PCR and Magnetic Bead-Mediated Target Capture for the Isolation of Short Interspersed Nucleotide Elements in Fishes


{kind=link}
{kind=link}
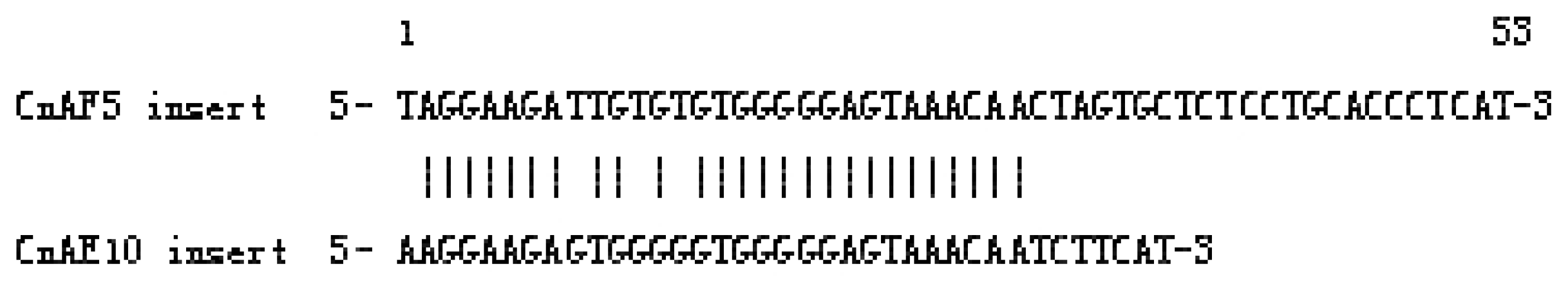{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
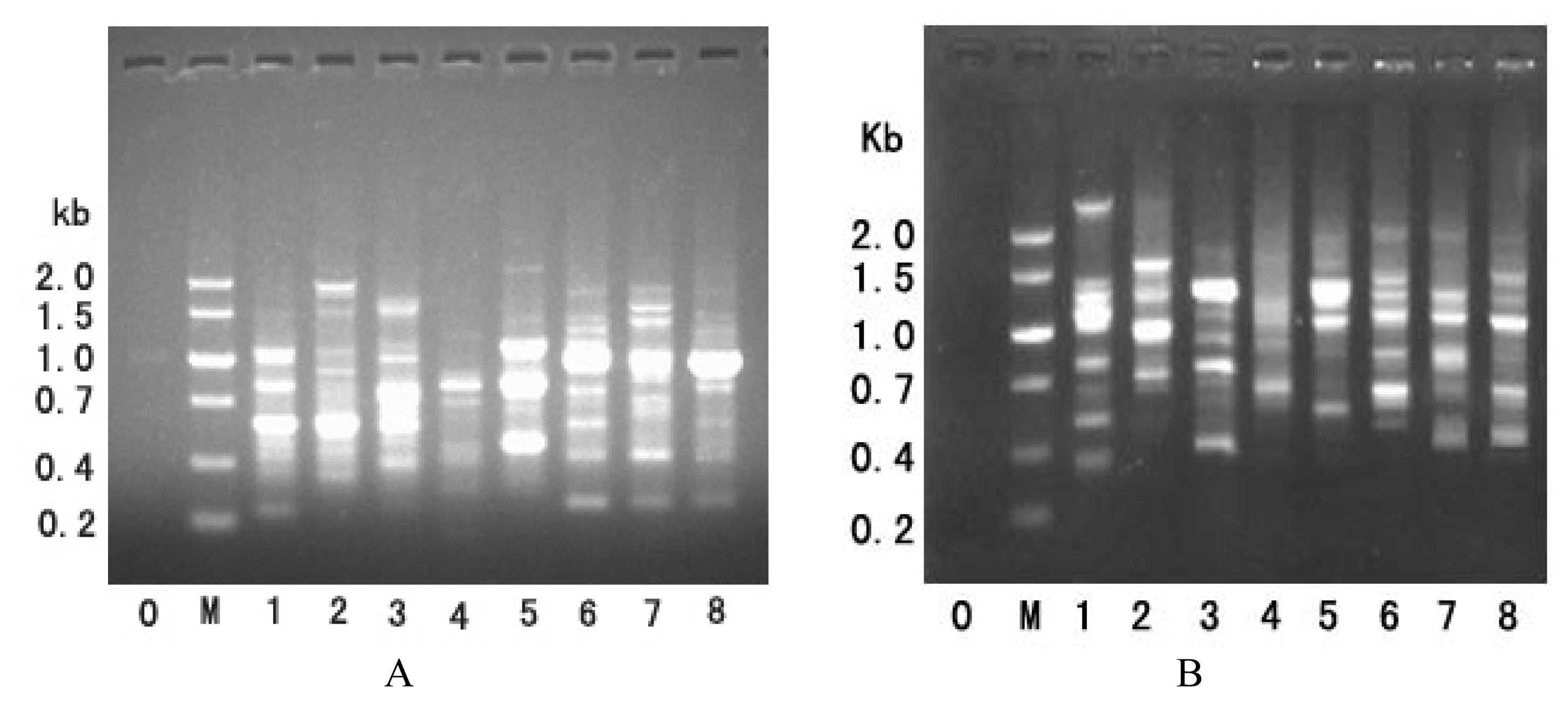
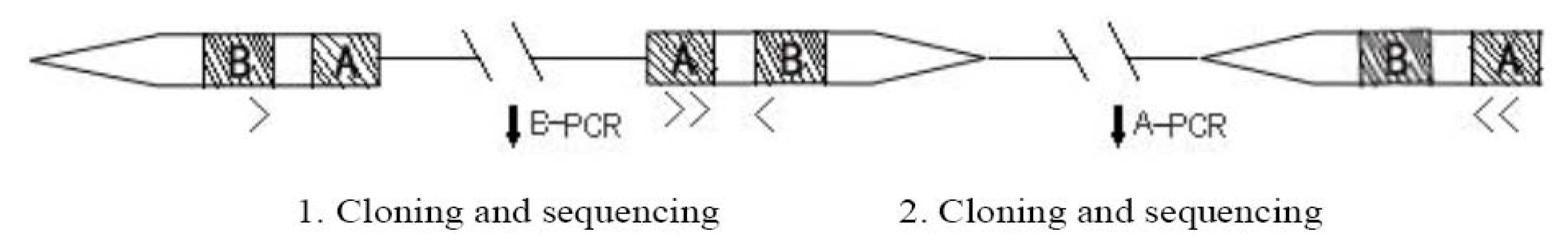
2.1.1. B-PCR
2.1.2. A-PCR
2.1.3. Isolation of SINEs from the C. nasus Genome
2.2. Discussion
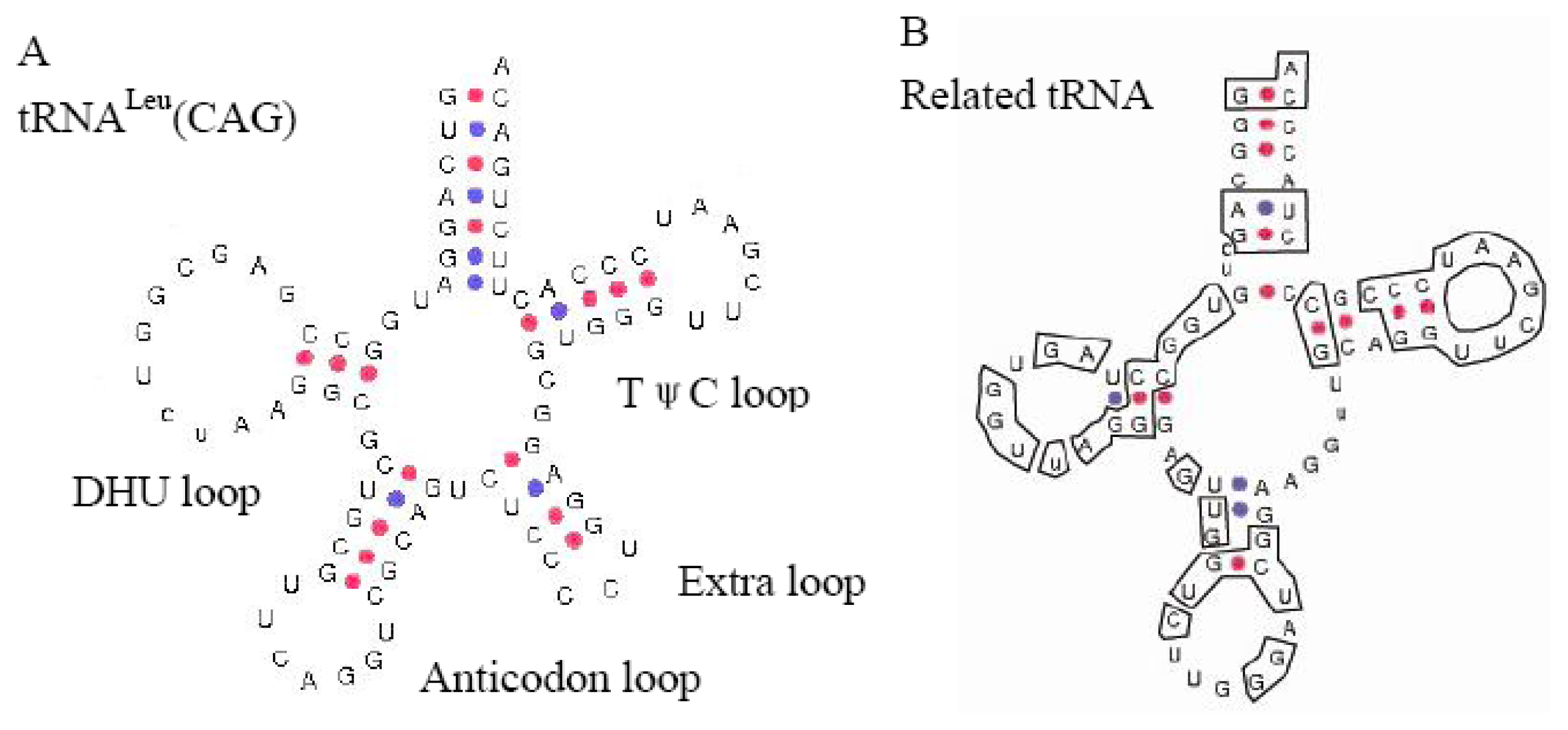
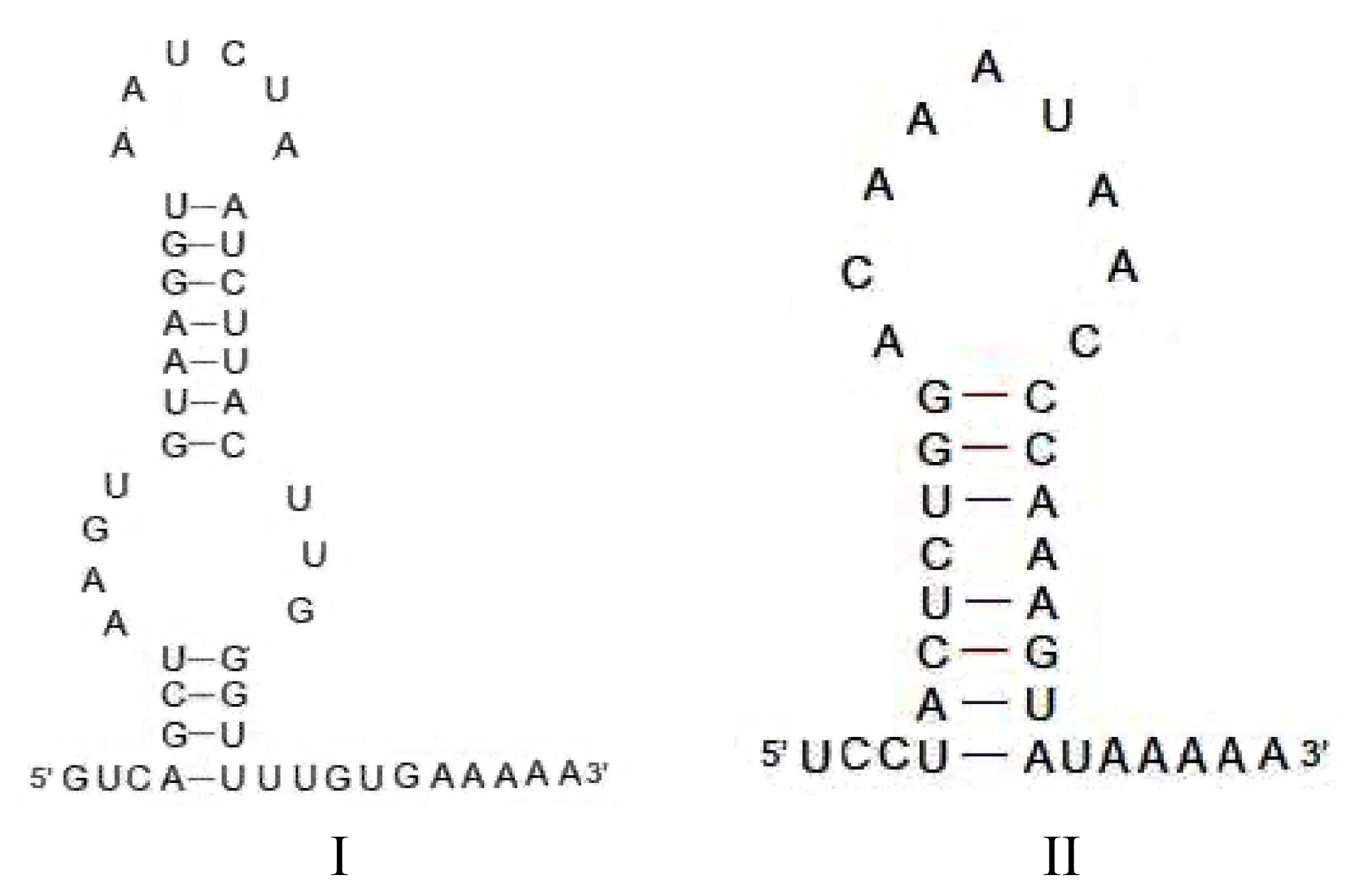
2.2.1. Structure-Based PCR for Capturing New SINEs
2.2.2. Origin of SINEs Isolated from C. nasus
3. Experimental Section
3.1. Fish Samples and DNA Extraction
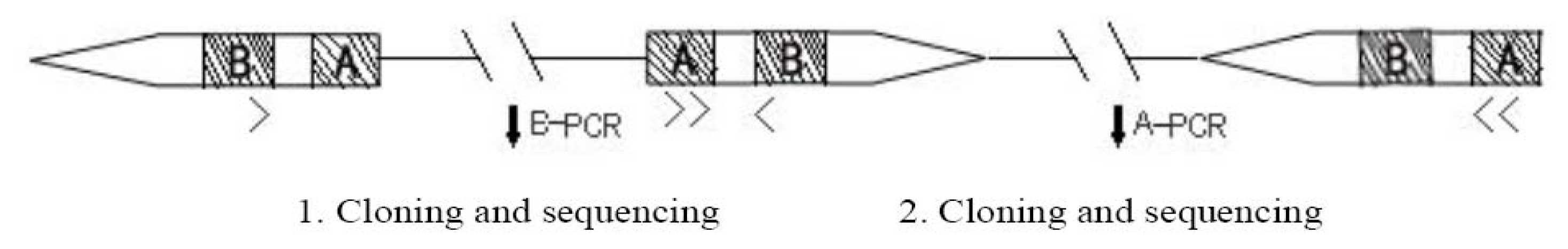
3.2. Design of B(A)-PCR, Cloning and Sequencing
3.3. Genomic DNA Enrichment
3.3.1. Restriction Digestion of Genomic DNA
3.3.2. Ligation of DNA Fragments to the Adaptor
3.3.3. PCR Amplification of Genomic DNA
3.4. Magnetic Particle Isolation of SINEs
3.4.1. Probe
3.4.2. Isolation of Target Sequences
3.4.3. Adaptor PCR, Cloning and Sequencing
4. Conclusions
Acknowledgements
References
- Liu, D.; Tang, W.Q.; Yang, J.Q.; Liu, S.J. Recent advancements in Tc1-like transposons. Sci. Sinica Vitae 2011, 41, 87–96. [Google Scholar]
- Kramerov, D.; Vassetzky, N. Origin and evolution of SINEs in eukaryotic genomes. Heredity 2011, 43, 1–9. [Google Scholar]
- Walters-Conte, K.B.; Johnson, D.L.E.; Allard, M.W.; Pecon-Slattery, J. Carnivore-specific SINEs (Can-SINEs): Distribution, evolution, and genomic impact. J. Hered 2011, 102, S2–S10. [Google Scholar]
- Kapitonov, V.V.; Jurka, J. A novel class of SINE elements derived from 5S rRNA. Mol. Biol. Evol 2003, 20, 694–702. [Google Scholar]
- Kramerov, D.A.; Vassetzky, N.S. SINEs. Wiley Interdiscip. Rev. RNA 2011, 2, 772–786. [Google Scholar]
- Okada, N.; Hamada, M.; Ogiwara, I.; Ohshima, K. SINEs and LINEs share common 3′ sequences: A review. Gene 1997, 205, 229–243. [Google Scholar]
- Shimamura, M.; Abe, H.; Nikaido, M.; Ohshima, K.; Okada, N. Genealogy of families of SINEs in cetaceans and artiodactyls: the presence of a huge superfamily of tRNA (Glu)-derived families of SINEs. Mol. Biol. Evol 1999, 16, 1046–1060. [Google Scholar]
- Xu, J.S.; Liu, T.; Li, D.; Zhang, Z.; Xia, Q.Y.; Zhou, Z.Y. BmSE, a SINE family with 3′ ends of (ATTT) repeats in domesticated silkworm (Bombyx mori). J. Genet. Genomics 2010, 37, 125–135. [Google Scholar]
- Kass, D.H.; Schaetz, B.A.; Beitler, L.; Bonney, K.M.; Jamison, N.; Wiesner, C. Guinea pig ID-like families of SINEs. Gene 2009, 436, 23–29. [Google Scholar]
- Tong, C.B.; Gan, X.N.; He, S.P. Multiple source genes of HAmo SINE actively expanded and ongoing retroposition in cyprinid genomes relying on its partner LINE. BMC Evol. Biol 2010, 10, 115. [Google Scholar]
- Hamada, M.; Kido, Y.; Himberg, M.; Reist, J.D.; Ying, C.; Hasegawa, M.; Okada, N. A newly isolated family of short interspersed repetitive elements (SINEs) in coregonid fishes (whitefish) with sequences that are almost identical to those of the SmaI family of repeats: possible evidence for the horizontal transfer of SINEs. Genetics 1997, 146, 355–367. [Google Scholar]
- Borodulina, O.R.; Kramerov, D.A. Wide distribution of short interspersed elements among eukaryotic genomes. FEBS Lett 1999, 457, 409–413. [Google Scholar]
- Borodulina, O.R.; Kramerov, D.A. PCR-based approach to SINE isolation: Simple and complex SINEs. Gene 2005, 349, 197–205. [Google Scholar]
- Tong, C.B.; Guo, B.C.; He, S.P. Bead-probe complex capture a couple of SINE and LINE family from genomes of two closely related species of East Asian cyprinid directly using magnetic separation. BMC Genomics 2009, 10, 83. [Google Scholar]
- Ogiwara, I.; Miya, M.; Ohshima, K.; Okada, N. V-SINEs: A new superfamily of vertebrate SINEs that are widespread in vertebrate genomes and retain a strongly conserved segment within each repetitive unit. Genome Res 2002, 12, 316–324. [Google Scholar]
- Takahashi, K.; Terai, Y.; Nishida, M.; Okada, N. A novel family of short interspersed repetitive elements (SINEs) from cichlids: the patterns of insertion of SINEs at orthologous loci support the proposed monophyly of four major groups of cichlid fishes in Lake Tanganyika. Mol. Biol. Evol 1998, 15, 391–407. [Google Scholar]
- Fantaccione, S.; Woodrow, P.; Pontecorvo, G. Identification of a family of SINEs and LINEs in the Pipistrellus kuhli genome: A new structural and functional symbiotic relationship. Genomics 2008, 91, 178–185. [Google Scholar]
- Han, Y.W.; He, S.P. Identifying novel SINEs from Cyprinidae and their proliferation mechanism in genome. Prog. Nat. Sci 2007, 17, 270–279. [Google Scholar]
- National Center of Biotechnology Information. Available online: http://www.ncbi.nlm.nih.gov accessed on 9 October 2011.
- Galli, G.; Hofstetter, H.; Birnstiel, M.L. Two conserved sequence blocks within eukaryotic tRNA genes are major promoter elements. Nature 1981, 294, 626–631. [Google Scholar]
- Kido, Y.; Saitoh, M.; Murata, S.; Okada, N. Evolution of the active sequences of the HpaI short interspersed elements. J. Mol. Evol 1995, 41, 986–995. [Google Scholar]
- Schattner, P.; Brooks, A.N.; Lowe, T.M. The tRNAscan-SE, snoscan and snoGPS web servers for the detection of tRNAs and snoRNAs. Nucleic Acids Res 2005, 33, W686–W689. [Google Scholar]
- Sharp, S.; DeFranco, D.; Dingermann, T.; Farrell, P.; Söll, D. Internal control regions for transcription of eukaryotic tRNA genes. Proc. Natl. Acad. Sci. USA 1981, 78, 6657–6661. [Google Scholar]
- Matveev, V.; Okada, N. Retroposons of salmonoid fishes (Actinopterygii: Salmonoidei) and their evolution. Gene 2009, 434, 16–28. [Google Scholar]
- Kido, Y.; Aono, M.; Yamaki, T.K.; Murata, S.; Saneyoshi, M.; Okada, N. Shaping and reshaping of salmonid genomes by amplification of tRNA-derived retroposons during evolution. Proc. Natl. Acad. Sci. USA 1991, 88, 2326–2330. [Google Scholar]
- Nishihara, H.; Smit, A.F.A.; Okada, N. Functional noncoding sequences derived from SINEs in the mammalian genome. Genome Res 2006, 16, 864–874. [Google Scholar]
- Kazazian, H.H., Jr. Mobile elements: drivers of genome evolution. Science 2004, 303, 1626–1632. [Google Scholar]
- Kajikawa, M.; Okada, N. LINEs mobilize SINEs in the eel through a shared 3′ sequence. Cell 2002, 111, 433–444. [Google Scholar]
- Zhang, S.Y. (Ed.) Fauna Sinica, Osteichthyes: Acipenseriformes, Elopiformes, Clupeiformes, Gonorhynchiformes (in Chinese); Science Press: Beijing, China, 2001; pp. 148–154.
- Fauna Sinica, Class Teleostei, Siluriformes (in Chinese); Chu, X.L.; Zhen, B.S.; Dai, D.Y. (Eds.) Science Press: Beijing, China, 1999; pp. 35–77.
- Fauna Sinica, Ostichthyes, Perciformes (V), Gobioidei (in Chinese); Wu, H.L.; Zhong, J.S. (Eds.) Science Press: Beijing, China, 2008; pp. 471–550.
- Lalitha, S. Primer premier 5. Biotech. Softw. Internet Rep 2000, 1, 270–272. [Google Scholar]

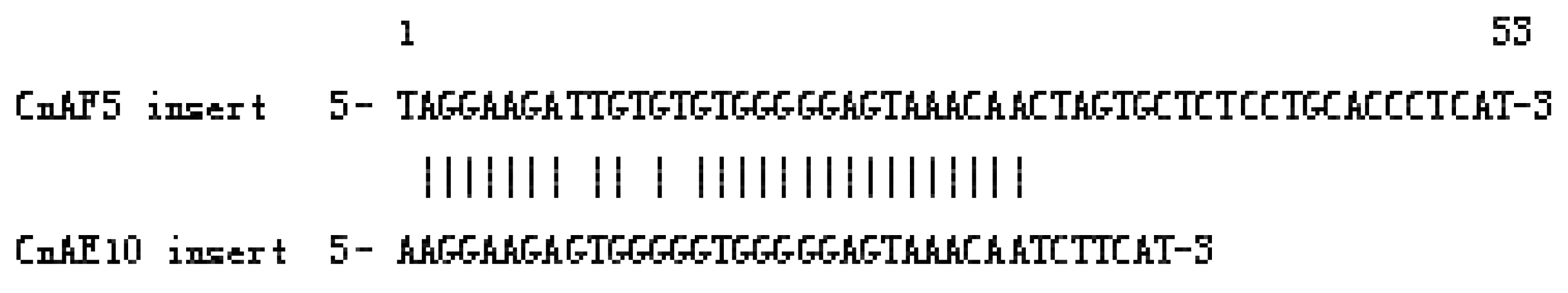

© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, D.; Zhu, G.; Tang, W.; Yang, J.; Guo, H. PCR and Magnetic Bead-Mediated Target Capture for the Isolation of Short Interspersed Nucleotide Elements in Fishes. Int. J. Mol. Sci. 2012, 13, 2048-2062. https://doi.org/10.3390/ijms13022048
Liu D, Zhu G, Tang W, Yang J, Guo H. PCR and Magnetic Bead-Mediated Target Capture for the Isolation of Short Interspersed Nucleotide Elements in Fishes. International Journal of Molecular Sciences. 2012; 13(2):2048-2062. https://doi.org/10.3390/ijms13022048
Chicago/Turabian StyleLiu, Dong, Guoli Zhu, Wenqiao Tang, Jinquan Yang, and Hongyi Guo. 2012. "PCR and Magnetic Bead-Mediated Target Capture for the Isolation of Short Interspersed Nucleotide Elements in Fishes" International Journal of Molecular Sciences 13, no. 2: 2048-2062. https://doi.org/10.3390/ijms13022048