Molecular Mechanisms of (R,R)ZX-5 on NO Synthesis and Its Anti-Angiogenic Effect

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
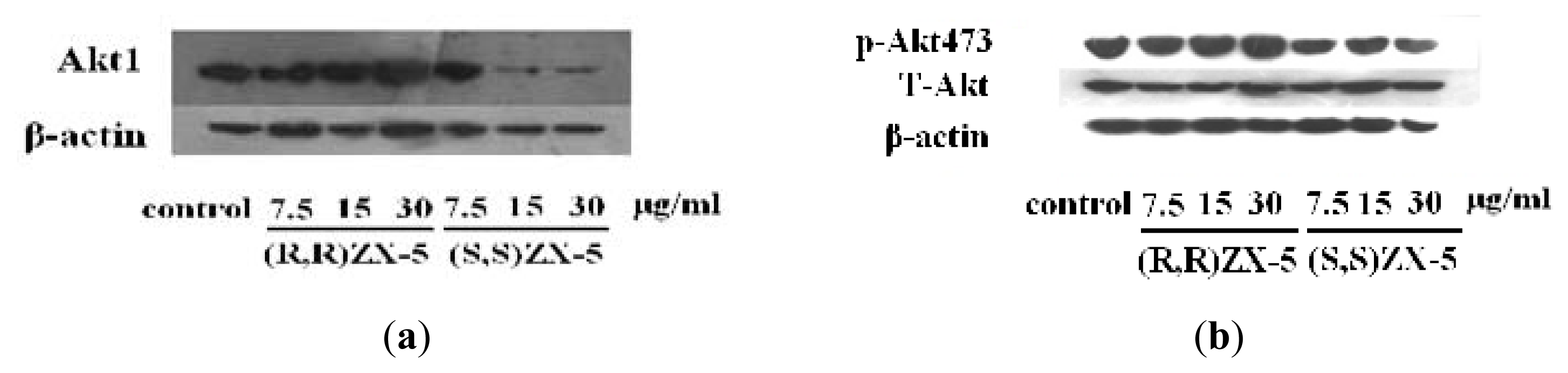
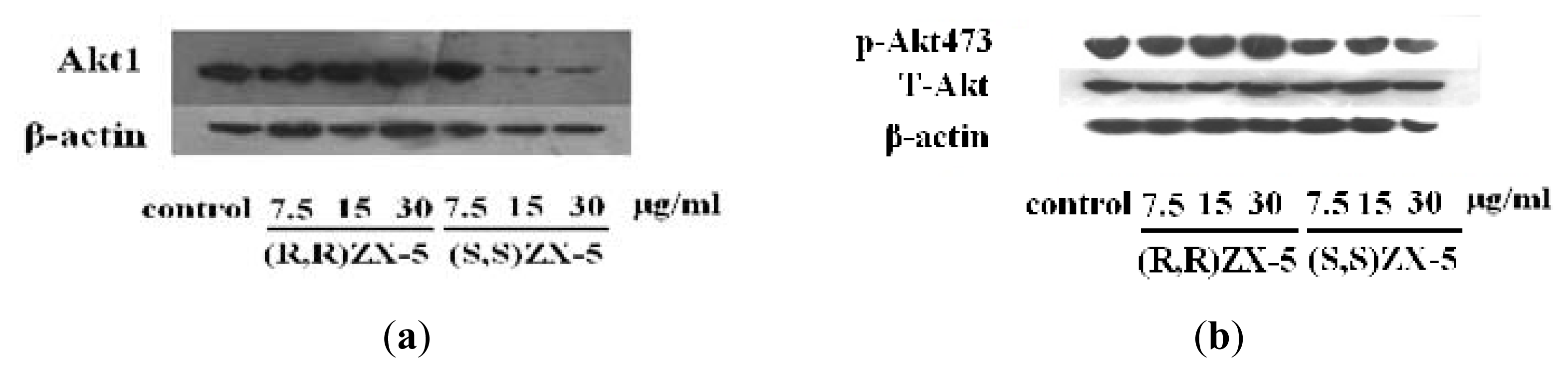
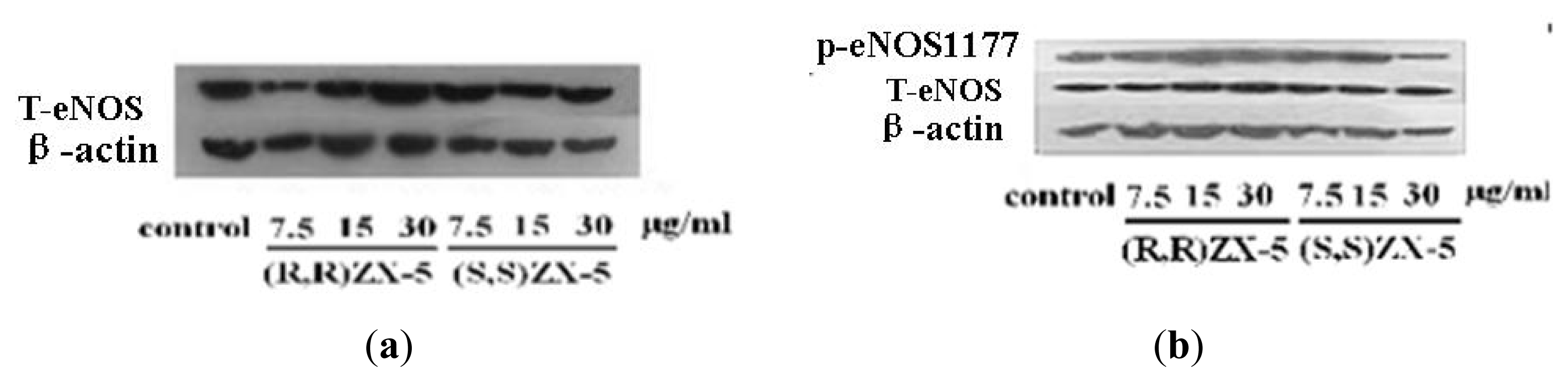
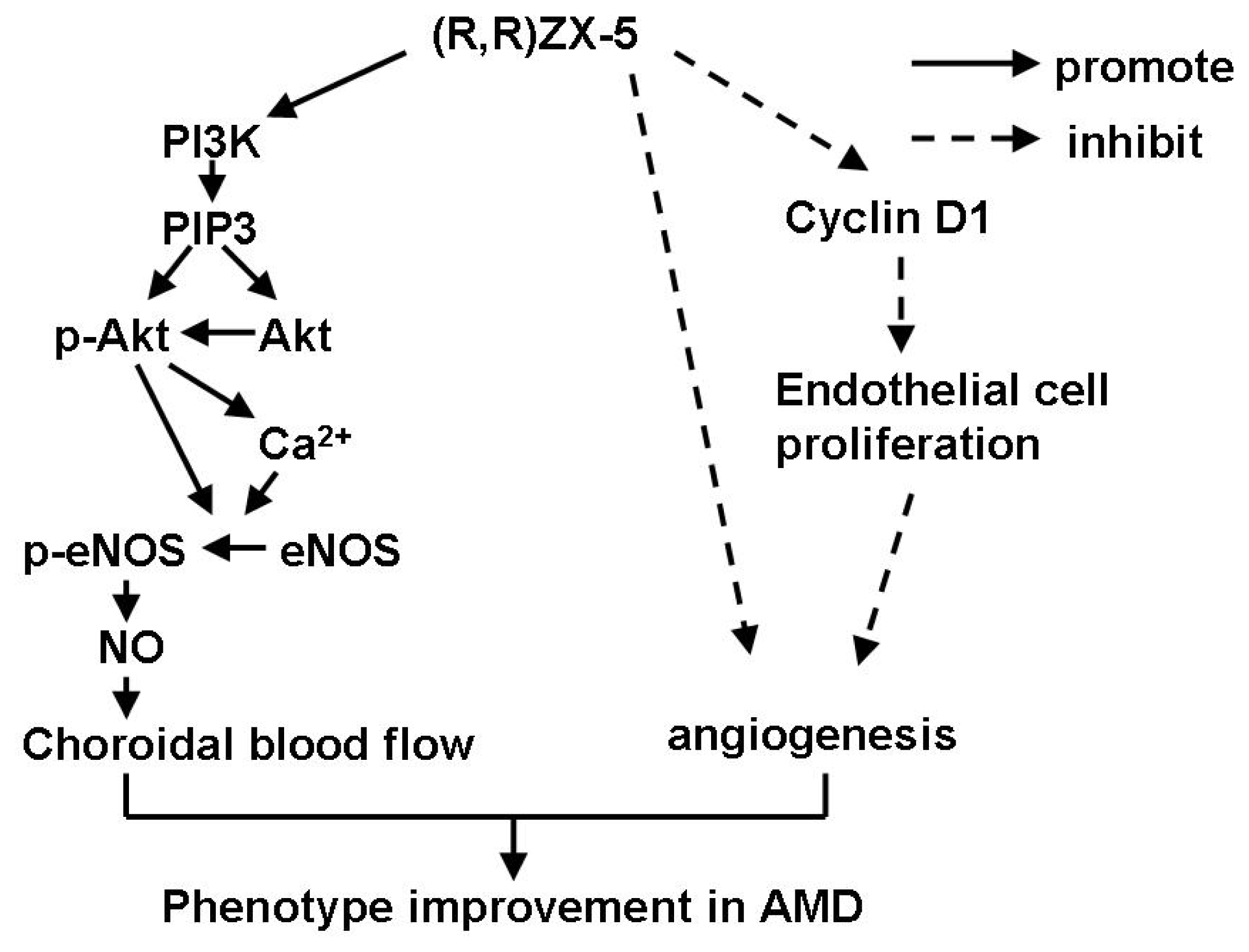
2.1. Effect of ZX-5 on Akt, Phospho-Akt (Thr473), eNOS and Phospho-eNOS (Ser1177) Levels in HRECs
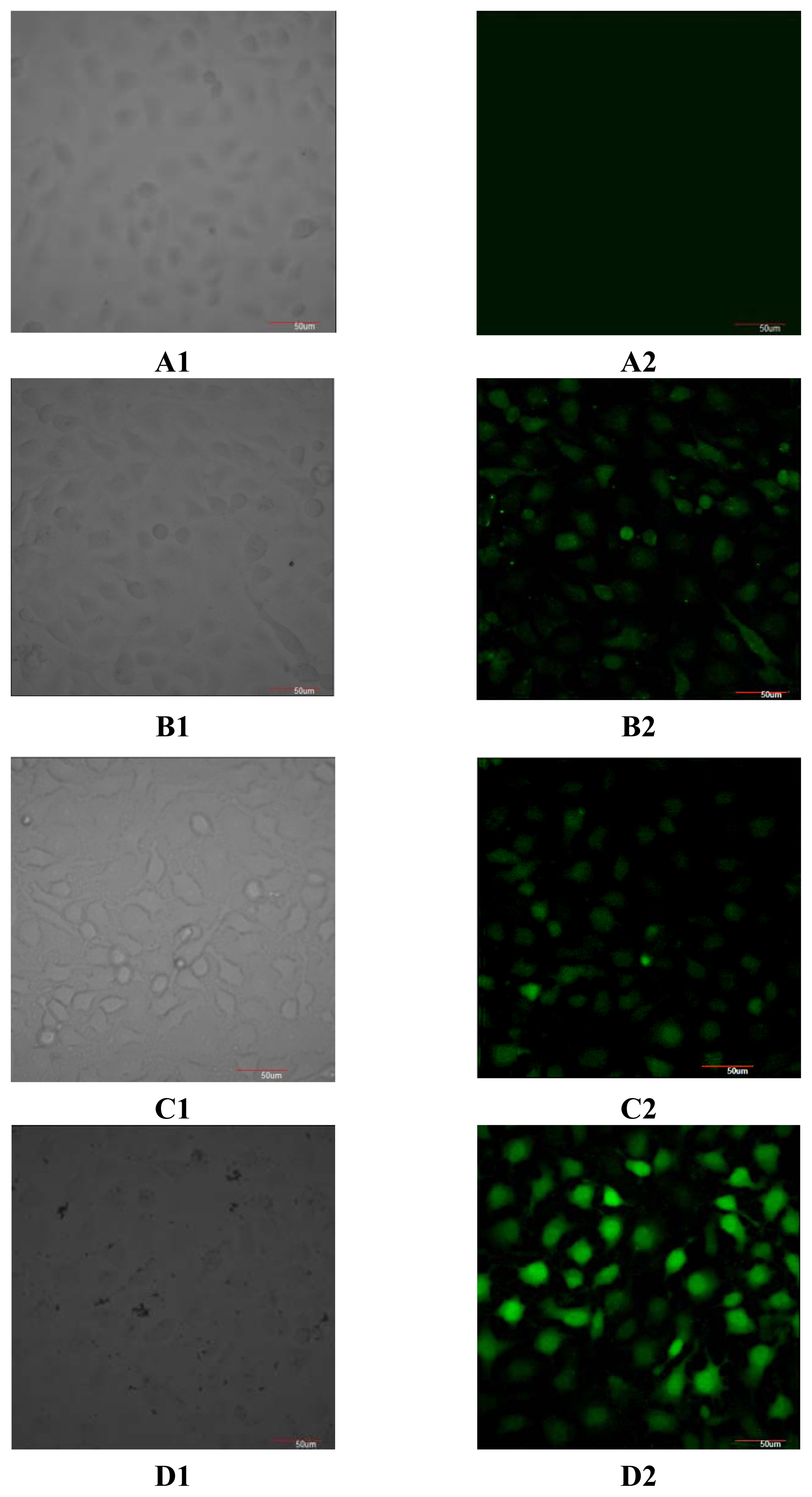
2.2. Effect of ZX-5 on Intracellular Free Ca2+ Concentration in HRECs
2.3. Effect of ZX-5 on Cyclin D1 Level in HRECs
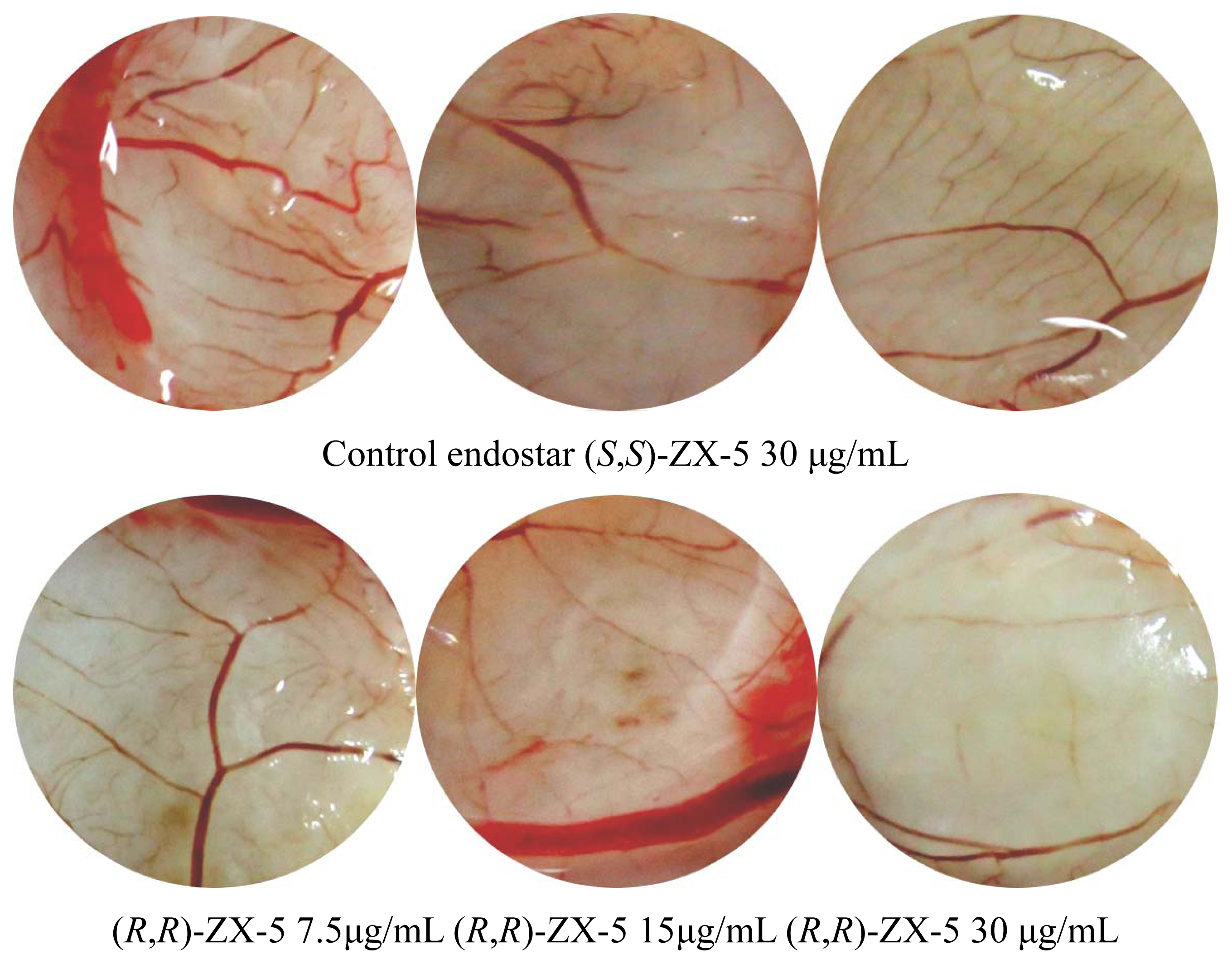
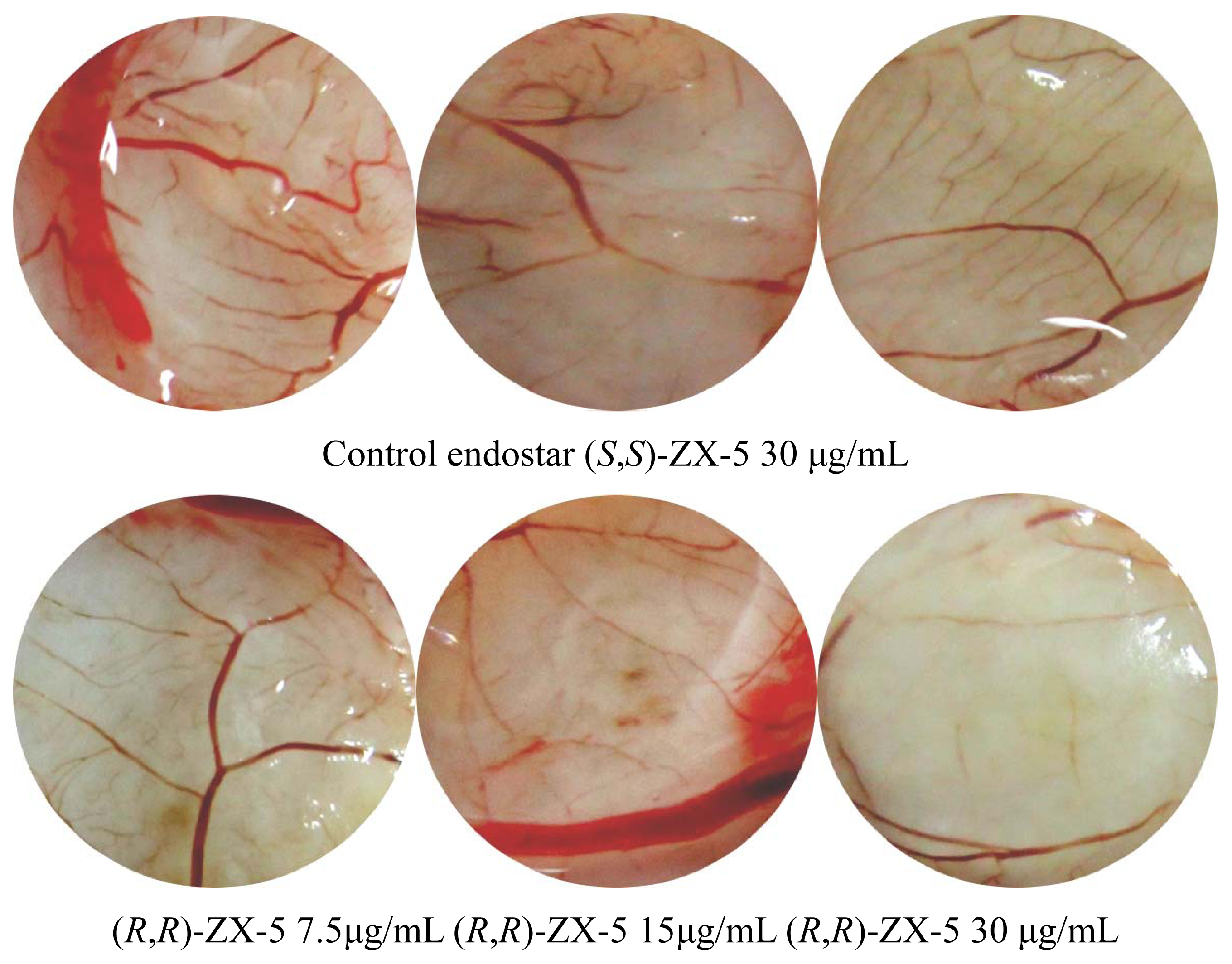
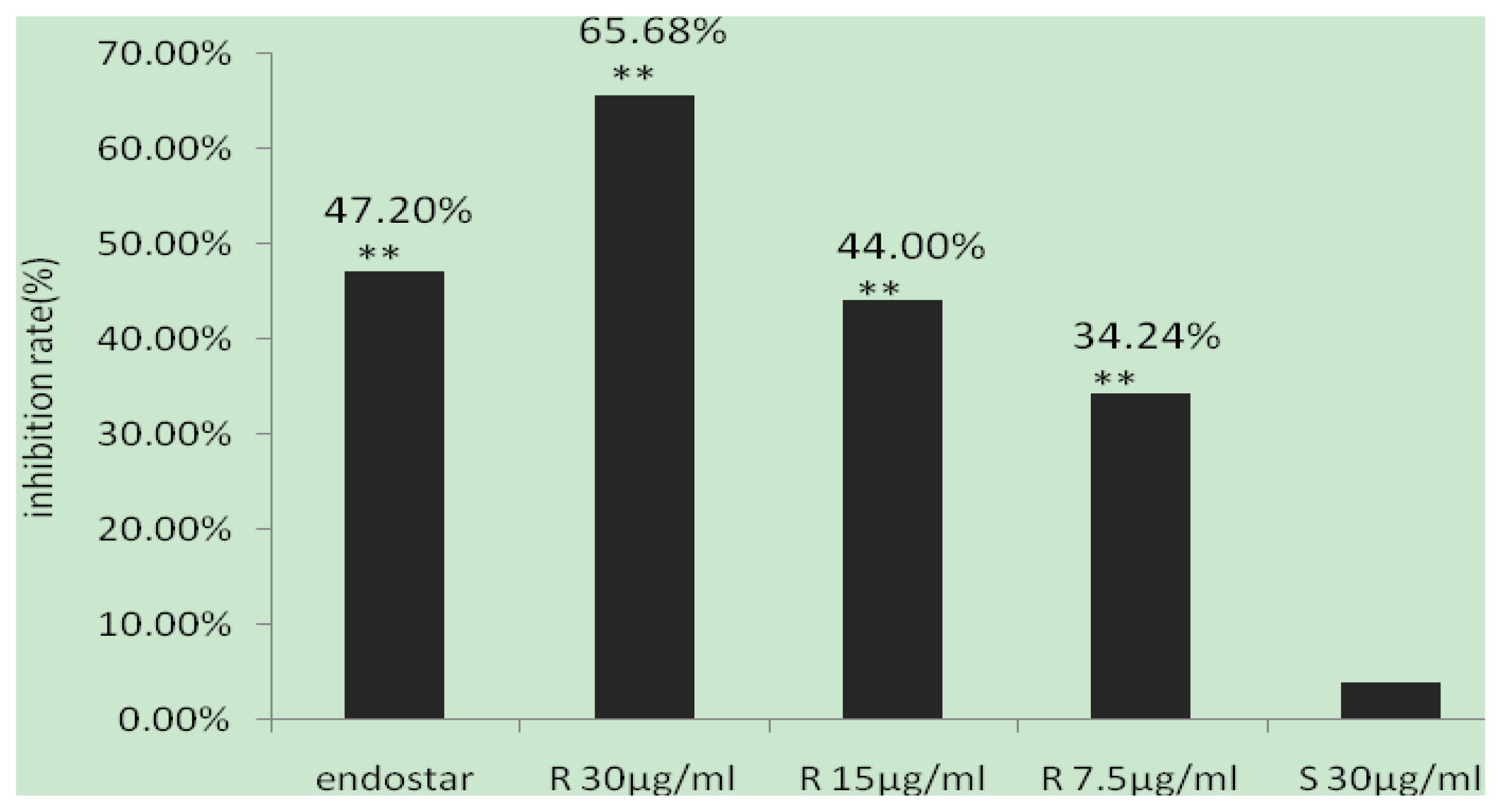
2.4. Inhibitory Effects of (R,R)ZX-5 on Angiogenesis in the CAM Model
3. Experimental Section
3.1. Cell Culture
3.2. Cell Treatment
3.3. Western Blot Analysis
3.4. Measurement of Intracellular Free Ca2+ Concentration by LSCM (Laser Scanning Confocal Microscope)
3.5. CAM Angiogenesis Assays
3.6. Statistical Analysis
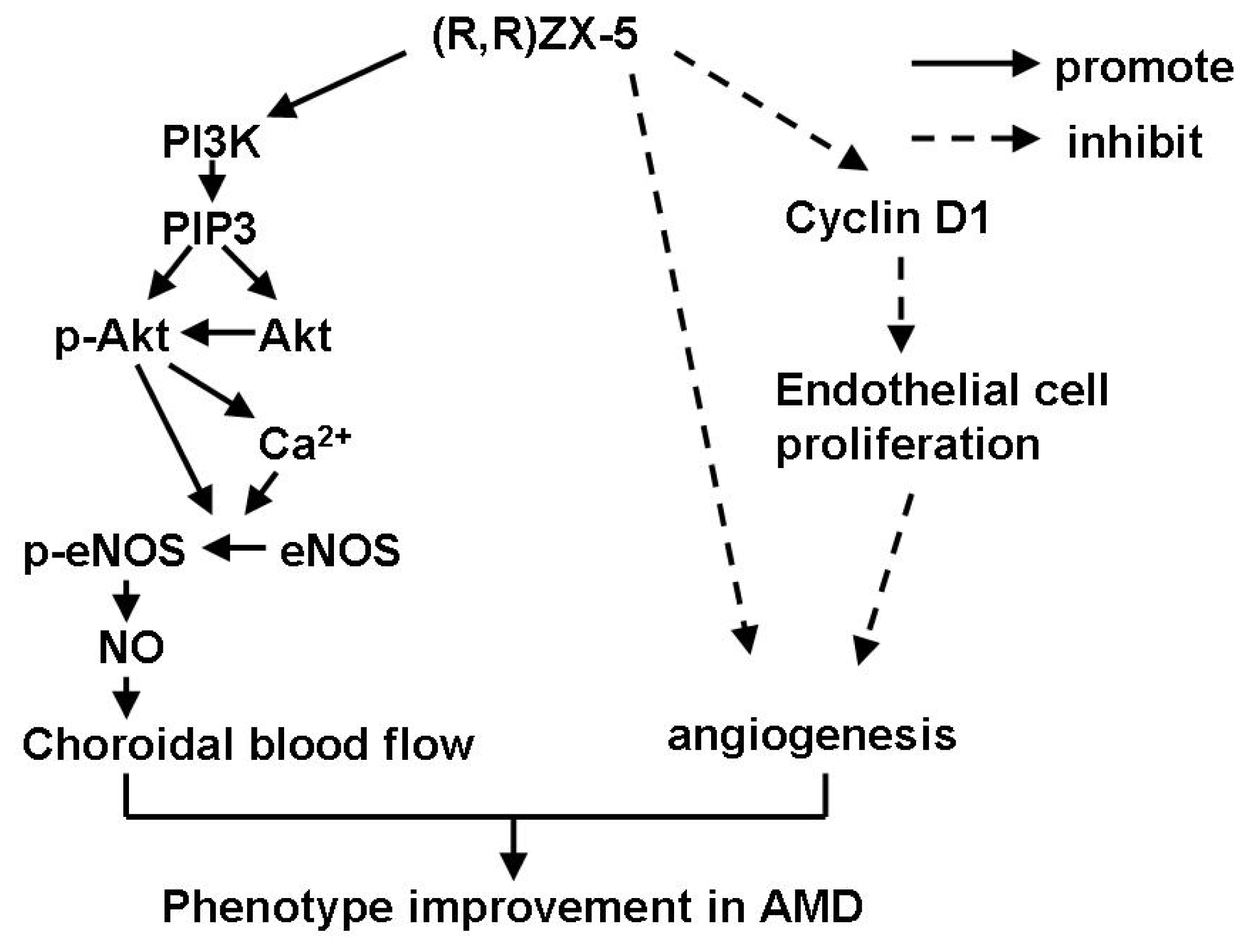
4. Conclusions
Acknowledgements
- Declaration of InterestThe authors announce that there are no actual or potential conflicts of interest of this article.
References
- Hua, J.; Spee, C.; Kase, S.; Rennel, E.S.; Magnussen, A.L.; Qiu, Y.; Varey, A.; Dhayade, S.; Churchill, A.J.; Harper, S.J.; et al. Recombinant human VEGF165b inhibits experimental Choroidal Neovascularization. Invest. Ophthalmol. Vis. Sci 2010, 51, 4282–4288. [Google Scholar]
- Ebrahem, Q.; Renganathan, K.; Sears, J.; Vasanji, A.; Gu, X.; Lu, L.; Salomon, R.G.; Crabb, J.W.; Anand-Apte, B. Carboxyethylpyrrole oxidative protein modifications stimulate neovascularization: Implications for age-related macular degeneration. Proc. Natl. Acad. Sci. USA 2006, 103, 13480–13484. [Google Scholar]
- Chiou, G.C. Review: Effects of nitric oxide on eye diseases and their treatment. J. Ocul. Pharmacol. Ther 2001, 17, 189–198. [Google Scholar]
- Xuan, B.; Xu, X.R.; Chiou, G.C.Y.; Zhang, Y.H.; Peng, S.X. Relationship between nitric oxide production and choroidal blood flow. J. Ocul. Pharmacol. Ther 2003, 19, 247–253. [Google Scholar]
- Bhutto, I.A.; Baba, T.; Merges, C.; McLeod, D.S.; Lutty, G.A. Low nitric oxides synthases (NOSs) in eyes with age-related macular degeneration (AMD). Exp. Eye Res 2010, 90, 155–167. [Google Scholar]
- Xu, H.M.; Wei, J.; Pan, L.; Lin, H.Y.; Wang, W.Q.; Zhang, Y.H.; Shen, Z.L. The mechanism of (R,R)ZX-5 on increasing NO release. Int. J. Mol. Sci 2010, 11, 3323–3333. [Google Scholar]
- Ji, X.B.; Zhang, Y.H.; Lai, Y.S.; Lin, H.Y.; Shen, Z.L.; Peng, S.X. Synthesis and Biological Evaluation of Enantiomers of ZX-5 as Novel Anti-AMD Agents. Proceedings of the 50th Anniversary Meeting, Eighth Tetrahedron Symposium: Delegate Manual, Berlin, Germany, 26–29 June 2007; Elsvier: Oxford UK, 2007; pp. 92–93. [Google Scholar]
- Husain, D.; Meyer, R.D.; Mehta, M.; Pfeifer, W.M.; Chou, E.; Nsvruzbekov, G.; Ahmed, E.; Rahimi, N. Role of c-Cbl-Dependent Regulation of Phospholipase Cγ1 Activation in Experimental Choroidal Neovascularization. Invest. Ophthalmol. Vis. Sci 2010, 51, 6803–6809. [Google Scholar]
- Ribatti, D.; Nico, B.; Vacca, A.; Roncali, L.; Burri, P.H.; Djonov, V. Chorioallantoic membrane capillary bed: A useful target for studying angiogenesis and anti-angiogenesis in vivo. Anat. Rec 2001, 264, 317–324. [Google Scholar]
- Stewart, E.A.; Samaranayake, G.J.; Browning, A.C.; Hopkinson, A.; Amoaku, W.M. Comparison of choroidal and retinal endothelial cells: Characteristics and response to VEGF isoforms and anti-VEGF treatments. Exp. Eye Res 2011, 93, 761–766. [Google Scholar]
- Fornaro, M.; Plescia, J.; Chheang, S.; Tallini, G.; Zhu, Y.M.; King, M.; Altieri, D.C.; Languino, L.R. Fibronectin protects prostate cancer cells from tumor necrosis factor-alpha-induced apoptosis via the AKT/survivin pathway. J. Biol. Chem 2003, 278, 50402–50411. [Google Scholar]
- Bauer, P.M.; Fulton, D.; Boo, Y.C.; Sorescu, G.P.; Kemp, B.E.; Sessa, W.C. Compensatory phosphorylation and protein-protein interactions revealed by loss of function and gain of function mutants of multiple serine phosphorylation sites in endothelial nitric oxide synthase. J. Biol. Chem 2003, 278, 14841–14849. [Google Scholar]
- Xue, L.; Chiang, L.; Kang, C.; Winoto, A. The role of the PI3K-AKT pathway in T cell development beyond the β checkpoint. Eur. J. Immunol 2008, 38, 3200–3207. [Google Scholar]
- Hofer, A.M. Another dimension to calcium signaling: A look at extracellular calcium. J. Cell Sci 2005, 118, 855–862. [Google Scholar]
- Fleming, I.; Busse, R. Molecular mechanisms involved in the regulation of the endothelial nitric oxide synthase. Am. J. Physiol. Regul. Integr. Comp. Physiol 2003, 284, R1–R12. [Google Scholar]


© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Pan, L.; Hu, J.-L.; Wang, W.-J.; Zhang, X.-J.; Wei, J.; Liu, Z.-D.; Zhang, Y.-H.; Xu, H.-M. Molecular Mechanisms of (R,R)ZX-5 on NO Synthesis and Its Anti-Angiogenic Effect. Int. J. Mol. Sci. 2012, 13, 2717-2726. https://doi.org/10.3390/ijms13032717
Pan L, Hu J-L, Wang W-J, Zhang X-J, Wei J, Liu Z-D, Zhang Y-H, Xu H-M. Molecular Mechanisms of (R,R)ZX-5 on NO Synthesis and Its Anti-Angiogenic Effect. International Journal of Molecular Sciences. 2012; 13(3):2717-2726. https://doi.org/10.3390/ijms13032717
Chicago/Turabian StylePan, Li, Jia-Liang Hu, Wen-Jing Wang, Xiao-Juan Zhang, Jin Wei, Zhen-Dong Liu, Yi-Hua Zhang, and Han-Mei Xu. 2012. "Molecular Mechanisms of (R,R)ZX-5 on NO Synthesis and Its Anti-Angiogenic Effect" International Journal of Molecular Sciences 13, no. 3: 2717-2726. https://doi.org/10.3390/ijms13032717
APA StylePan, L., Hu, J.-L., Wang, W.-J., Zhang, X.-J., Wei, J., Liu, Z.-D., Zhang, Y.-H., & Xu, H.-M. (2012). Molecular Mechanisms of (R,R)ZX-5 on NO Synthesis and Its Anti-Angiogenic Effect. International Journal of Molecular Sciences, 13(3), 2717-2726. https://doi.org/10.3390/ijms13032717