Curcumin Inhibits Glutamate Release from Rat Prefrontal Nerve Endings by Affecting Vesicle Mobilization


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
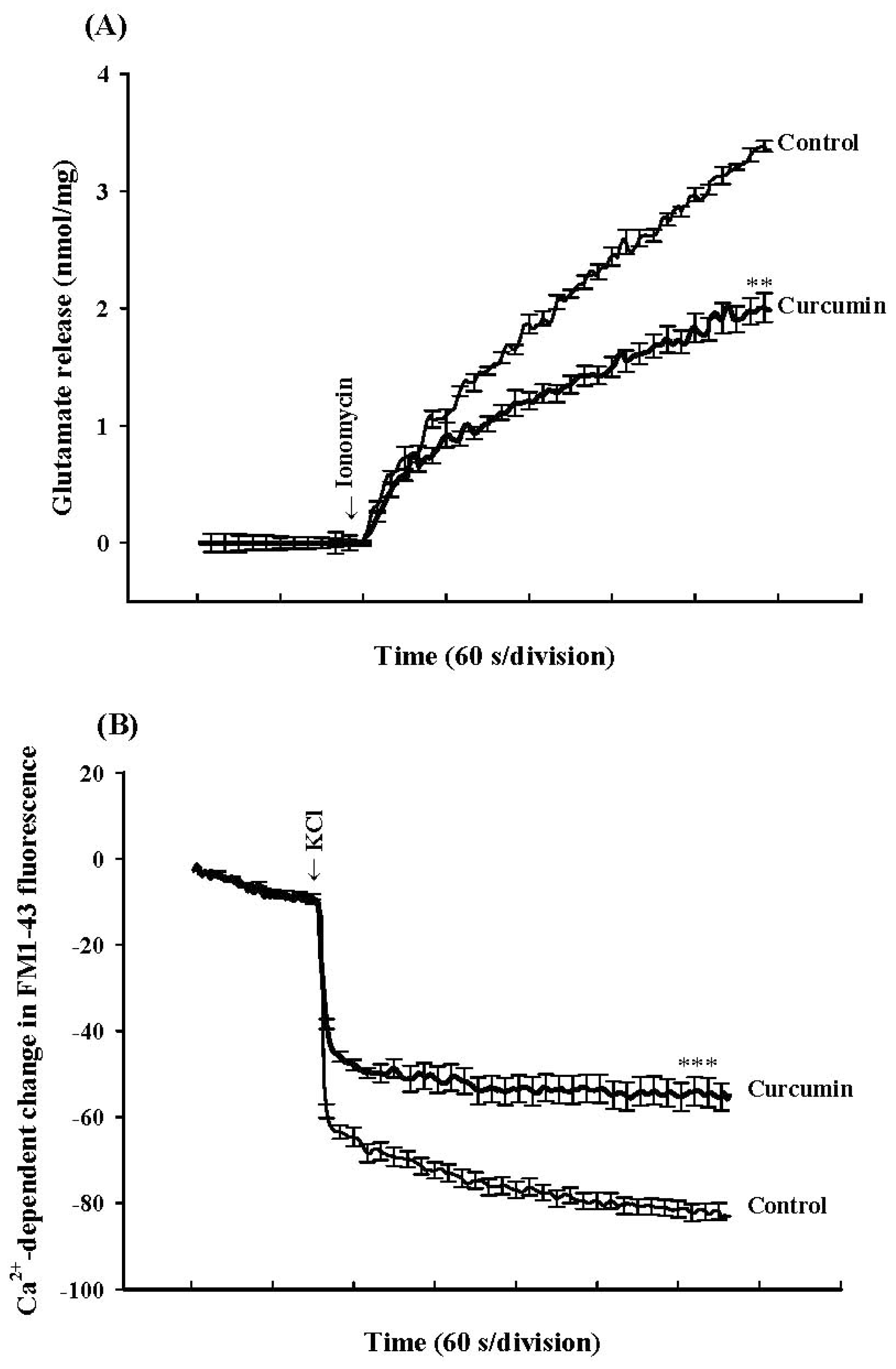
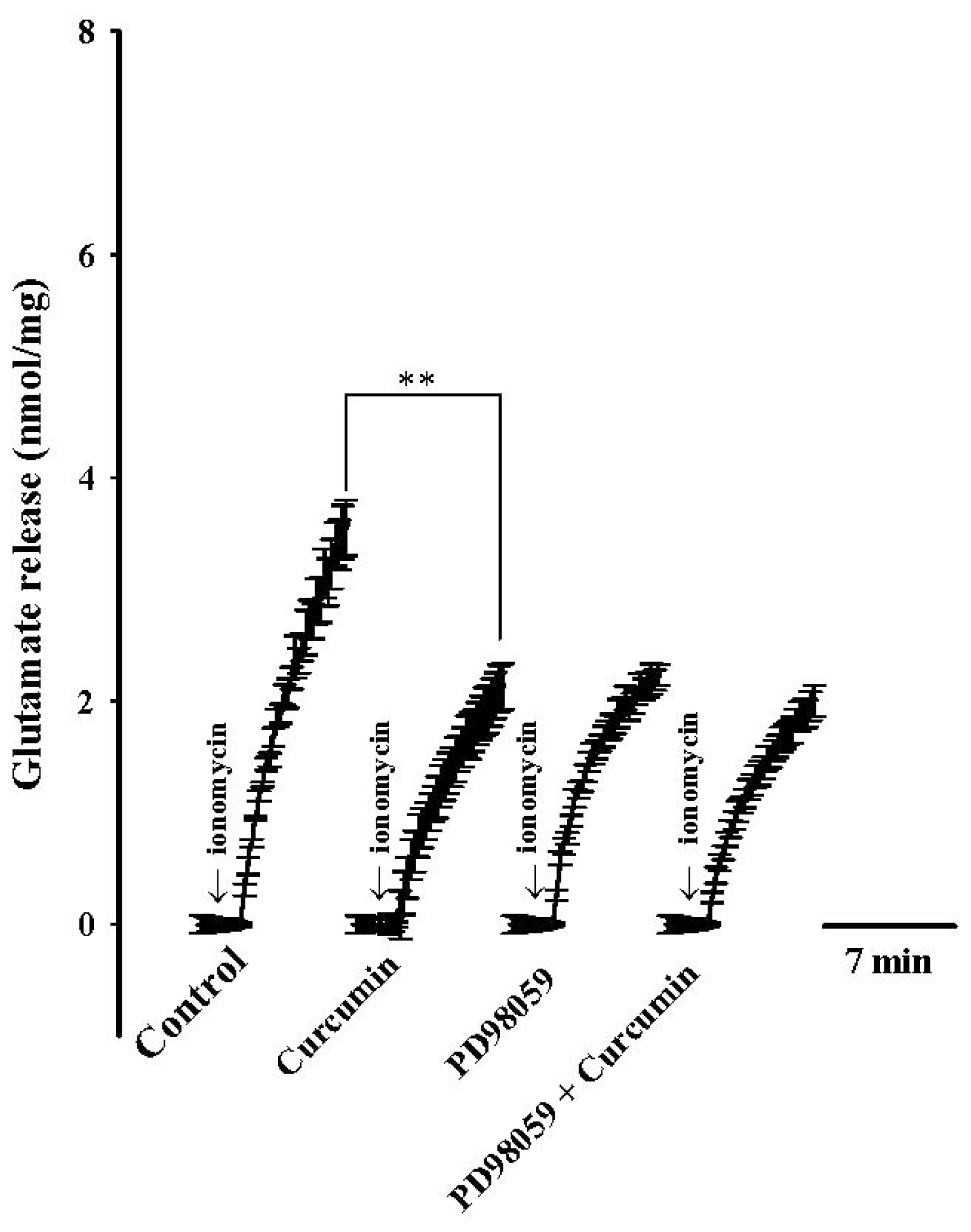
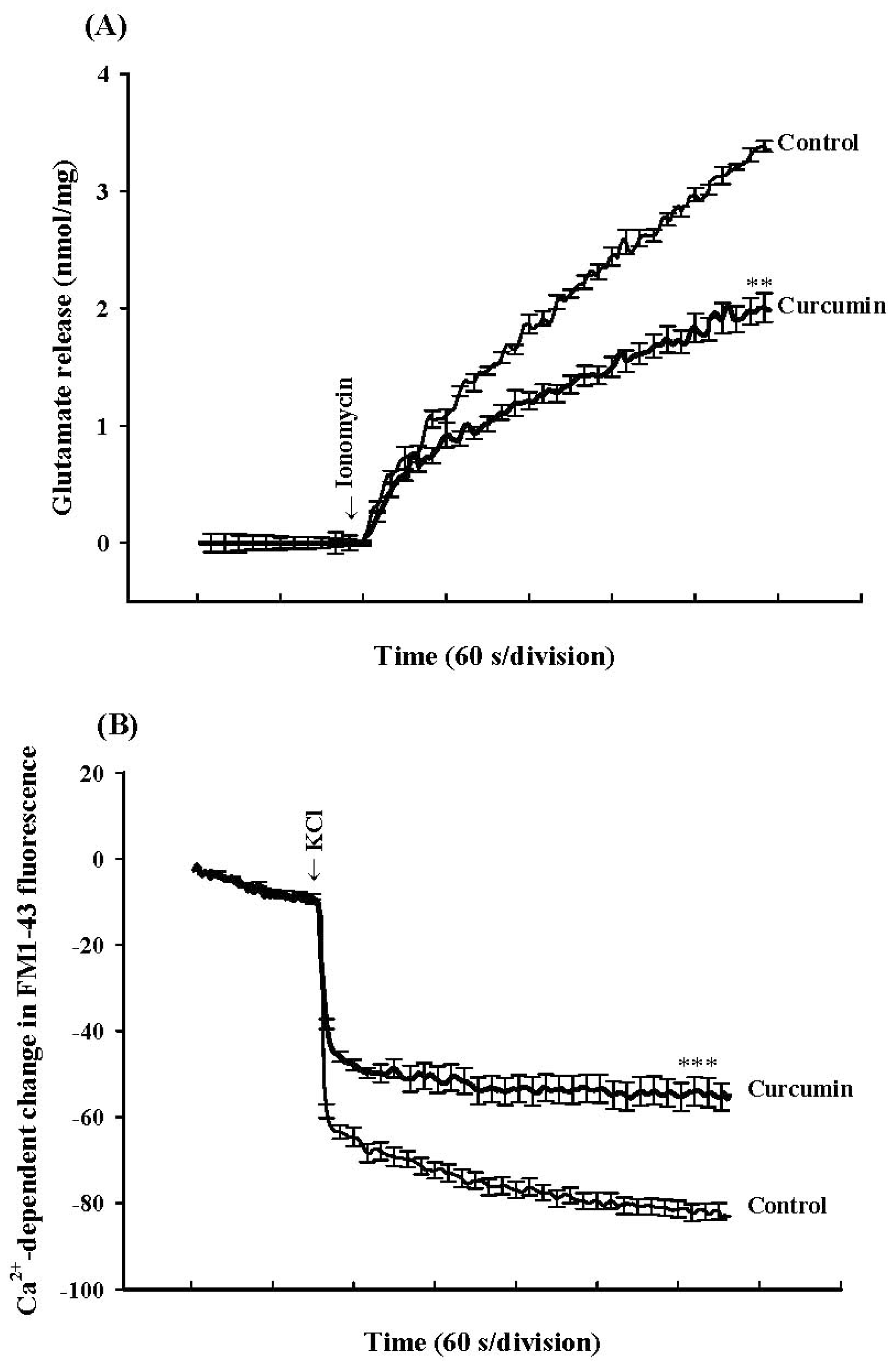
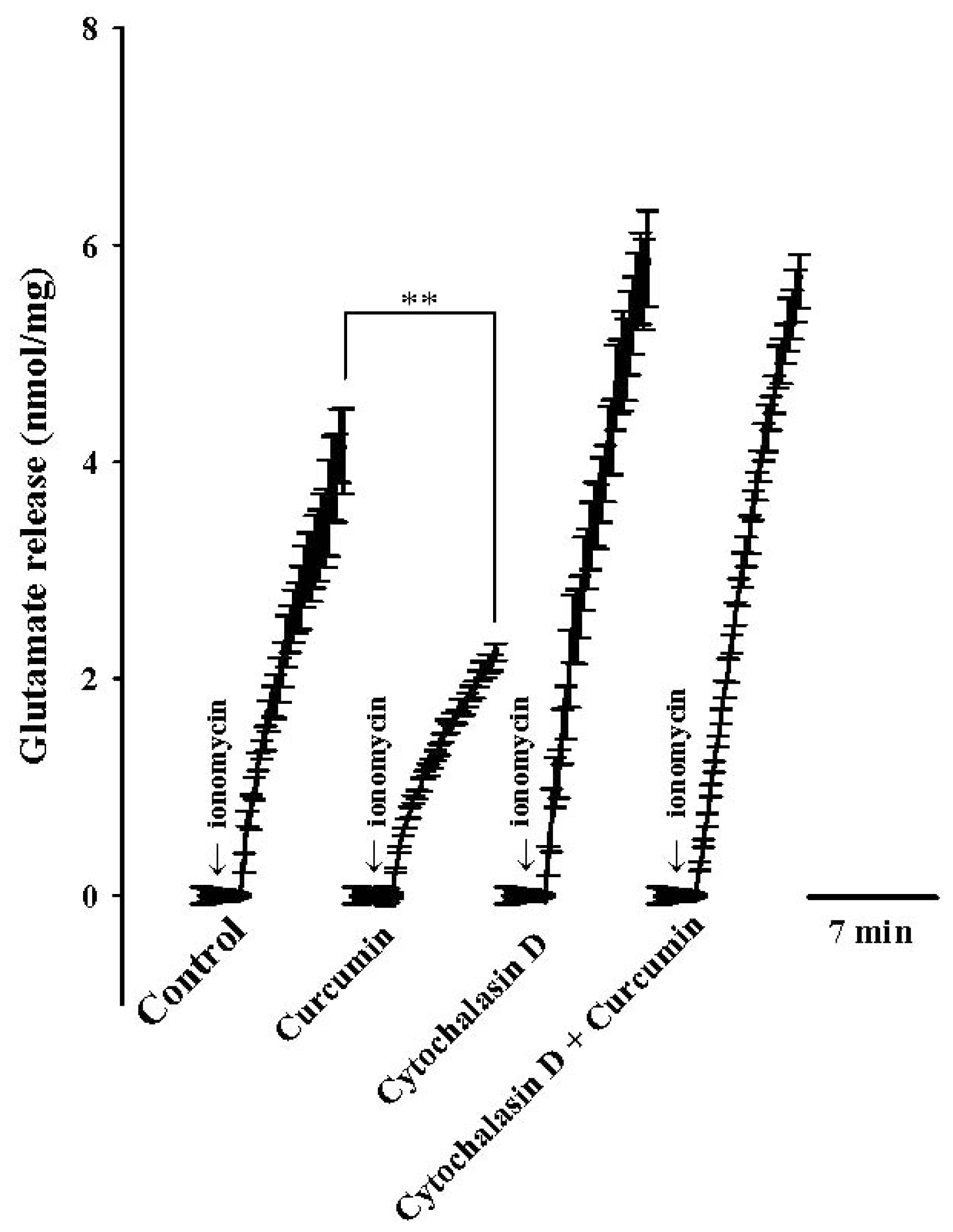
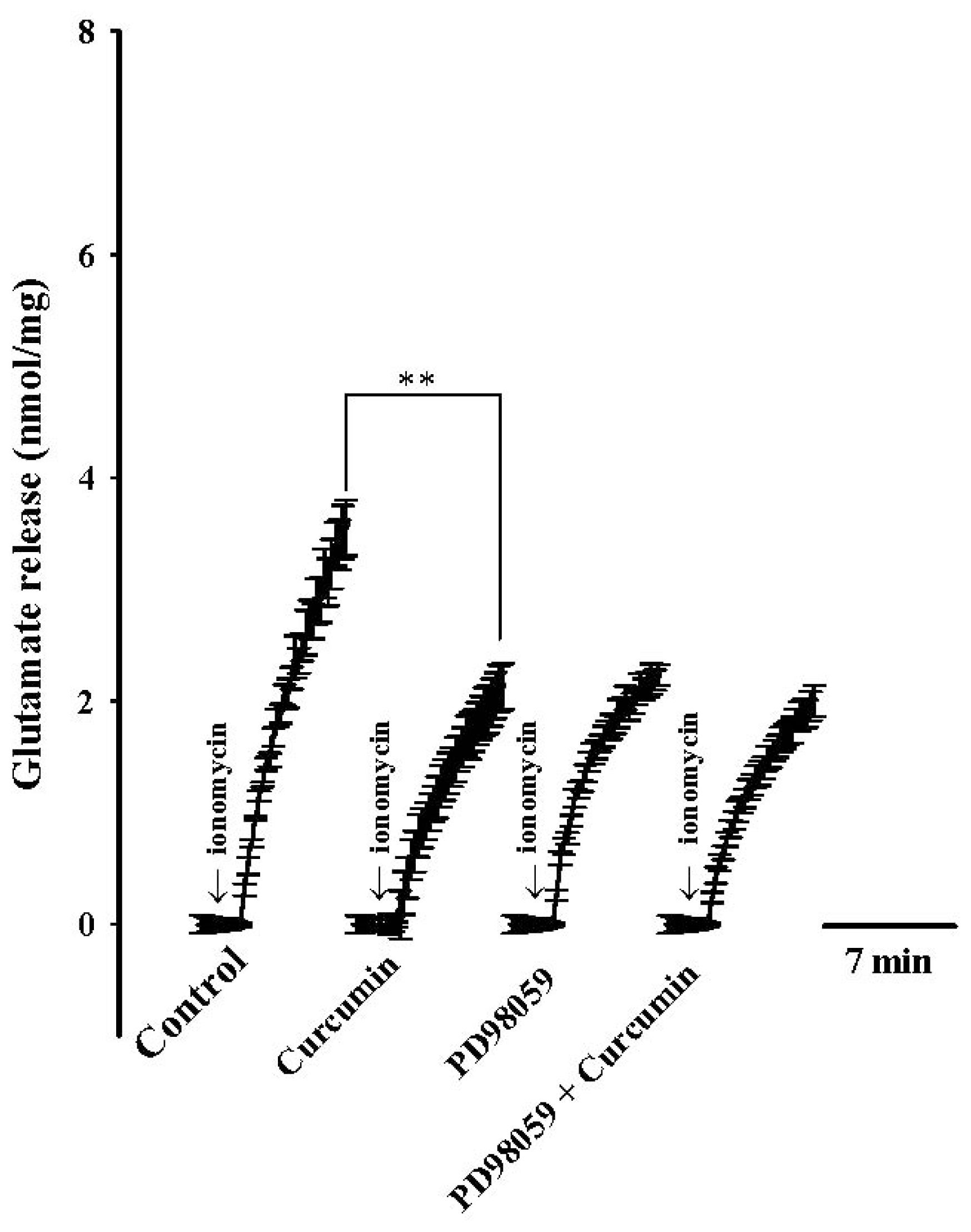
2. Results
3. Discussion
4. Experimental Section
4.1. Materials
4.2. Animals and Synaptosomal Preparation
4.3. Glutamate Release
4.4. Styryl Dye Release
4.5. Western Blot
4.6. Statistical Analysis
5. Conclusions
Acknowledgment
- Conflict of InterestThe authors declare that they have no conflict of interest.
References
- Di Santo, R.; Costi, R.; Artico, M.; Ragno, R.; Greco, G.; Novellino, E.; Marchand, C.; Pommier, Y. Design, synthesis and biological evaluation of heteroaryl diketohexenoic and diketobutanoic acids as HIV-1 integrase inhibitors endowed with antiretroviral activity. Il Farmaco 2005, 60, 409–417. [Google Scholar]
- Schaffer, M.; Schaffer, P.M.; Zidan, J.; Bar Sela, G. Curcuma as a functional food in the control of cancer and inflammation. Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 588–597. [Google Scholar]
- Motterlini, R.; Foresti, R.; Bassi, R.; Green, C.J. Curcumin, an antioxidant and anti-inflammatory agent, induces heme oxygenase-1 and protects endothelial cells against oxidative stress. Free Radic. Biol. Med 2000, 28, 1303–1312. [Google Scholar]
- Thiyagarajan, M.; Sharma, S.S. Neuroprotective effect of curcumin in middle cerebral artery occlusion induced focal cerebral ischemia in rats. Life Sci 2004, 74, 969–985. [Google Scholar]
- Mizushina, Y.; Hirota, M.; Murakami, C.; Ishidoh, T.; Kamisuki, S.; Shimazaki, N.; Takemura, M.; Perpelescu, M.; Suzuki, M.; Yoshida, H.; et al. Some anti-chronic inflammatory compounds are DNA polymerase l-specific inhibitors. Biochem. Pharmacol 2003, 66, 1935–1944. [Google Scholar]
- Park, S.Y.; Kim, H.S.; Cho, E.K.; Kwon, B.Y.; Phark, S.; Hwang, K.W.; Sul, D. Curcumin protected PC12 cells against beta-amyloid-induced toxicity through the inhibition of oxidative damage and tau hyperphosphorylation. Food Chem. Toxicol 2008, 46, 2881–2887. [Google Scholar]
- Vajragupta, O.; Boonchoong, P.; Watanabe, H.; Tohda, M.; Kummasud, N.; Sumanont, Y. Manganese complexes of curcumin and its derivatives: Evaluation for the radical scavenging ability and neuroprotective activity. Free Radic. Biol. Med 2003, 35, 1632–1644. [Google Scholar]
- Suh, H.W.; Kang, S.; Kwon, K.S. Curcumin attenuates glutamate-induced HT22 cell death by suppressing MAP kinase signaling. Mol. Cell Biochem 2007, 298, 187–194. [Google Scholar]
- Pan, R.; Qiu, S.; Lu, D.X.; Dong, J. Curcumin improves learning and memory ability and its neuroprotective mechanism in mice. Chin. Med. J 2008, 121, 832–839. [Google Scholar]
- Ghoneim, A.I.; Abdel-Naim, A.B.; Khalifa, A.E.; El-Denshary, E.S. Protective effects of curcumin against ischaemia/reperfusion insult in rat forebrain. Pharmacol. Res 2002, 46, 273–279. [Google Scholar]
- Du, P.; Li, X.; Lin, H.J.; Peng, W.F.; Liu, J.Y.; Ma, Y.; Fan, W.; Wang, X. Curcumin inhibits amygdaloid kindled seizures in rats. Chin. Med. J 2009, 122, 1435–1438. [Google Scholar]
- Rajeswari, A. Curcumin protects mouse brain from oxidative stress caused by 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine. Eur. Rev. Med. Pharmacol. Sci 2006, 10, 157–161. [Google Scholar]
- Zbarsky, V.; Datla, K.P.; Parkar, S.; Rai, D.K.; Aruoma, O.I.; Dexter, D.T. Neuroprotective properties of the natural phenolic antioxidants curcumin and naringenin but not quercetin and fisetin in a 6-OHDA model of Parkinson’s disease. Free Radic. Res 2005, 39, 1119–1125. [Google Scholar]
- Fonnum, F. Glutamate: A neurotransmitter in mammalian brain. J. Neurochem 1984, 42, 1–11. [Google Scholar]
- Greenamyre, J.T.; Porter, R.H. Anatomy and physiology of glutamate in the CNS. Neurology 1994, 44, S7–S13. [Google Scholar]
- Meldrum, B.S. Glutamate as a neurotransmitter in the brain: Review of physiology and pathology. J. Nutr 2000, 130, 1007S–1015S. [Google Scholar]
- Obrenovitch, T.P.; Urenjak, J. Altered glutamatergic transmission in neurological disorders: From high extracellular glutamate to excessive synaptic efficacy. Prog. Neurobiol 1997, 51, 39–87. [Google Scholar]
- Raiteri, L.; Raiteri, M. Synaptosomes still viable after 25 years of superfusion. Neurochem. Res 2000, 25, 1265–1274. [Google Scholar]
- Lin, T.Y.; Lu, C.W.; Wang, C.C.; Wang, Y.C.; Wang, S.J. Curcumin inhibits glutamate release in nerve terminals from rat prefrontal cortex: Possible relevance to its antidepressant mechanism. Prog. Neuropsychopharmacol. Biol. Psychiatry 2011, 35, 1785–1793. [Google Scholar]
- Sihra, T.S.; Bogonez, E.; Nicholls, D.G. Localized Ca2+ entry preferentially effects protein dephosphorylation, phosphorylation, and glutamate release. J. Biol. Chem 1992, 267, 1983–1989. [Google Scholar]
- Baldwin, M.L.; Rostas, J.A.; Sim, A.T. Two models of exocytosis from synaptosomes are differentially regulated by protein phosphatase types 2A and 2B. J. Neurochem 2003, 85, 1190–1199. [Google Scholar]
- Greengard, P.; Valtorta, F.; Czernik, A.J.; Benfenati, F. Synaptic vesicle phosphoproteins and regulation of synaptic function. Science 1993, 259, 780–785. [Google Scholar]
- Pereira, D.B.; Carvalho, A.P.; Duarte, C.B. Non-specific effects of the MEK inhibitors PD098,059 and U0126 on glutamate release from hippocampal synaptosomes. Neuropharmacology 2002, 42, 9–19. [Google Scholar]
- Lin, T.Y.; Lu, C.W.; Wang, S.J. Astaxanthin inhibits glutamate release in rat cerebral cortex nerve terminals via suppression of voltage-dependent Ca(2+) entry and mitogen-activated protein kinase signaling pathway. J. Agric. Food Chem 2010, 58, 8271–8278. [Google Scholar]
- Alessi, D.R.; Cuenda, A.; Cohen, P.; Dudley, D.T.; Saltiel, A.R. PD 098059 is a specific inhibitor of the activation of mitogen-activated protein kinase kinase in vitro and in vivo. J Biol. Chem 1995, 270, 27489–27494. [Google Scholar]
- Terrian, D.M.; Ways, D.K. Persistent enhancement of sustained calcium-dependent glutamate release by phorbol esters: Role of calmodulin-independent serine/threonine phosphorylation and actin disassembly. J. Neurochem 1995, 64, 181–190. [Google Scholar]
- Cooper, J.A. Effects of cytochalasin and phalloidin on actin. J. Cell Biol 1987, 105, 1473–1478. [Google Scholar]
- Zhang, L.; Ruehr, M.L.; Dorman, R.V. Arachidonic acid and oleoylacetylglycerol induce a synergistic facilitation of Ca21-dependent glutamate release from hippocampal mossy fiber nerve endings. J. Neurochem 1996, 66, 177–185. [Google Scholar]
- Grewal, S.; York, R.; Stork, P. Extracellular-signal-regulated kinase signaling in neurons. Curr. Opin. Neurobiol 1999, 9, 544–553. [Google Scholar]
- Chi, P.; Greengard, P.; Pyan, T.A. Synaptic vesicle mobilization is regulated by distinct synapsin I phosphorylation pathways at different frequencies. Neuron 2003, 38, 69–78. [Google Scholar]
- Jovabovic, J.N.; Benfenati, F.; Siow, Y.L.; Sihra, T.S.; Sanghera, J.S.; Pelech, S.L.; Greengard, P.; Czernicj, A.J. Neurotrophins stimulate phosphorylation of synapsin I by MAP kinase and regulate synapsin I-actin interactions. Proc. Natl. Acad. Sci. USA 1996, 93, 3679–3683. [Google Scholar]
- Chi, P.; Greengard, P.; Pyan, T.A. Synapsin dispersion and reclustering during synaptic activity. Nat. Neurosci 2001, 4, 1187–1193. [Google Scholar]
- Hilfiker, F.E. Vesicle pools and synapsins: New insights into old enigmas. Brain Cell Biol 2006, 35, 107–115. [Google Scholar]
- Jovanovic, J.N.; Czernik, A.J.; Fienberg, A.A.; Greengard, P.; Sihra, T.S. Synapsins as mediators of BDNF-enhanced neurotransmitter release. Nat. Neurosci 2000, 3, 323–329. [Google Scholar]
- Wang, R.; Li, Y.B.; Li, Y.H.; Xu, Y.; Wu, H.L.; Li, X.J. Curcumin protects against glutamate excitotoxicity in rat cerebral cortical neurons by increasing brain-derived neurotrophic factor level and activating TrkB. Brain Res 2008, 1210, 84–91. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem 1976, 72, 248–254. [Google Scholar]
- Dunkley, P.R.; Jarvie, P.E.; Heath, J.W.; Kidd, G.J.; Rostas, J.A. A rapid method for isolation of synaptosomes on Percoll gradients. Brain Res 1986, 372, 115–129. [Google Scholar]
- Dunkley, P.R.; Jarvie, P.E.; Robinson, P.J. A rapid Percoll gradient procedure for preparation of synaptosomes. Nat. Protoc 2008, 3, 1718–1728. [Google Scholar]
- Nicholls, D.G. The glutamatergic nerve terminal. Eur. J. Biochem 1993, 212, 613–631. [Google Scholar]

© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lin, T.Y.; Lu, C.W.; Huang, S.K.; Wang, S.J. Curcumin Inhibits Glutamate Release from Rat Prefrontal Nerve Endings by Affecting Vesicle Mobilization. Int. J. Mol. Sci. 2012, 13, 9097-9109. https://doi.org/10.3390/ijms13079097
Lin TY, Lu CW, Huang SK, Wang SJ. Curcumin Inhibits Glutamate Release from Rat Prefrontal Nerve Endings by Affecting Vesicle Mobilization. International Journal of Molecular Sciences. 2012; 13(7):9097-9109. https://doi.org/10.3390/ijms13079097
Chicago/Turabian StyleLin, Tzu Yu, Cheng Wei Lu, Shu Kuei Huang, and Su Jane Wang. 2012. "Curcumin Inhibits Glutamate Release from Rat Prefrontal Nerve Endings by Affecting Vesicle Mobilization" International Journal of Molecular Sciences 13, no. 7: 9097-9109. https://doi.org/10.3390/ijms13079097