Comparative Effects of Coenzyme Q10 or n-3 Polyunsaturated Fatty Acid Supplementation on Retinal Angiogenesis in a Rat Model of Oxygen-Induced Retinopathy

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Design
2.3. Neonatal Intermittent Hypoxia (IH) Profiles
2.4. Sample Collection and Processing
2.5. Assay of Ocular Growth Factors
2.6. Total Cellular Protein Levels
2.7. Retinal Flatmounts
2.8. ADPase Staining of the Retinas
2.9. GFAP and Isolectin B4 Staining
2.10. Vascular Density Quantification
2.11. Retinal Scoring
2.12. Statistical Analysis
3. Results
3.1. Eye-Opening at P14
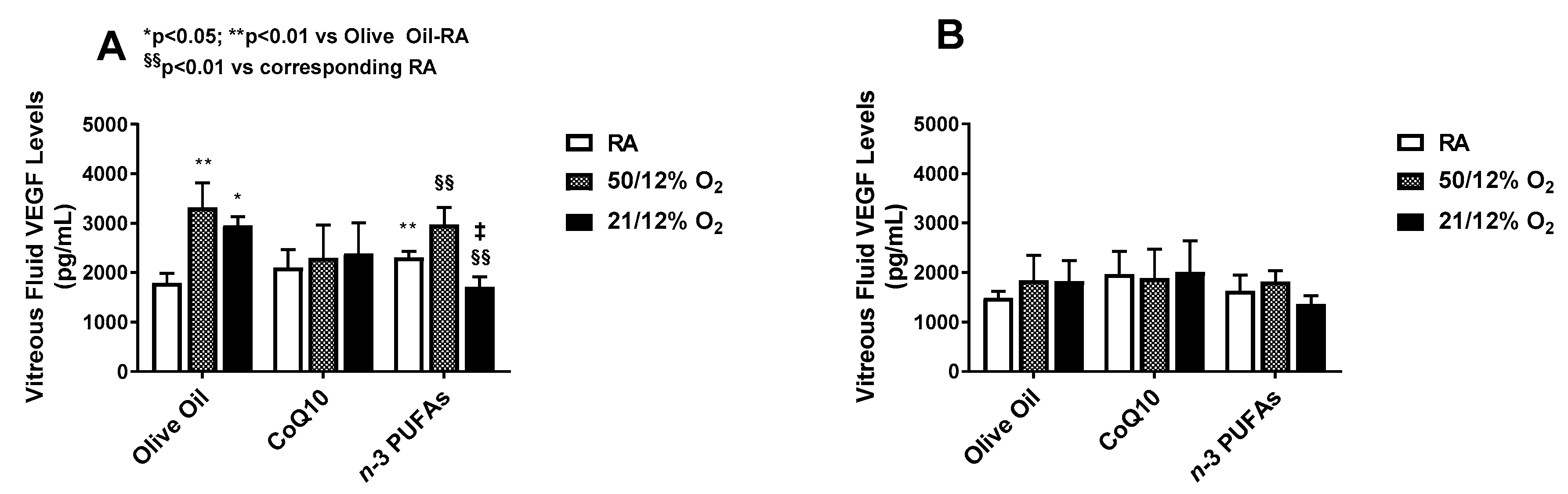
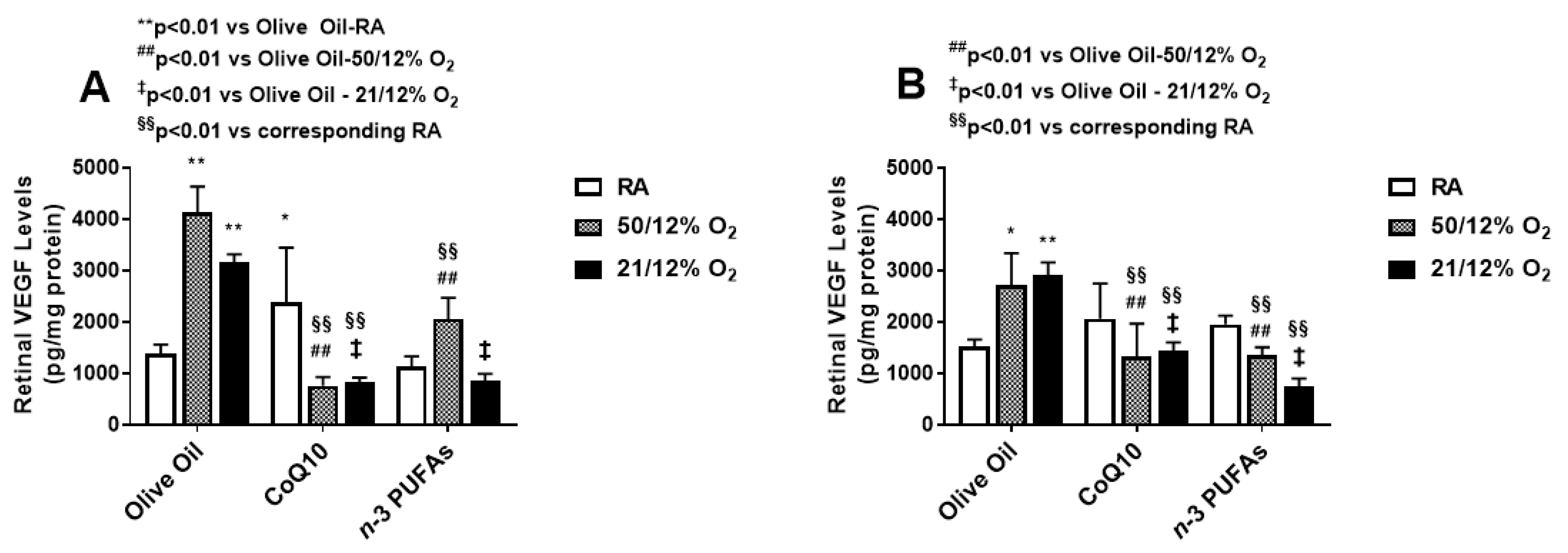
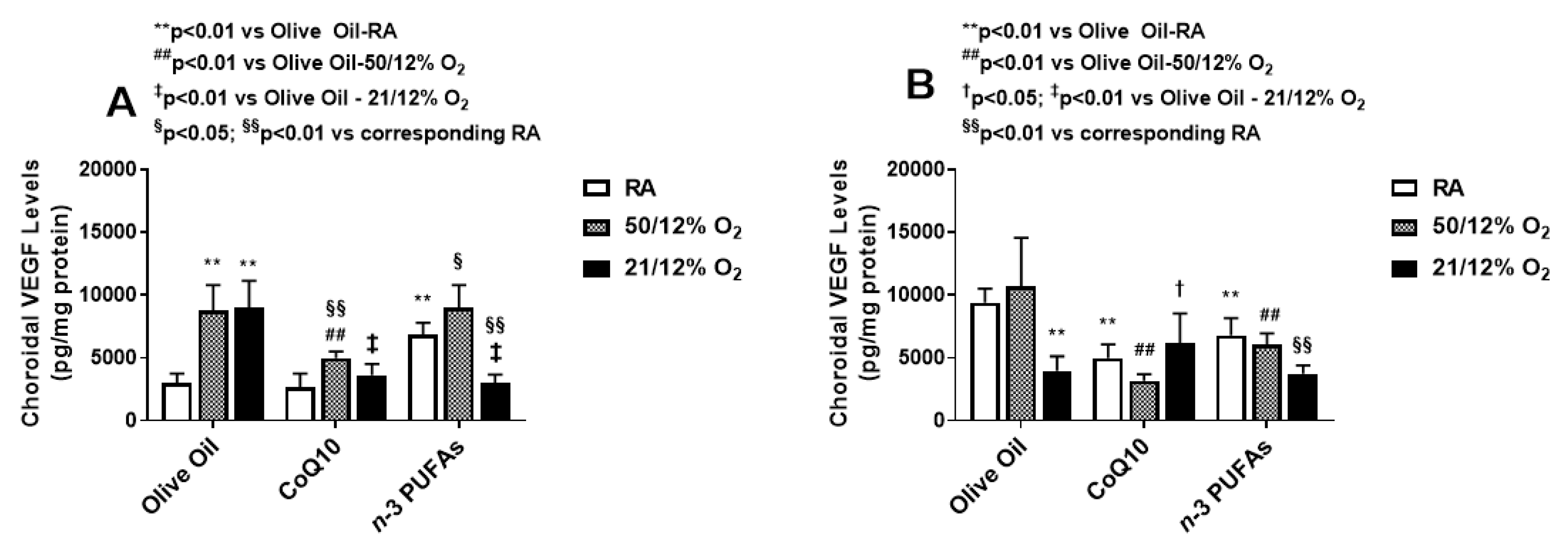
3.2. VEGF Levels
3.3. Ocular sVEGFR-1
3.4. Ocular IGF-I
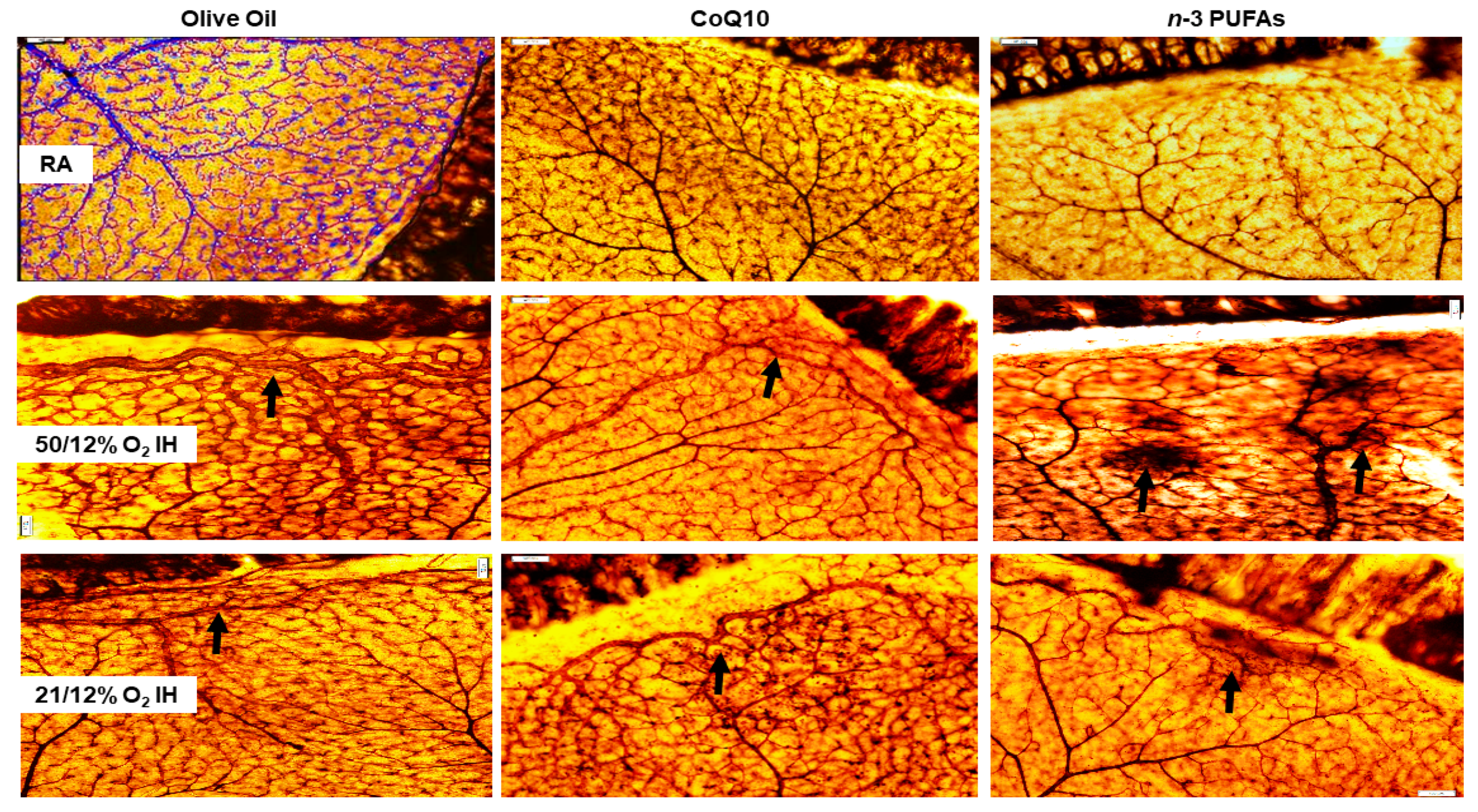
3.5. Retinal Angiogenesis
3.6. Retinal Scoring
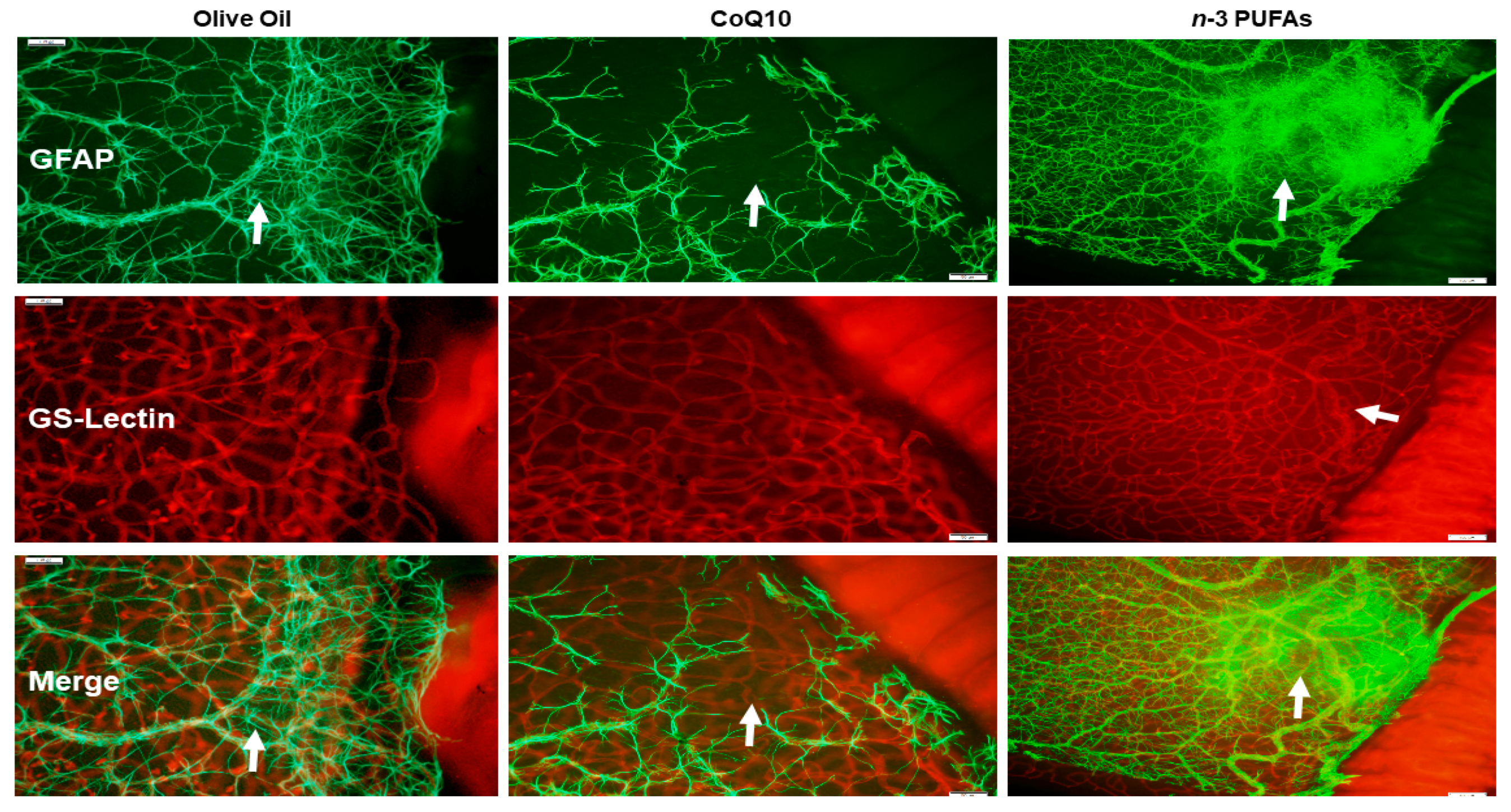
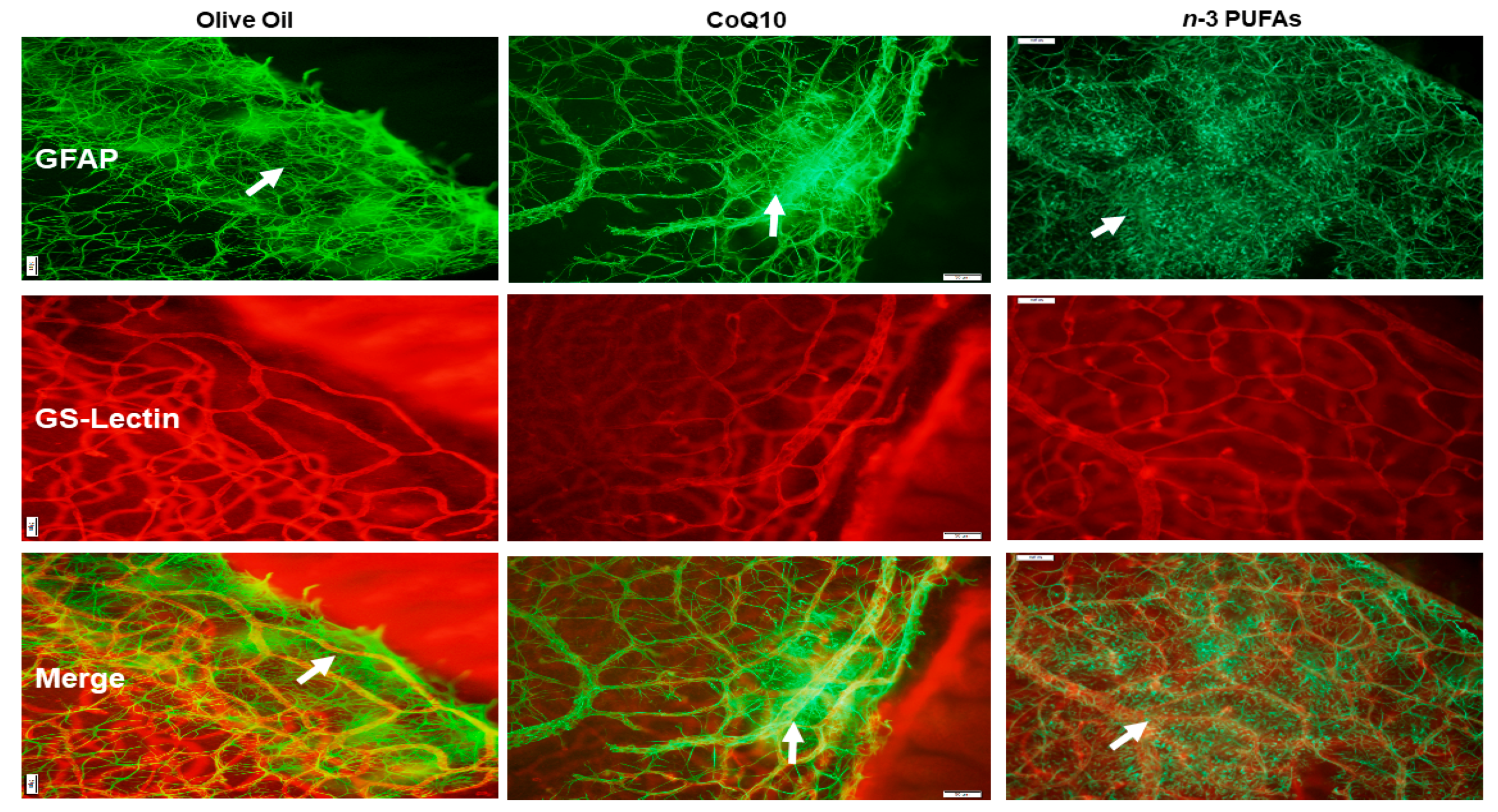
3.7. Astrocytic Template
3.8. Retinal Vascular Quantification
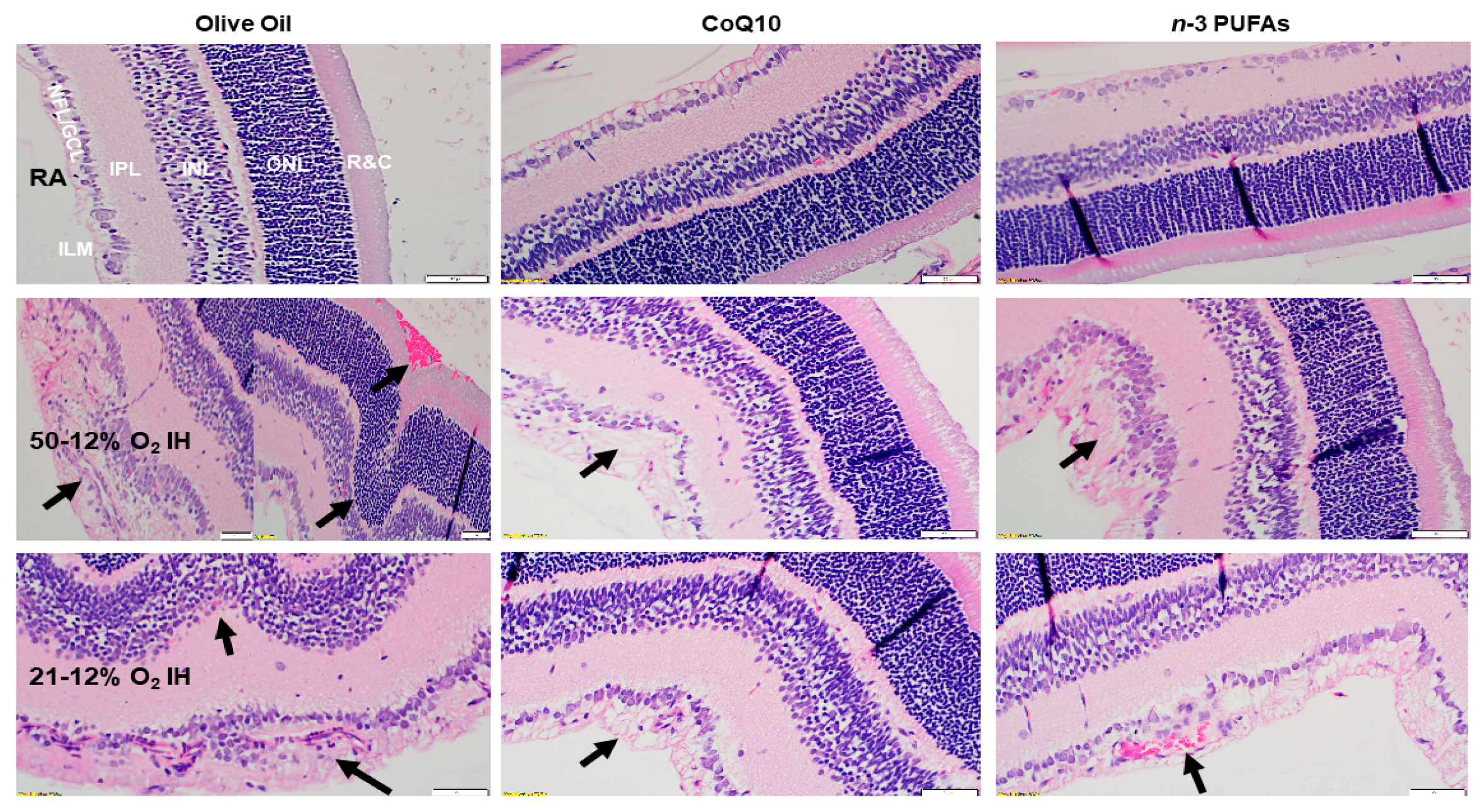
3.9. Retinal Histopathology
3.10. Retinal Morphometry
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kong, L.; Fry, M.; Al-Samarraie, M.; Gilbert, C.; Steinkuller, P.G. An update on progress and the changing epidemiology of causes of childhood blindness worldwide. J. Am. Assoc. Pediatr. Ophthalmol. Strabismus 2012, 16, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.T.; Tasman, W.S. Retinopathy of prematurity: Late complications in the baby boomer generation (1946–1964). Trans. Am. Ophthalmol. Soc. 2005, 103, 225–234. [Google Scholar] [PubMed]
- Baker, P.S.; Tasman, W. Myopia in adults with retinopathy of prematurity. Am. J. Ophthalmol. 2008, 145, 1090–1094. [Google Scholar] [CrossRef] [PubMed]
- Rothschild, M.I.; Russ, R.; Brennan, K.A.; Williams, C.J.; Berrones, D.; Patel, B.; Martinez-Castellanos, M.A.; Fernandes, A.; Hubbard, G.B., 3rd; Chan, R.V.P.; et al. The Economic Model of Retinopathy of Prematurity (EcROP) Screening and Treatment: Mexico and the United States. Am. J. Ophthalmol. 2016, 168, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Dave, H.B.; Gordillo, L.; Yang, Z.; Zhang, M.S.; Hubbard, G.B., 3rd; Olsen, T.W. The societal burden of blindness secondary to retinopathy of prematurity in Lima, Peru. Am. J. Ophthalmol. 2012, 154, 750–755. [Google Scholar] [CrossRef] [PubMed]
- Eckert, K.A.; Carter, M.J.; Lansingh, V.C.; Wilson, D.A.; Furtado, J.M.; Frick, K.D.; Resnikoff, S. A simple method for estimating the economic cost of productivity loss due to blindness and moderate to severe visual impairment. Ophthalmic Epidemiol. 2015, 22, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Frick, K.D.; Foster, A. The magnitude and cost of global blindness: An increasing problem that can be alleviated. Am. J. Ophthalmol. 2003, 135, 471–476. [Google Scholar] [CrossRef]
- Smith, L.E. Pathogenesis of Retinopathy of Prematurity. Growth Horm. IGF Res. 2004, 14, S140–S144. [Google Scholar] [CrossRef] [PubMed]
- Cringle, S.J.; Yu, D.Y. Oxygen supply and consumption in the retina: Implications for studies of retinopathy of prematurity. Doc. Ophthalmol. 2010, 120, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Phelps, D.L. Retinopathy of prematurity: History, classification, and pathophysiology. NeoReviews 2001, 2, e153–e166. [Google Scholar] [CrossRef]
- Phelps, D.L. Retinopathy of prematurity. In Neonatal Perinatal Medicine: Diseases of the Fetus and Infant, 7th ed.; Fanaroff, A.A., Martin, R.J., Eds.; Mosby: St. Louis, MO, USA, 2002; Volume 2, pp. 1595–1599. [Google Scholar]
- Rhein, L.M.; Dobson, N.R.; Darnall, R.A.; Corwin, M.J.; Heeren, T.C.; Poets, C.F.; McEntire, B.L.; Hunt, C.E.; Caffeine Pilot Study Group. Effects of caffeine on intermittent hypoxia in infants born prematurely: A randomized clinical trial. JAMA Pediatr. 2014, 168, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Di Fiore, J.M.; Martin, R.J.; Gauda, E.B. Apnea of prematurity-perfect storm. Respir. Physiol. Neurobiol. 2013, 189, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Gonzalez, F.; Mu, D. Apnea of prematurity: From cause to treatment. Eur. J. Pediatr. 2011, 170, 1097–1105. [Google Scholar] [CrossRef] [PubMed]
- Stokowski, L.A. A primer on Apnea of prematurity. Adv. Neonatal Care 2005, 5, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.J.; Di Fiore, J.M.; Macfarlane, P.M.; Wilson, C.G. Physiologic basis for intermittent hypoxic episodes in preterm infants. Adv. Exp. Med. Biol. 2012, 758, 351–358. [Google Scholar] [CrossRef] [PubMed]
- York, J.R.; Landers, S.; Kirby, R.S.; Arbogast, P.G.; Penn, J.S. Arterial oxygen fluctuation and retinopathy of prematurity in very-low-birth-weight infants. J. Perinatol. 2004, 24, 82–87. [Google Scholar] [CrossRef] [PubMed]
- DiFiore, J.M.; Bloom, J.N.; Orge, F.; Schutt, A.; Schluchter, M.; Cheruvu, V.K.; Walsh, M.; Finer, N.; Martin, R.J. A higher incidence of intermittent hypoxemic episodes is associated with severe retinopathy of prematurity. J. Pediatr. 2010, 157, 69–73. [Google Scholar] [CrossRef] [PubMed]
- DiFiore, J.M.; Kaffashi, F.; Loparo, K.; Sattar, A.; Schluchter, M.; Foglyano, R.; Martin, R.J.; Wilson, C.G. The relationship between patterns of intermittent hypoxia and retinopathy of prematurity in preterm infants. Pediatr. Res. 2012, 72, 606–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, C.G.; Benitz, W.E.; Madan, A. Retinopathy of prematurity and pulse oximetry: A national survey of recent practices. J. Perinatol. 2004, 24, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Castillo, A.; Deulofeut, R.; Critz, A.; Sola, A. Prevention of retinopathy of prematurity in preterm infants through changes in clinical practice and SpO2 technology. Acta Paediatr. 2011, 100, 188–192. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, S.; Fleck, B.W.; Elton, R.A.; McIntosh, N. Transcutaneous oxygen levels in retinopathy of prematurity. Lancet 1995, 346, 1464–1465. [Google Scholar] [CrossRef]
- Saito, Y.; Omoto, T.; Cho, Y.; Hatsukawa, Y.; Fujimura, M.; Takeuchi, T. The progression of retinopathy of prematurity and fluctuation in blood gas tension. Graefe’s Arch. Clin. Exp. Ophthalmol. 1993, 231, 151–156. [Google Scholar] [CrossRef]
- Di Fiore, J.M.; Martin, R.J.; Li, H.; Morris, N.; Carlo, W.A.; Finer, N.; Walsh, M. SUPPORT Study Group of the Eunice Kennedy Shriver National Institute of Child Health, and Human Development Neonatal Research Network. Patterns of Oxygenation, Mortality, and Growth Status in the Surfactant Positive Pressure and Oxygen Trial Cohort. J. Pediatr. 2017, 186, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.R.; Dasilva, D.A.; Cluette-Brown, J.E.; Dimonda, C.; Hamill, A.; Bhutta, A.Q.; Coronel, E.; Wilschanski, M.; Stephens, A.J.; Driscoll, D.F.; et al. Decreased postnatal docosahexaenoic and arachidonic acid blood levels in premature infants are associated with neonatal morbidities. J. Pediatr. 2011, 159, 743–749. [Google Scholar] [CrossRef] [PubMed]
- Bernhard, W.; Raith, M.; Koch, V.; Kunze, R.; Maas, C.; Abele, H.; Poets, C.F.; Franz, A.R. Plasma phospholipids indicate impaired fatty acid homeostasis in preterm infants. Eur. J. Nutr. 2014, 53, 1533–1547. [Google Scholar] [CrossRef] [PubMed]
- Lapillonne, A.; Jensen, C.L. Reevaluation of the DHA requirement for the premature infant. Prostaglandins Leukot. Essent. Fat. Acids 2009, 81, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Lavoie, P.M.; Lacaze-Masmonteil, T.; Rhainds, M.; Marc, I. Omega-3 long chain polyunsaturated fatty acids for extremely preterm infants: A systematic review. Pediatrics 2014, 134, 120–134. [Google Scholar] [CrossRef] [PubMed]
- Lapillonne, A.; Moltu, S.J. Long-Chain Polyunsaturated Fatty Acids and Clinical Outcomes of Preterm Infants. Ann. Nutr. Metab. 2016, 69 (Suppl. 1), 35–44. [Google Scholar] [CrossRef] [PubMed]
- Richard, D.; Kefi, K.; Barbe, U.; Bausero, P.; Visioli, F. Polyunsaturated fatty acids as antioxidants. Pharmacol. Res. 2008, 57, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Mansfield, K.D.; Guzy, R.D.; Pan, Y.; Young, R.M.; Cash, T.P.; Schumacker, P.T.; Simon, M.C. Mitochondrial dysfunction resulting from loss of cytochrome c impairs cellular oxygen sensing and hypoxic HIF-alpha activation. Cell Metab. 2005, 1, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Semenza, G.L. Oxygen-dependent regulation of mitochondrial respiration by hypoxia-inducible factor 1. Biochem. J. 2007, 405, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, D.Y.; Cringle, S.J. Oxygen distribution and consumption within the retina in vascularised and avascular retinas and in animal models of retinal disease. Prog. Retin. Eye Res. 2001, 20, 175–208. [Google Scholar] [CrossRef]
- Aberg, F.; Appelkvist, E.L.; Dallner, G.; Ernster, L. Distribution and redox state of ubiquinones in rat and human tissues. Arch. Biochem. Biophys. 1992, 295, 230–234. [Google Scholar] [CrossRef]
- Sarmiento, A.; Diaz-Castro, J.; Pulido-Moran, M.; Kajarabille, N.; Guisado, R.; Ochoa, J.J. Coenzyme Q10 supplementation and exercise in healthy humans: A systematic review. Curr. Drug Metab. 2016, 17, 345–358. [Google Scholar] [CrossRef] [PubMed]
- Fitch, C.D.; Folkers, K. Coenzyme Q and the stability of biological membranes. Biochem. Biophys. Res. Commun. 1967, 26, 128–231. [Google Scholar] [CrossRef]
- Garrido-Maraver, J.; Cordero, M.D.; Oropesa-Avila, M.; Vega, A.F.; de la Mata, M.; Pavon, A.D.; Alcocer-Gomez, E.; Calero, C.P.; Paz, M.V.; Alanis, M.; et al. Clinical applications of coenzyme Q10. Front. Biosci. 2014, 19, 619–633. [Google Scholar] [CrossRef]
- Lee, D.; Kim, K.Y.; Shim, M.S.; Kim, S.Y.; Ellisman, M.H.; Weinreb, R.N.; Ju, W.K. Coenzyme Q10 ameliorates oxidative stress and prevents mitochondrial alteration in ischemic retinal injury. Apoptosis 2014, 19, 603–614. [Google Scholar] [CrossRef] [PubMed]
- Bergamini, C.; Moruzzi, N.; Sblendido, A.; Lenaz, G.; Fato, R. A water soluble CoQ10 formulation improves intracellular distribution and promotes mitochondrial respiration in cultured cells. PLoS ONE 2012, 7, e33712. [Google Scholar] [CrossRef] [PubMed]
- Beharry, K.D.; Cai, C.L.; Henry, M.M.; Chowdhury, S.; Valencia, G.B.; Aranda, J.V. Co-Enzyme Q10 and n-3 polyunsaturated fatty acid supplementation reverse intermittent hypoxia-induced growth restriction and improved antioxidant profiles in neonatal rats. Antioxidants 2017, 6, 103. [Google Scholar] [CrossRef] [PubMed]
- Aranda, J.V.; Cai, C.L.; Ahmad, T.; Bronshtein, V.; Sadeh, J.; Valencia, G.B.; Lazzaro, D.R.; Beharry, K.D. Pharmacologic synergism of ocular ketorolac and systemic caffeine citrate in rat oxygen-induced retinopathy. Pediatr. Res. 2016, 80, 554–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, C.; Beharry, K.D.; Shen, X.; Li, J.; Wang, L.; Aranda, J.V.; Qu, J. Proteomic profiling of the retinas in a neonatal rat model of oxygen-induced retinopathy with a reproducible ion-current-based MS1 approach. J. Proteome Res. 2015, 14, 2109–2120. [Google Scholar] [CrossRef] [PubMed]
- Beharry, K.D.; Cai, C.L.; Sharma, P.; Bronshtein, V.; Valencia, G.B.; Lazzaro, D.R.; Aranda, J.V. Hydrogen peroxide accumulation in the choroid during intermittent hypoxia increases risk of severe oxygen-induced retinopathy in neonatal rats. Investig. Ophthalmol. Vis. Sci. 2013, 54, 7644–7657. [Google Scholar] [CrossRef] [PubMed]
- Beharry, K.D.; Cai, C.L.; Skelton, J.; Siddiqui, F.; D’Agrosa, C.; Calo, J.; Valencia, G.B.; Aranda, J.V. Oxygen-induced retinopathy from recurrent intermittent hypoxia is not dependent on resolution with room air or oxygen, in neonatal rats. Int. J. Mol. Sci. 2018, 19, 1337. [Google Scholar] [CrossRef] [PubMed]
- Beharry, K.D.; Cai, C.L.; Ahmad, T.; Guzel, S.; Valencia, G.B.; Aranda, J.V. Impact of chronic neonatal intermittent hypoxia on severity of retinal damage in a rat model of oxygen-induced retinopathy. J. Nat. Sci. 2018, 4, e488. [Google Scholar] [PubMed]
- Tan, J.J.; Cai, C.L.; Shrier, E.M.; McNally, L.; Lazzaro, D.R.; Aranda, J.V.; Beharry, K.D. Ocular adverse effects of intravitreal bevacizumab are potentiated by intermittent hypoxia in a rat model of oxygen-induced retinopathy. J. Ophthalmol. 2017, 2017, 4353129. [Google Scholar] [CrossRef] [PubMed]
- Nakano, A.; Nakahara, T.; Mori, A.; Ushikubo, H.; Sakamoto, K.; Ishii, K. Short-term treatment with VEGF receptor inhibitors induces retinopathy of prematurity-like abnormal vascular growth in neonatal rats. Exp. Eye Res. 2016, 143, 120–131. [Google Scholar] [CrossRef] [PubMed]
- Higgins, R.D.; Yu, K.; Sanders, R.J.; Nandgaonkar, B.N.; Rotschild, T.; Rifkin, D.B. Diltiazem reduces retinal neovascularization in a mouse model of oxygen induced retinopathy. Curr. Eye Res. 1999, 18, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Binnes, K.E.; Salt, T.E. Post eye-opening maturation of visual receptive field diameters in superior colliculus of normal- and dark-reared rats. Brain Res. Dev. Brain Res. 1997, 99, 263–266. [Google Scholar] [CrossRef]
- Robinson, S.R.; Dreher, B. The visual pathways of eutherian mammals and marsupials develop according to a common timetable. Brain Behav. Evol. 1990, 36, 177–195. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, D.R.; Theuer, R.C.; Castañeda, Y.S.; Wheaton, D.H.; Bosworth, R.G.; O’Connor, A.R.; Morale, S.E.; Wiedemann, L.E.; Birch, E.E. Maturation of visual acuity is accelerated in breast-fed term infants fed baby food containing DHA-enriched egg yolk. J. Nutr. 2004, 134, 2307–2313. [Google Scholar] [CrossRef] [PubMed]
- Uauy, R.; Hoffman, D.R.; Peirano, P.; Birch, D.G.; Birch, E.E. Essential fatty acids in visual and brain development. Lipids 2001, 36, 885–895. [Google Scholar] [CrossRef] [PubMed]
- Bazan, N.G. Cell survival matters: Docosahexaenoic acid signaling, neuroprotection and photoreceptors. Trends Neurosci. 2006, 29, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Pawlik, D.; Lauterbach, R.; Walczak, M.; Hurkala, J.; Sherman, M.P. Fish oil fat emulsion supplementation reduces the risk of retinopathy in very low birth weight infants: A prospective, randomized study. JPEN J. Parenter. Enter. Nutr. 2014, 38, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Pawlik, D.; Lauterbach, R.; Turyk, E. Fish-oil fat emulsion supplementation may reduce the risk of severe retinopathy in VLBW infants. Pediatrics 2011, 127, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Davis, B.M.; Tian, K.; Pahlitzsch, M.; Brenton, J.; Ravindran, N.; Butt, G.; Malaguarnera, G.; Normando, E.M.; Guo, L.; Cordeiro, M.F. Topical Coenzyme Q10 demonstrates mitochondrial-mediated neuroprotection in a rodent model of ocular hypertension. Mitochondrion 2017, 36, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Lulli, M.; Witort, E.; Papucci, L.; Torre, E.; Schipani, C.; Bergamini, C.; Dal Monte, M.; Capaccioli, S. Coenzyme Q10 instilled as eye drops on the cornea reaches the retina and protects retinal layers from apoptosis in a mouse model of kainate-induced retinal damage. Investig. Ophthalmol. Vis. Sci. 2012, 53, 8295–8302. [Google Scholar] [CrossRef] [PubMed]
- Nucci, C.; Tartaglione, R.; Cerulli, A.; Mancino, R.; Spanò, A.; Cavaliere, F.; Rombolà, L.; Bagetta, G.; Corasaniti, M.T.; Morrone, L.A. Retinal damage caused by high intraocular pressure-induced transient ischemia is prevented by coenzyme Q10 in rat. Int. Rev. Neurobiol. 2007, 82, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Shindou, H.; Koso, H.; Sasaki, J.; Nakanishi, H.; Sagara, H.; Nakagawa, K.M.; Takahashi, Y.; Hishikawa, D.; Iizuka-Hishikawa, Y.; Tokumasu, F.; et al. Docosahexaenoic acid preserves visual function by maintaining correct disc morphology in retinal photoreceptor cells. J. Biol. Chem. 2017, 292, 12054–12064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, J.; Kaufman, Y.; Washington, I. Coenzyme Q10 in the human retina. Investig. Ophthalmol. Vis. Sci. 2009, 50, 1814–1818. [Google Scholar] [CrossRef] [PubMed]
- Alleva, R.; Tomasetti, M.; Battino, M.; Curatola, G.; Littarru, G.P.; Folkers, K. The roles of coenzyme Q10 and vitamin E on the peroxidation of human low density lipoprotein subfractions. Proc. Natl. Acad. Sci. USA 1995, 92, 9388–9391. [Google Scholar] [CrossRef] [PubMed]
- Ashton, N. Pathological basis of retrolental fibroplasia. Br. J. Ophthalmol. 1954, 38, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Ashton, N.; Cook, C. Direct observation of the effect of oxygen on developing vessels: Preliminary report. Br. J. Ophthalmol. 1954, 38, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Ashton, N.; Ward, B.; Serpell, G. Effect of oxygen on developing retinal vessels with particular reference to the problem of retrolental fibroplasia. Br. J. Ophthalmol. 1954, 38, 397–432. [Google Scholar] [CrossRef] [PubMed]
- Hellström, A.; Smith, L.E.; Dammann, O. Retinopathy of prematurity. Lancet 2013, 382, 1445–1447. [Google Scholar] [CrossRef]
- Smith, L.E. Pathogenesis of retinopathy of prematurity. Semin. Neonatol. 2003, 8, 469–473. [Google Scholar] [CrossRef]
- Chan-Ling, T.; Gock, B.; Stone, J. The effect of oxygen on vasoformative cell division: Evidence that ‘physiological hypoxia’ is the stimulus for normal retinal vasculogenesis. Investig. Ophthalmol. Vis. Sci. 1995, 36, 1201–1214. [Google Scholar]
- Gajos, G.; Zalewski, J.; Rostoff, P.; Nessler, J.; Piwowarska, W.; Undas, A. Reduced thrombin formation and altered fibrin clot properties induced by polyunsaturated omega-3 fatty acids on top of dual antiplatelet therapy in patients undergoing percutaneous coronary intervention (OMEGA-PCI clot). Arterioscler. Thromb. Vasc. Biol. 2011, 31, 1696–1702. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, H.S.; Mulvad, G.; Seidelin, K.N.; Malcom, G.T.; Boudreau, D.A. N-3 fatty acids as a risk factor for haemorrhagic stroke. Lancet 1999, 353, 812–813. [Google Scholar] [CrossRef]
- Modanlou, H.D.; Gharraee, Z.; Hasan, J.; Waltzman, J.; Nageotte, S.; Beharry, K.D. Ontogeny of VEGF, IGF-I, and GH in neonatal rat serum, vitreous fluid, and retina from birth to weaning. Investig. Ophthalmol. Vis. Sci. 2006, 47, 738–744. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, S.; Jiang, F.; You, C.; Mao, C.; Yu, J.; Han, J.; Zhang, Z.; Yan, H. Vitreous and plasma VEGF levels as predictive factors in the progression of proliferative diabetic retinopathy after vitrectomy. PLoS ONE 2014, 9, e110531. [Google Scholar] [CrossRef] [PubMed]
- Barleon, B.; Siemeister, G.; Martiny-Baron, G.; Weindel, K.; Herzog, C.; Marmé, D. Vascular endothelial growth factor up-regulates its receptor fms-like tyrosine kinase 1 (FLT-1) and a soluble variant of FLT-1 in human vascular endothelial cells. Cancer Res. 1997, 57, 5421–5425. [Google Scholar] [PubMed]
- Holash, J.; Davis, S.; Papadopoulos, N.; Croll, S.D.; Ho, L.; Russell, M.; Boland, P.; Leidich, R.; Hylton, D.; Burova, E.; et al. VEGF-Trap: A VEGF blocker with potent antitumor effects. Proc. Natl. Acad. Sci. USA 2002, 99, 11393–11398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, L.; Uehara, H.; Zhang, X.; Das, S.K.; Olsen, T.; Holt, D.; Simonis, J.M.; Jackman, K.; Singh, N.; Miya, T.R.; et al. Photoreceptor avascular privilege is shielded by solubleVEGF receptor-1. Elife 2013, 2, e00324. [Google Scholar] [CrossRef] [PubMed]
- Hellström, A.; Engström, E.; Hård, A.L.; Albertsson-Wikland, K.; Carlsson, B.; Niklasson, A.; Löfqvist, C.; Svensson, E.; Holm, S.; Ewald, U.; et al. Postnatal serum insulin-like growth factor I deficiency is associated with retinopathy of prematurity and other complications of premature birth. Pediatrics 2003, 112, 1016–1020. [Google Scholar]
- Young, L.R.; Kurzer, M.S.; Thomas, W.; Redmon, J.B.; Raatz, S.K. Low-fat diet with omega-3 fatty acids increases plasma insulin-like growth factor concentration in healthy postmenopausal women. Nutr. Res. 2013, 33, 565–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heidary, G.; Vanderveen, D.; Smith, L.E. Retinopathy of prematurity: Current concepts in molecular pathogenesis. Semin. Ophthalmol. 2009, 24, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Stone, J.; Itin, A.; Alon, T.; Pe’er, J.; Gnessin, H.; Chan-Ling, T.; Keshet, E. Development of retinal vasculature is mediated by hypoxia-induced vascular endothelial growth factor (VEGF) expression by neuroglia. J. Neurosci. 1995, 15, 4738–4747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorrell, M.I.; Aguilar, E.; Jacobson, R.; Trauger, S.A.; Friedlander, J.; Siuzdak, G.; Friedlander, M. Maintaining retinal astrocytes normalizes revascularization and prevents vascular pathology associated with oxygen-induced retinopathy. Glia 2010, 58, 43–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan-Ling, T.; McLeod, D.S.; Hughes, S.; Baxter, L.; Chu, Y.; Hasegawa, T.; Lutty, G.A. Astrocyte-endothelial cell relationships during human retinal vascular development. Investig. Ophthalmol. Vis. Sci. 2004, 45, 2020–2032. [Google Scholar] [CrossRef]
- Weidemann, A.; Krohne, T.U.; Aguilar, E.; Kurihara, T.; Takeda, N.; Dorrell, M.I.; Simon, M.C.; Haase, V.H.; Friedlander, M.; Johnson, R.S. Astrocyte hypoxic response is essential for pathological but not developmental angiogenesis of the retina. Glia 2010, 58, 1177–1185. [Google Scholar] [CrossRef] [PubMed]
- Luna, G.; Keeley, P.W.; Reese, B.E.; Linberg, K.A.; Lewis, G.P.; Fisher, S.K. Astrocyte structural reactivity and plasticity in models of retinal detachment. Exp. Eye Res. 2016, 150, 4–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Formichella, C.R.; Abella, S.K.; Sims, S.M.; Cathcart, H.M.; Sappington, R.M. Astrocyte reactivity: A biomarker for retinal ganglion cell health in retinal neurodegeneration. J. Clin. Cell. Immunol. 2014, 5, 188. [Google Scholar] [CrossRef] [PubMed]
- Pekny, M.; Pekna, M. Astrocyte reactivity and reactive astrogliosis: Costs and benefits. Physiol. Rev. 2014, 4, 1077–1098. [Google Scholar] [CrossRef] [PubMed]
- Ashton, N. Oxygen and the growth and development of retinal vessels. Am. J. Ophthalmol. 1966, 62, 412–435. [Google Scholar] [CrossRef]
- Penn, J.S.; Tolman, B.L.; Lowery, L.A.; Koutz, C.A. Oxygen-induced retinopathy in the rat: Hemorrhages and dysplasias may lead to retinal detachment. Curr. Eye Res. 1992, 11, 939–953. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, R.S.; O’Connell, R.; Ascher, S.B.; Sarin, N.; Freedman, S.F.; Wallace, D.K.; Chiu, S.J.; Farsiu, S.; Cotton, M.; Toth, C.A. Spectral-domain optical coherence tomographic assessment of severity of cystoid macular edema in retinopathy of prematurity. Arch. Ophthalmol. 2012, 130, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Vogler, S.; Pannicke, T.; Hollborn, M.; Grosche, A.; Busch, S.; Hoffmann, S.; Wiedemann, P.; Reichenbach, A.; Hammes, H.P.; Bringmann, A. Müller cell reactivity in response to photoreceptor degeneration in rats with defective polycystin-2. PLoS ONE 2013, 8, e61631. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Eyes | RA | 50%/12% O2 | 21%/12% O2 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Olive Oil | CoQ10 | n-3 PUFAs | Olive Oil | CoQ10 | n-3 PUFAs | Olive Oil | CoQ10 | n-3 PUFAs | |
| Left Eye | 15 (42%) | 11 (31%) | 32 (89%) *** | 11 (31%) | 19 (53%) | 34 (94%) ### | 19 (53%) | 28 (78%) ‡ | 29 (81%) § |
| Right Eye | 14 (39%) | 12 (33%) | 32 (89%) *** | 9 (25%) | 19 (53%) | 35 (97%) ### | 22 (61%) | 32 (89%) ‡ | 28 (78%) |
| Both Eyes | 11 (31%) | 10 (28%) | 31 (86%) *** | 7 (19%) | 18 (50%) | 33 (62%) ### | 18 (50%) | 27 (75%) ‡ | 25 (69%) |
| Ocular Compartment | Olive Oil | CoQ10 | n-3 PUFAs |
|---|---|---|---|
| P14-RA: | |||
| Vitreous (pg/mL): | 195.9 ± 61.0 | 127.5 ± 60.3 | 64.2 ± 17.6 |
| Retina (pg/mg protein): | 96.0 ± 12.5 | 181.6 ± 96.8 * | 65.8 ± 11.5 |
| Choroid: (pg/mg protein): | 419.0 ± 102.4 | 297.1 ± 133.0 | 913.7 ± 128.8 ** |
| P14-50/12% IH: | |||
| Vitreous (pg/mL): | 330.9 ± 59.3 § | 93.2 ± 50.9 ## | 42.7 ± 11.5 ## |
| Retina (pg/mg protein): | 240.4 ± 26.0 ** | 54.2 ± 13.2 ##§§ | 87.4 ± 16.7 ##§§ |
| Choroid: (pg/mg protein): | 1329.9 ± 198.9 ** | 681.3 ± 329.2 ##§ | 1044.4 ± 196.2 |
| P14-21/12% IH: | |||
| Vitreous (pg/mL): | 161.5 ± 95.6 | 50.0 ± 36.0 § | 37.9 ± 35.0 ‡§§ |
| Retina (pg/mg protein): | 237.5 ± 11.3 ** | 49.9 ± 9.1 ‡§§ | 51.8 ± 11.5 ‡ |
| Choroid: (pg/mg protein): | 1191.3 ± 292.7 ** | 525.2 ± 189.1 ‡ | 392.9 ± 124.7 ‡§ |
| P21-RA: | |||
| Vitreous (pg/mL): | 183.5 ± 20.3 | 216.8 ± 116.6 | 165.2 ± 27.2 |
| Retina (pg/mg protein): | 85.9 ± 5.1 | 104.1 ± 33.3 | 116.1 ± 2.9 * |
| Choroid: (pg/mg protein): | 1217.8 ± 162.4 | 632.3 ± 234.4 ** | 997.3 ± 131.3 ** |
| P21-50/12% IH: | |||
| Vitreous (pg/mL): | 260.9 ± 34.3 §§ | 225.4 ± 79.4 | 198.8 ± 60.0 |
| Retina (pg/mg protein): | 112.4 ± 55.4 | 89.5 ± 34.5 | 80.8 ± 9.8 §§ |
| Choroid: (pg/mg protein): | 1580.0 ± 223.4 ** | 514.8 ± 111.0 ## | 757.7 ± 55.4 ##§§ |
| P21-21/12% IH: | |||
| Vitreous (pg/mL): | 245.9 ± 29.4 §§ | 287.6 ± 132.8 §§ | 273.3 ± 55.1 |
| Retina (pg/mg protein): | 40.6 ± 23.8 | 94.1 ± 6.4 ‡ | 53.2 ± 9.6 §§ |
| Choroid: (pg/mg protein): | 595.0 ± 95.5 ** | 1215.4 ± 176.4 ‡§§ | 624.6 ± 130.1 §§ |
| Ocular Compartment | Olive Oil | CoQ10 | n-3 PUFAs |
|---|---|---|---|
| P14-RA: | |||
| Vitreous (ng/mL): | 795.1 ± 34.0 | 753.5 ± 130.1 | 793.1 ± 72.3 |
| Retina (ng/mg protein): | 8.4 ± 3.9 | 16.5 ± 11.8 | 23.6 ± 5.4 ** |
| Choroid: (ng/mg protein): | 30.5 ± 14.9 | 34.4 ± 33.8 | 173.3 ± 53.2 ** |
| P14-50/12% IH: | |||
| Vitreous (ng/mL): | 924.3 ± 127.1 | 872.2 ± 64.4 | 767.5 ± 62.7 # |
| Retina (ng/mg protein): | 27.2 ± 13.5 ** | 12.9 ± 10.8 | 21.1 ± 11.8 |
| Choroid: (ng/mg protein): | 12.1 ± 20.8 | 105.2 ± 23.5 #§§ | 204.4 ± 86.5 ## |
| P14-21/12% IH: | |||
| Vitreous (ng/mL): | 763.0 ± 276.8 | 803.6 ± 85.0 | 831.5 ± 54.1 |
| Retina (ng/mg protein): | 2.2 ± 2.9 | 25.2 ± 8.1 ‡ | 29.5 ± 4.7 ‡ |
| Choroid: (ng/mg protein): | 94.3 ± 52.2 ** | 125.4 ± 34.5 §§ | 49.4 ± 17.9 §§ |
| P21-RA: | |||
| Vitreous (ng/mL): | 708.7 ± 39.9 | 690.0 ± 108.0 | 509.7 ± 17.6 |
| Retina (ng/mg protein): | 25.8 ± 9.6 | 5.9 ± 4.2 ** | 40.0 ± 2.9 ** |
| Choroid: (ng/mg protein): | 83.4 ± 8.6 | 33.1 ± 9.6 ** | 80.6 ± 41.2 |
| P21-50/12% IH: | |||
| Vitreous (ng/mL): | 647.0 ± 67.1 | 756.2 ± 64.4 | 594.6 ± 45.8 |
| Retina (ng/mg protein): | 13.9 ± 8.3 * | 26.0 ± 10.8 #§§ | 13.1 ± 5.9 §§ |
| Choroid: (ng/mg protein): | 0 ± 0 ** | 49.2 ± 8.1 ##§ | 81.7 ± 23.0 ## |
| P21-21/12% IH: | |||
| Vitreous (ng/mL): | 603.1 ± 56.6 | 711.4 ± 89.9 | 888.1 ± 24.3 |
| Retina (ng/mg protein): | 1.3 ± 1.5 ** | 21.6 ± 6.1 ‡§§ | 14.0 ± 2.7 ‡§§ |
| Choroid: (ng/mg protein): | 46.7 ± 17.1 ** | 57.4 ± 10.3 §§ | 36.5 ± 10.8 § |
| Vascular Parameters | Olive Oil | CoQ10 | n-3 PUFAs |
|---|---|---|---|
| RA: | |||
| Vascular Density (%) | 34.9 ± 3.33 | 36.8 ± 7.3 | 33.7 ± 4.8 |
| Total Vascular Area | 28,068.8 ± 3366.1 | 27,084.5 ± 4843.6 | 22,814.7 ± 3429.8 ** |
| No. Branching Points | 568.9 ± 151.9 | 567.0 ± 245.4 | 424.9 ± 129.3 * |
| No. Segments | 1084.7 ± 205.3 | 1042.4 ± 392.9 | 813.4 ± 224.4 ** |
| Mean Segment Length | 25.7 ± 3.6 | 27.0 ± 4.4 | 28.9 ± 3.7 * |
| 50/12% IH: | |||
| Vascular Density (%) | 46.4 ± 16.7 ** | 32.9 ± 5.9 ## | 32.2 ± 2.6 ## |
| Total Vascular Area | 38,444.3 ± 10,394.0 ** | 25,052.0 ± 5403.6 ## | 21,725.2 ± 1943.4 ## |
| No. Branching Points | 839.9 ± 668.7 | 483.1 ± 230.7 # | 328.3 ± 51.4 ##§§ |
| No. Segments | 1458.1 ± 1035.2 | 905.6 ± 382.6 # | 655.5 ± 345.9 ## |
| Mean Segment Length | 23.6 ± 7.3 | 29.3 ± 4.8 ## | 33.3 ± 2.4 ##§§ |
| 21/12% IH: | |||
| Vascular Density (%) | 38.7 ± 3.6 ** | 31.9 ± 2.9 ‡§ | 33.3 ± 3.1 ‡ |
| Total Vascular Area | 29,057.7 ± 2680.2 | 24,396.9 ± 2427.0 ‡ | 26,001.0 ± 3100.1 ‡§§ |
| No. Branching Points | 652.4 ± 108.8 | 444.3 ± 94.1 ‡ | 500.5 ± 94.6 ‡§ |
| No. Segments | 1110.4 ± 182.3 | 848.1 ± 158.2 ‡ | 933.2 ± 161.2 ‡ |
| Mean Segment Length | 28.4 ± 2.5 | 29.5 ± 2.6 | 28.0 ± 2.1 |
| Parameters | Olive Oil | CoQ10 | n-3 PUFAs |
|---|---|---|---|
| RA: | |||
| Tortuosity Index | 0.96 ± 0.05 | 0.97 ± 0.05 | 0.98 ± 0.03 |
| Diameter of Arteries | 55.3 ± 9.3 | 50.6 ± 11.8 | 46.1 ± 7.3 ** |
| Diameter of Veins | 41.9 ± 10.3 | 38.4 ± 9.8 | 32.2 ± 5.9 ** |
| Number of Cells in NFL/GCL | 106.0 ± 19.6 | 115.0 ± 1.8 * | 125.6 ± 1.3 ** |
| Retinal Thickness (µm) | 342.2 ± 65.6 | 291.6 ± 76.9 * | 356.9 ± 71.5 |
| NFL/GCL Thickness (µm) | 48.0 ± 20.1 | 41.6 ± 21.1 | 48.3 ± 11.8 |
| IPL Thickness (µm) | 65.5 ± 21.1 | 61.0 ± 17.6 | 40.0 ± 16.2 ** |
| INL Thickness (µm) | 74.3 ± 13.7 | 58.9 ± 13.7 ** | 69.5 ± 12.7 |
| ONL Thickness (µm) | 109.6 ± 36.7 | 78.7 ± 12.7 ** | 103.3 ± 21.1 |
| 50/12% IH: | |||
| Tortuosity Index | 1.52 ± 0.24 ** | 1.1 ± 0.1 ##§§ | 1.08 ± 0.04 ##§§ |
| Diameter of Arteries | 60.9 ± 3.4 * | 45.1 ± 12.2 ## | 42.7 ± 10.3 ## |
| Diameter of Veins | 48.3 ± 8.3 * | 37.6 ± 5.9 ## | 34.9 ± 7.3 ## |
| Number of Cells in NFL/GCL | 231.8 ± 12.7 ** | 133.9 ± 1.8 ##§§ | 142.2 ± 2.0 ##§§ |
| Retinal Thickness (µm) | 433.9 ± 97.5 ** | 280.1 ± 45.1 ## | 408.1 ± 117.6 |
| NFL/GCL Thickness (µm) | 103.4 ± 44.6 ** | 44.5 ± 22.0 ## | 55.9 ± 28.4 ## |
| IPL Thickness (µm) | 86.4 ± 16.2 ** | 52.2 ± 8.8 ## | 71.6 ± 21.6 §§ |
| INL Thickness (µm) | 105.4 ± 44.6 ** | 60.6 ± 13.7 ## | 83.1 ± 25.5 # |
| ONL Thickness (µm) | 175.0 ± 91.1 ** | 82.3 ± 12.2 ## | 129.0 ± 73.0 # |
| 21/12% IH: | |||
| Tortuosity Index | 1.21 ± 0.15 ** | 1.09 ± 1.1 ‡§§ | 1.17 ± 0.15 §§ |
| Diameter of Arteries | 50.7 ± 8.3 | 44.6 ± 8.3 † | 47.9 ± 8.3 |
| Diameter of Veins | 35.8 ± 6.4 * | 31.7 ± 5.9 †§§ | 34.8 ± 6.4 |
| Number of Cells in NFL/GCL | 148.8 ± 2.9 ** | 139.9 ± 1.4 ‡§§ | 131.7 ± 1.7 ‡§§ |
| Retinal Thickness (µm) | 424.8 ± 82.3 ** | 411.8 ± 65.2 §§ | 392.9 ± 72.0 |
| NFL/GCL Thickness (µm) | 78.6 ± 23.5 ** | 65.7 ± 30.9 §§ | 59.3 ± 23.0 † |
| IPL Thickness (µm) | 63.7 ± 12.2 | 69.2 ± 18.1 | 66.0 ± 14.2 §§ |
| INL Thickness (µm) | 85.8 ± 12.2 | 78.1 ± 31.8 §§ | 78.3 ± 27.9 |
| ONL Thickness (µm) | 148.6 ± 44.6 | 110.9 ± 29.4 †§§ | 132.6 ± 79.4 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beharry, K.D.; Cai, C.L.; Siddiqui, F.; Chowdhury, S.; D’Agrosa, C.; Valencia, G.B.; Aranda, J.V. Comparative Effects of Coenzyme Q10 or n-3 Polyunsaturated Fatty Acid Supplementation on Retinal Angiogenesis in a Rat Model of Oxygen-Induced Retinopathy. Antioxidants 2018, 7, 160. https://doi.org/10.3390/antiox7110160
Beharry KD, Cai CL, Siddiqui F, Chowdhury S, D’Agrosa C, Valencia GB, Aranda JV. Comparative Effects of Coenzyme Q10 or n-3 Polyunsaturated Fatty Acid Supplementation on Retinal Angiogenesis in a Rat Model of Oxygen-Induced Retinopathy. Antioxidants. 2018; 7(11):160. https://doi.org/10.3390/antiox7110160
Chicago/Turabian StyleBeharry, Kay D., Charles L. Cai, Faisal Siddiqui, Sara Chowdhury, Christina D’Agrosa, Gloria B. Valencia, and Jacob V. Aranda. 2018. "Comparative Effects of Coenzyme Q10 or n-3 Polyunsaturated Fatty Acid Supplementation on Retinal Angiogenesis in a Rat Model of Oxygen-Induced Retinopathy" Antioxidants 7, no. 11: 160. https://doi.org/10.3390/antiox7110160