Identification of Novel Pepper Genes Involved in Bax- or INF1-Mediated Cell Death Responses by High-Throughput Virus-Induced Gene Silencing


Abstract
:1. Introduction
2. Results and Discussion
2.1. Silencing of Pathogen-Induced Pepper ESTs in N. benthamiana Plants
2.2. Screening of Pepper Genes Coupled with Bax- or INF1-Induced Cell Death Responses
2.3. Silencing of the Putative Calcium-Binding Protein CML30-Like and Cys-3-His Zinc Finger Protein Suppressed Bax-Induced Cell Dath, but Not INF1-Induced Cell Death
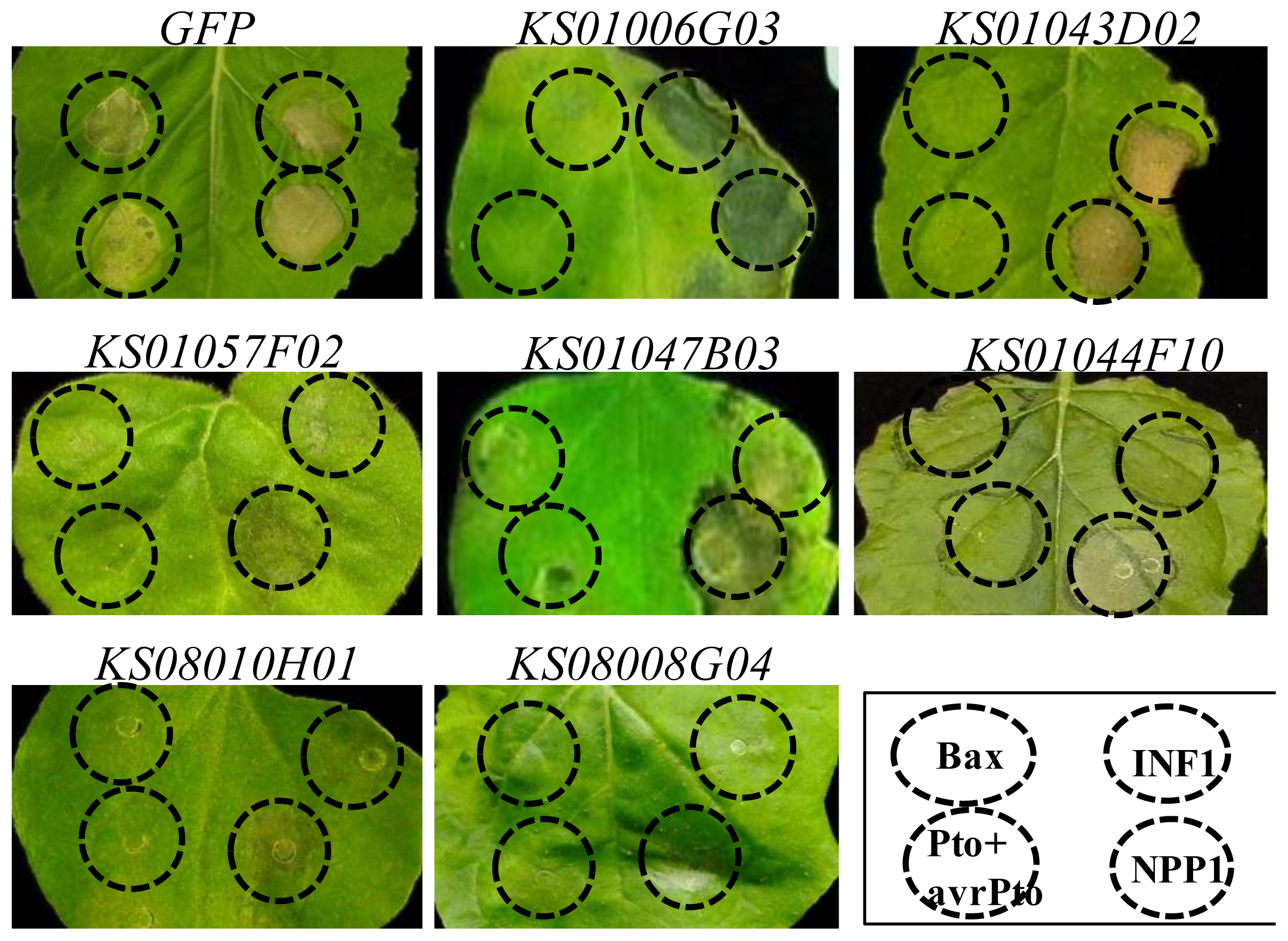
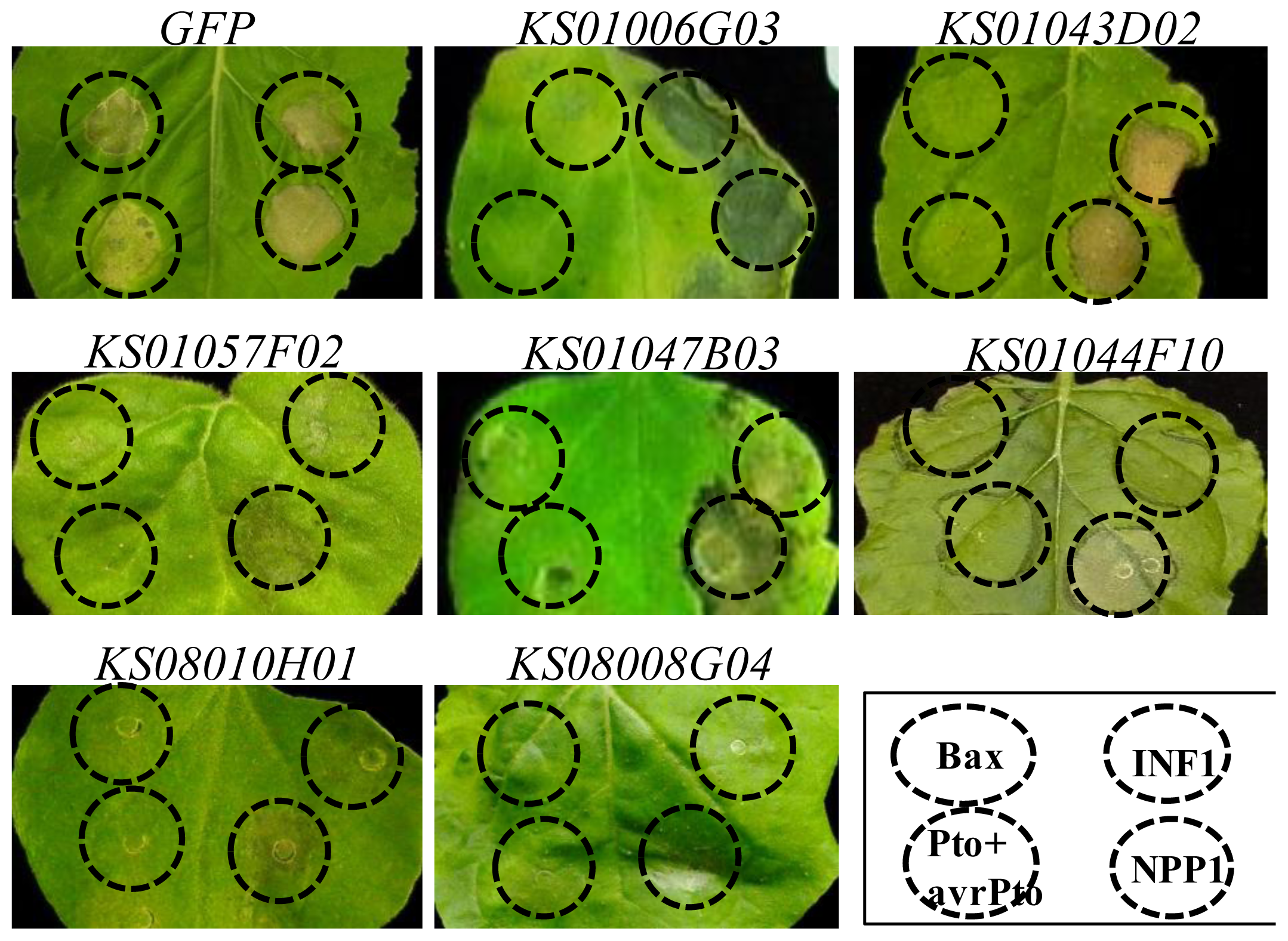
2.4. Bax- and INF1-Induced Cell Death Were Suppressed in N. benthamiana Plants in Which One of Five Pepper Genes Were Silenced
3. Experimental Section
3.1. Plant Materials and VIGS
3.2. Agrobacterium-Mediated Transient Expression
3.3. Non-Host Pathogen-Induced Pepper ESTs
3.4. RT-PCR Analysis
4. Conclusions
Supplementary Information
ijms-14-22782-s001.pdf



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EST ID | BLASTX * | VIGSed phenotypes |
|---|---|---|
| Catalytic activity/ligase activity | ||
| KS01017F01 | Q8RU74_LYCES Dehydroquinate synthase (EC 4.6.1.3) | Stunted |
| KS26035G04 | Q5Z8K3_ORYSA Putative ZEITLUPE | Stunted |
| Catalytic activity/oxidoreductase activity | ||
| KS01002H05 | Q6S5A3_TROMA Sterol delta-7 reductase | Dead |
| KS13009F01 | Q9SVA9_ARATH Cytochrome P450-like protein | Leaf necrotic spot |
| Catalytic activity/transferase activity | ||
| KS13055B07 | Q5DKV0_TOBAC Adenosine kinase isoform 1T | Wrinkled leaf |
| Catalytic activity/hydrolase activity | ||
| KS23052B07 | chitinase/lysozyme [Nicotiana tabacum] | Pale green |
| KS08010H01 | probable pectate lyase P56-like [S. lycopersicum] | Stunted & wrinkled leaf |
| KS01052C01 | Q8H272_LYCES Metacaspase 1 (Fragment) | Wrinkled leaf |
| Binding/ion binding | ||
| KS12078F02 | calmodulin [Capsicum annuum] | Stunted |
| KS01006G03 | probable calcium-binding protein CML30-like [S. lycopersicum] | Stunted |
| Binding/protein binding | ||
| KS01047E07 | O04133_SOYBN SRC2 | Stunted |
| Binding/nucleic acid binding | ||
| KS01060G11 | Q93XV7_9ROSI Hydroxypyruvate reductase (EC 1.1.1.29) | Small leaf size |
| KS12047A04 | Q9XI07_ARATH F8K7.13 protein (Putative transcriptional regulatory protein) | Weak dwarf |
| Binding/others | ||
| KS01044F10 | BIP5_TOBAC Luminal binding protein 5 precursor (BiP 5) | Stunted |
| KS01057F02 | mannose-binding lectin [C. annuum] | Stunted |
| KS14050G06 | Q41424_SOLTU Chlorophyll a/b binding protein | Pale green |
| Structural molecule activity | ||
| KS10105B11 | Q9SCM3_ARATH 40S ribosomal protein S2 homolog (At3g57490) | Stunted & curling leaf |
| KS08008G04 | 60S ribosomal protein L10 [S. melongena] | Stunted & curling leaf |
| Transporter activity | ||
| KS24014C12 | Q93Y42_ARATH Coatomer delta subunit (Delta-coat protein) | Dead |
| KS01047B03 | Q9XF20_CAPCH Acyl carrier protein | Stunted |
| KS08017G06 | O80774_ARATH Hypothetical protein At2g34250 | Stunted & curling leaf |
| Transcription regulator activity | ||
| KS01043D02 | Q9XEE6_ARATH Hypothetical Cys-3-His zinc finger | Stunted |
| KS09041E04 | Q710C3_SPIOL Sigma factor (Fragment) | Pale green |
| KS09088A04 | Q7Y039_LYCES MADS-box protein 5 | Abnormal leaf |
| KS12065G09 | Q8RXK4_ARATH Hypothetical protein At4g38900 | Stunted |
| Unknown | ||
| KS20002B10 | later embryo abundant protein [Prunus dulcis] | Stunted |
| KS01047F02 | Q9C9T7_ARATH Hypothetical protein F25P22.17 | Stunted |
| KS01056A09 | No_hits | Complete dead |
| KS20014C01 | No_hits | Leaf local dead |
| KS07002E11 | AAD25142.1 (Hypothetical protein At3g24506) | Pale green |
| KS07012E06 | Q9FJZ7_ARATH Arabidopsis thaliana genomic DNA, chromosome 5, | Chlorosis |
| KS19065G03 | Q9C9V9_ARATH Putative golgi transport complex protein; 67058-70172 | Stunted & curling leaf |
| EST ID | BLASTX * | E-value | Phenotypes | ||
|---|---|---|---|---|---|
| Morphology | HR | ||||
| Bax | INF | ||||
| KS01044F10 | glucose-regulated protein 78 (BiP), partial [S. lycopersicum] | 2.0 × 10−87 | stunted | X | X |
| KS01047B03 | Acyl carrier protein [C. chinense] | 3.0 × 10−58 | stunted | X | X |
| KS08008G04 | 60S ribosomal protein L10 [S. melongena] | 7.0 × 10−121 | stunted & wrinkled leaf | X | X |
| KS08010H01 | probable pectate lyase P56-like [S. lycopersicum] | 8.0 × 10−77 | stunted & wrinkled leaf | X | X |
| KS01057F02 | mannose-binding lectin [C. annuum] | 1.0 × 10−77 | stunted | X | X |
| KS01006G03 | probable calcium-binding protein CML30-like [S. lycopersicum] | 1.0 × 10−77 | stunted | X | Δ |
| KS01043D02 | Hypothetical Cys-3-His zinc finger protein [A. thaliana] | 1.0 × 10−125 | stunted | X | O |
Acknowledgments
Conflicts of Interest
References
- Baulcombe, D.C. Fast forward genetics based on virus-induced gene silencing. Curr. Opin. Plant Biol 1999, 2, 109–113. [Google Scholar]
- Ruiz, M.T.; Voinnet, O.; Baulcombe, D.C. Initiation and maintenance of virus-induced gene silencing. Plant Cell 1998, 10, 937–946. [Google Scholar]
- Terauchi, R.; Bin Nasir, K.H.; Ito, A.; Saitoh, H.; Berberich, T.; Takahashi, Y. High-throughput functional screening of plant and pathogen genes in planta. Plant Biotechnol 2005, 22, 455–459. [Google Scholar]
- Anand, A.; Vaghchhipawala, Z.; Ryu, C.M.; Kang, L.; Wang, K.; Del-Pozo, O.; Martin, G.B.; Mysore, K.S. Identification and characterization of plant genes involved in Agrobacterium-mediated plant transformation by virus-induced silencing. Mol. Plant Microbe Interact 2007, 20, 41–52. [Google Scholar]
- Senthil-Kumar, M.; Udayakumar, M. High-throughput virus-induced gene-silencing approach to assess the functional relevance of a moisture stress-induced cDNA homologous to lea4. J. Exp. Bot 2006, 57, 2291–2302. [Google Scholar]
- Lu, R.; Malcuit, I.; Moffett, P.; Ruiz, M.T.; Peart, J.; Wu, A.J.; Rathjen, J.P.; Bendahmane, A.; Day, L.; Baulcombe, D.C. High throughput virus-induced gene silencing implicates heat shock protein 90 in plant disease resistance. EMBO J 2003, 22, 5690–5699. [Google Scholar]
- Bae, C.; Kim, S.M.; Lee, D.J.; Choi, D. Multiple classes of immune-related proteases associated with the cell death response in pepper plants. PLoS One 2013, 8, e63533. [Google Scholar]
- Senthil-Kumar, M.; Hema, R.; Anand, A.; Kang, L.; Udayakumar, M.; Mysore, K.S. A systematic study to determine the extent of gene silencing in Nicotiana benthamiana and other Solanaceae species when heterologous gene sequences are used for virus-induced gene silencing. New Phytol 2007, 176, 782–791. [Google Scholar]
- Greenberg, J.T.; Yao, N. The role and regulation of programmed cell death in plant-pathogen interactions. Cell. Microbiol 2004, 6, 201–211. [Google Scholar]
- Lam, E. Controlled cell death, plant survival and development. Nat. Rev. Mol. Cell Biol 2004, 5, 305–315. [Google Scholar]
- Lacomme, C.; Santa Cruz, S. Bax-induced cell death in tobacco is similar to the hypersensitive response. Proc. Natl. Acad. Sci. USA 1999, 96, 7956–7961. [Google Scholar]
- Kawai-Yamada, M.; Jin, L.; Yoshinaga, K.; Hirata, A.; Uchimiya, H. Mammalian Bax-induced plant cell death can be down-regulated by overexpression of Arabidopsis Bax Inhibitor-1 (AtBI-1). Proc. Natl. Acad. Sci. USA 2001, 98, 12295–12300. [Google Scholar]
- Matsumura, H.; Nirasawa, S.; Kiba, A.; Urasaki, N.; Saitoh, H.; Ito, M.; Kawai-Yamada, M.; Uchimiya, H.; Terauchi, R. Overexpression of Bax inhibitor suppresses the fungal elicitor-induced cell death in rice (Oryza sativa L.) cells. Plant J 2003, 33, 425–434. [Google Scholar]
- Kamoun, S.; van West, P.; Vleeshouwers, V.G.; de Groot, K.E.; Govers, F. Resistance of nicotiana benthamiana to phytophthora infestans is mediated by the recognition of the elicitor protein INF1. Plant Cell 1998, 10, 1413–1426. [Google Scholar]
- Kamoun, S.; Young, M.; Glascok, C.; Tyler, B.M. Extracellular protein elicitors from Phytophthora: Host-specificity and induction of resistance to fungal and bacterial phytopathogens. Mol. Plant Microbe Interact 1993, 10, 13–20. [Google Scholar]
- Kanzaki, H.; Saitoh, H.; Takahashi, Y.; Berberich, T.; Ito, A.; Kamoun, S.; Terauchi, R. NbLRK1, a lectin-like receptor kinase protein of Nicotiana benthamiana, interacts with Phytophthora infestans INF1 elicitin and mediates INF1-induced cell death. Planta 2008, 228, 977–987. [Google Scholar]
- Yoshioka, H.; Numata, N.; Nakajima, K.; Katou, S.; Kawakita, K.; Rowland, O.; Jones, J.D.G.; Doke, N. Nicotiana benthamiana gp91phox homologs NbrbohA and NbrbohB participate in H2O2 accumulation and resistance to Phytophthora infestans. Plant Cell 2003, 15, 706–718. [Google Scholar]
- Kanzaki, H.; Saitoh, H.; Ito, A.; Fujisawa, S.; Kamoun, S.; Katou, S.; Yoshioka, H.; Terauchi, R. Cytosolic HSP90 and HSP70 are essential components of INF1-mediated hypersensitive response and non-host resistance to pseudomonas cichorii in Nicotiana benthamiana. Mol. Plant Pathol 2003, 4, 383–391. [Google Scholar]
- Lee, S.; Kim, S.Y.; Chung, E.; Joung, Y.H.; Pai, H.S.; Hur, C.G.; Choi, D. EST and microarray analyses of pathogen-responsive genes in hot pepper (Capsicum annuum L.) non-host resistance against soybean pustule pathogen (Xanthomonas axonopodis pv. glycines). Funct. Integr. Genomics 2004, 4, 196–205. [Google Scholar]
- Liu, Y.; Schiff, M.; Dinesh-Kumar, S.P. Virus-induced gene silencing in tomato. Plant J 2002, 31, 777–786. [Google Scholar]
- Chung, E.; Seong, E.; Kim, Y.C.; Chung, E.J.; Oh, S.K.; Lee, S.; Park, J.M.; Joung, Y.H.; Choi, D. A method of high frequency virus-induced gene silencing in chili pepper (Capsicum annuum L. cv Bukang). Mol. Cells 2004, 17, 377–380. [Google Scholar]
- Fellbrich, G.; Romanski, A.; Varet, A.; Blume, B.; Brunner, F.; Engelhardt, S.; Felix, G.; Kemmerling, B.; Krzymowska, M.; Nürnberger, T. NPP1 a Phytophthora-associated trigger of plant defense in parsley and Arabidopsis. Plant J 2002, 32, 375–390. [Google Scholar]
- Yamakawa, H.; Mitsuhara, I.; Ito, N.; Seo, S.; Kamada, H.; Ohashi, Y. Transcriptionally and post-transcriptionally regulated response of 13 calmodulin genes to tobacco mosaic virus-induced cell death and wounding in tobacco plant. Eur. J. Biochem 2001, 268, 3916–3929. [Google Scholar]
- Takabatake, R.; Karita, E.; Seo, S.; Mitsuhara, I.; Kuchitsu, K.; Ohashi, Y. Pathogen-induced calmodulin isoforms in basal resistance against bacterial and fungal pathogens in tobacco. Plant Cell Physiol 2006, 48, 414–423. [Google Scholar]
- Sun, J.; Jiang, H.; Xu, Y.; Li, H.; Wu, X.; Xie, Q.; Li, C. The CCCH-type zinc finger proteins AtSZF1 and AtSZF2 regulate salt stress responses in Arabidopsis. Plant Cell Physiol 2007, 48, 1148–1158. [Google Scholar]
- Haas, I.G. BiP (GRP78) an essential hsp70 resident protein in the endoplasmic reticulum. Experientia 1994, 50, 1012–1020. [Google Scholar]
- Rutkowski, D.T.; Kaufman, R.J. A trip to the ER: Coping with stress. Trends Cell Biol 2004, 14, 20–28. [Google Scholar]
- Koizumi, N.; Ujino, T.; Sano, H.; Chrispeels, M.J. Overexpression of a gene that encodes the first enzyme in the biosynthesis of asparagine-linked glycans makes plants resistant to tunicamycin and obviates the tunicamycin-induced unfolded protein response. Plant Physiol 1999, 121, 353–361. [Google Scholar]
- Bonaventure, G.; Ohlrogge, J.B. Differential regulation of mRNA levels of acyl carrier protein isoforms in Arabidopsis. Plant Physiol 2002, 128, 223–235. [Google Scholar]
- Feng, D.; Witkowski, A.; Smith, S. Down-regulation of mitochondrial acyl carrier protein in mammalian cells compromises protein lipoylation and respiratory complex I and results in cell death. J. Biol. Chem 2009, 284, 11436–11445. [Google Scholar]
- Chakravarthy, S.; Tuori, R.P.; D’Ascenzo, M.D.; Fobert, P.R.; Despres, C.; Martin, G.B. The tomato transcription factor Pti4 regulates defense-related gene expression via GCC box and non-GCC box cis elements. Plant Cell 2003, 15, 3033–3050. [Google Scholar]
- Zago, E.; Morsa, S.; Dat, J.F.; Alard, P.; Ferrarini, A.; Inze, D.; Delledonne, M.; van Breusegem, F. Nitric oxide- and hydrogen peroxide-responsive gene regulation during cell death induction in tobacco. Plant Physiol 2006, 141, 404–411. [Google Scholar]
- Hwang, I.S.; Hwang, B.K. The pepper mannose-binding lectin gene CaMBL1 is required to regulate cell death and defense responses to microbial pathogens. Plant Physiol 2011, 155, 447–463. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lee, J.H.; Kim, Y.C.; Choi, D.; Park, J.M. Identification of Novel Pepper Genes Involved in Bax- or INF1-Mediated Cell Death Responses by High-Throughput Virus-Induced Gene Silencing. Int. J. Mol. Sci. 2013, 14, 22782-22795. https://doi.org/10.3390/ijms141122782
Lee JH, Kim YC, Choi D, Park JM. Identification of Novel Pepper Genes Involved in Bax- or INF1-Mediated Cell Death Responses by High-Throughput Virus-Induced Gene Silencing. International Journal of Molecular Sciences. 2013; 14(11):22782-22795. https://doi.org/10.3390/ijms141122782
Chicago/Turabian StyleLee, Jeong Hee, Young Cheol Kim, Doil Choi, and Jeong Mee Park. 2013. "Identification of Novel Pepper Genes Involved in Bax- or INF1-Mediated Cell Death Responses by High-Throughput Virus-Induced Gene Silencing" International Journal of Molecular Sciences 14, no. 11: 22782-22795. https://doi.org/10.3390/ijms141122782