | - | - | - | 27 | 16.32 | 0.00619 |
| GO:0006979 | response to oxidative stress | 4 | - | - | - | 27 | 15.65 | 0.00347 |
| GO:0009991 | response to extracellular stimulus | 4 | - | - | - | 18 | 9.93 | 0.00921 |
| GO:0046164 | alcohol catabolic process | 4 | - | - | - | 21 | 12.22 | 0.00959 |
| GO:0009225 | nucleotide-sugar metabolic process | 5 | - | - | - | 18 | 8.11 | 0.00094 |
| GO:0019318 | hexose metabolic process | 6 | - | - | - | 24 | 13.75 | 0.00482 |
| GO:0046365 | monosaccharide catabolic process | 6 | - | - | - | 21 | 11.74 | 0.00608 |
| GO:0007155 | cell adhesion | 3 | 9 | 3.67 | 0.00876 | - | - | - |
| GO:0006810 | transport | 4 | 222 | 188.16 | 0.00338 | - | - | - |
| GO:0006811 | ion transport | 5 | 63 | 40.11 | 0.00019 | - | - | - |
| GO:0015837 | amine transport | 5 | 18 | 8.54 | 0.00171 | - | - | - |
| GO:0015849 | organic acid transport | 5 | 18 | 8.74 | 0.00224 | - | - | - |
| GO:0055085 | transmembrane transport | 5 | 31 | 19.16 | 0.00487 | - | - | - |
| GO:0006812 | cation transport | 6 | 49 | 34.65 | 0.00803 | - | - | - |
| GO:0006820 | anion transport | 6 | 11 | 4.57 | 0.00452 | - | - | - |
| GO:0006855 | multidrug transport | 6 | 10 | 4.37 | 0.00966 | - | - | - |
| GO:0009309 | amine biosynthetic process | 6 | 24 | 14.3 | 0.00781 | - | - | - |
| GO:0006865 | amino acid transport | 7 | 18 | 8.54 | 0.00171 | - | - | - |
| GO:0015698 | inorganic anion transport | 7 | 10 | 3.08 | 0.00057 | - | - | - |
| GO:0008652 | cellular amino acid biosynthetic process | 8 | 22 | 12.81 | 0.00809 | - | - | - |
| GO:0009064 | glutamine family amino acid metabolic process | 8 | 10 | 3.97 | 0.00475 | - | - | - |
| GO:0009069 | serine family amino acid metabolic process | 8 | 11 | 4.96 | 0.00876 | - | - | - |
| GO:0004553 | hydrolase activity, hydrolyzing O-glycosyl compounds | 5 | 47 | 32.8 | 0.00784 | - | - | - |
| TIGR Locus | Annotation (TIGR version 6.1) | Normal a | Drought b | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ZS97 | IRAT109 | NIL-ZS | NIL-IR | ZS97 | IRAT109 | NIL-ZS | NIL-IR | C1 c | C2 d | ||
| LOC_Os02g32250 | retrotransposon protein | − | + | − | + | ↑ | ↑ | ↑ | ↑ | 4 | a |
| LOC_Os02g40240 | receptor kinase | + | − | ↓ | ↓ | ↑ | 1 | d | |||
| LOC_Os02g41710 | cyclic nucleotide-gated ion channel | + | − | + | − | ↑ | ↑ | ↓ | ↑ | 2 | d |
| LOC_Os02g44780 | polyprenyl synthetase | ↑ | ↑ | ↑ | ↑ | 4 | a | ||||
| LOC_Os02g48360 | pyrophosphate-fructose 6-phosphate 1-phosphotransferase subunit alpha | − | + | − | + | ↑ | ↑ | ↑ | ↑ | 4 | a |
| LOC_Os02g48710 | expressed protein | + | − | ↑ | ↑ | ↑ | 3 | b | |||
| LOC_Os02g51840 | expressed protein | + | − | ↑ | 3 | d | |||||
| LOC_Os02g53200 | glucan endo-1,3-beta-glucosidase precursor | + | − | + | − | ↓ | ↑ | ↓ | ↑ | 4 | c |
| LOC_Os02g54254 | saccharopine dehydrogenase | + | − | ↑ | ↑ | ↑ | ↑ | 4 | a | ||
| LOC_Os03g03034 | flavonol synthase/flavanone 3-hydroxylase | − | + | ↑ | ↑ | ↓ | 1 | d | |||
| LOC_Os03g03050 | expressed protein | + | − | ↑ | ↑ | ↑ | ↑ | 4 | a | ||
| LOC_Os03g03790 | AMP-binding domain containing protein | − | + | ↓ | ↓ | ↓ | ↓ | 4 | a | ||
| LOC_Os03g36550 | transposon protein | + | − | ↑ | ↑ | ↑ | 2 | d | |||
| LOC_Os03g37960 | acyl CoA binding protein | ↑ | ↑ | 1 | c | ||||||
| LOC_Os03g38800 | AAA family ATPase | − | + | ↓ | ↓ | ↓ | 2 | b | |||
| LOC_Os04g07890 | AGAP002737-PA | + | − | ↑ | ↑ | ↑ | 2 | d | |||
| LOC_Os04g08280 | retrotransposon protein,Ty3-gypsy subclass | + | − | − | + | ↑ | ↑ | ↑ | ↓ | 3 | d |
| LOC_Os04g08800 | expressed protein | − | + | ↑ | ↑ | ↑ | ↑ | 4 | a | ||
| LOC_Os04g52450 | aminotransferase | + | − | + | − | ↑ | ↑ | 4 | c | ||
| LOC_Os04g52640 | SHR5-receptor-like kinase | − | + | − | + | ↑ | ↑ | ↓ | 1 | d | |
| LOC_Os04g52670 | OsSAUR21 - Auxin-responsive SAUR gene family member | + | − | + | − | ↑ | ↑ | 4 | c | ||
| LOC_Os05g37450 | starch binding domain containing protein | − | + | ↑ | ↑ | ↑ | 3 | b | |||
| LOC_Os05g37830 | expressed protein | + | − | ↑ | ↑ | ↑ | 2 | b | |||
| LOC_Os05g38360 | DHHC zinc finger domain containing protein | + | − | ↑ | 3 | b | |||||
| LOC_Os07g10970 | leucine zipper protein-like | − | + | ↓ | ↑ | 1 | c | ||||
| LOC_Os07g15770 | CCT motif family protein | − | + | − | + | ↓ | ↑ | ↑ | 1 | c | |
| LOC_Os09g13440 | expressed protein | + | − | + | − | ↑↓ | ↓ | ↑ | 1 | c | |
| LOC_Os09g14450 | pollen signaling protein with adenylyl cyclase activity | − | + | ↓ | ↑ | 3 | c | ||||
| LOC_Os11g29790 | receptor kinase | − | + | ↑ | ↓ | ↓ | ↓ | 2 | b | ||
| LOC_Os11g35274 | protein kinase domain containing protein | + | − | + | − | ↑ | ↑ | ↑ | 2 | b | |
| LOC_Os12g03740 | OsFBX438 - F-box domain containing protein | ↑ | ↑ | ↑ | ↑ | 4 | a | ||||
| LOC_Os12g07280 | ZOS12-02 - C2H2 zinc finger protein | − | + | − | + | ↓ | ↓↑ | ↓ | 1 | c | |
| LOC_Os12g08260 | dehydrogenase E1 component domain containing protein | + | − | ↑ | ↑ | ↑ | ↑ | 4 | a | ||
| LOC_Os12g39360 | aspartic proteinase nepenthesin precursor | + | − | ↓ | ↓ | ↓ | ↓ | 4 | a | ||
| LOC_Os12g39520 | OsFBDUF66 - F-box and DUF domain containing protein | + | − | ↑ | ↑ | ↑ | ↑ | 4 | a | ||
| LOC_Os12g44100 | peptide transporter PTR2 | + | − | ↓ | ↑ | ↓ | ↓ | 3 | b | ||
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ding, X.; Li, X.; Xiong, L. Insight into Differential Responses of Upland and Paddy Rice to Drought Stress by Comparative Expression Profiling Analysis. Int. J. Mol. Sci. 2013, 14, 5214-5238. https://doi.org/10.3390/ijms14035214
Ding X, Li X, Xiong L. Insight into Differential Responses of Upland and Paddy Rice to Drought Stress by Comparative Expression Profiling Analysis. International Journal of Molecular Sciences. 2013; 14(3):5214-5238. https://doi.org/10.3390/ijms14035214
Chicago/Turabian StyleDing, Xipeng, Xiaokai Li, and Lizhong Xiong. 2013. "Insight into Differential Responses of Upland and Paddy Rice to Drought Stress by Comparative Expression Profiling Analysis" International Journal of Molecular Sciences 14, no. 3: 5214-5238. https://doi.org/10.3390/ijms14035214