
Bioprospecting of Marine Macrophytes Using MS-Based Lipidomics as a New Approach


 , and
, and 
Abstract
:
1. Introduction
2. Marine Natural Products from Macrophytes
3. Bioactive Lipids from Marine Macrophytes
3.1. Fatty Acids
3.2. Glycolipids
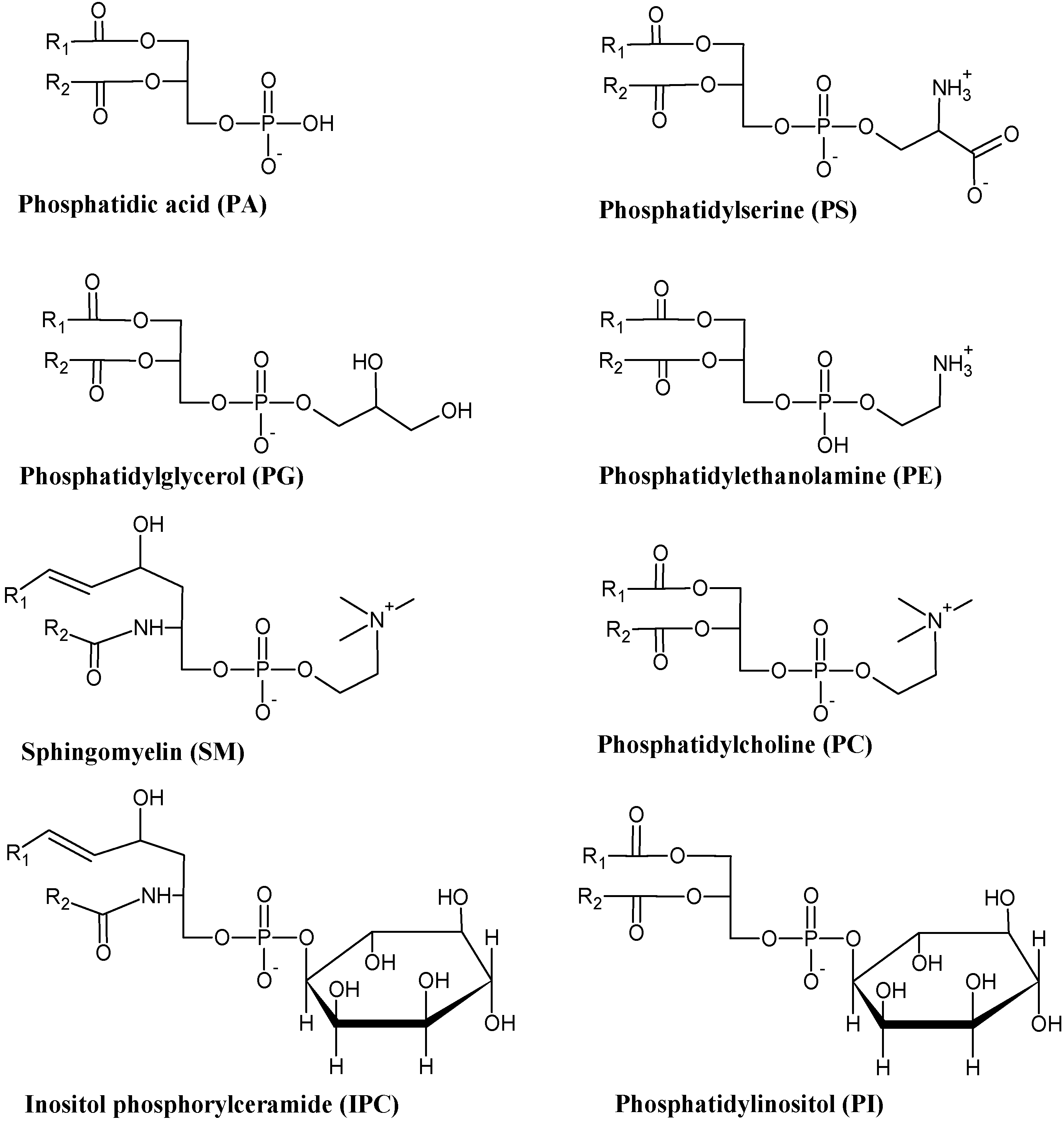
3.3. Phospholipids
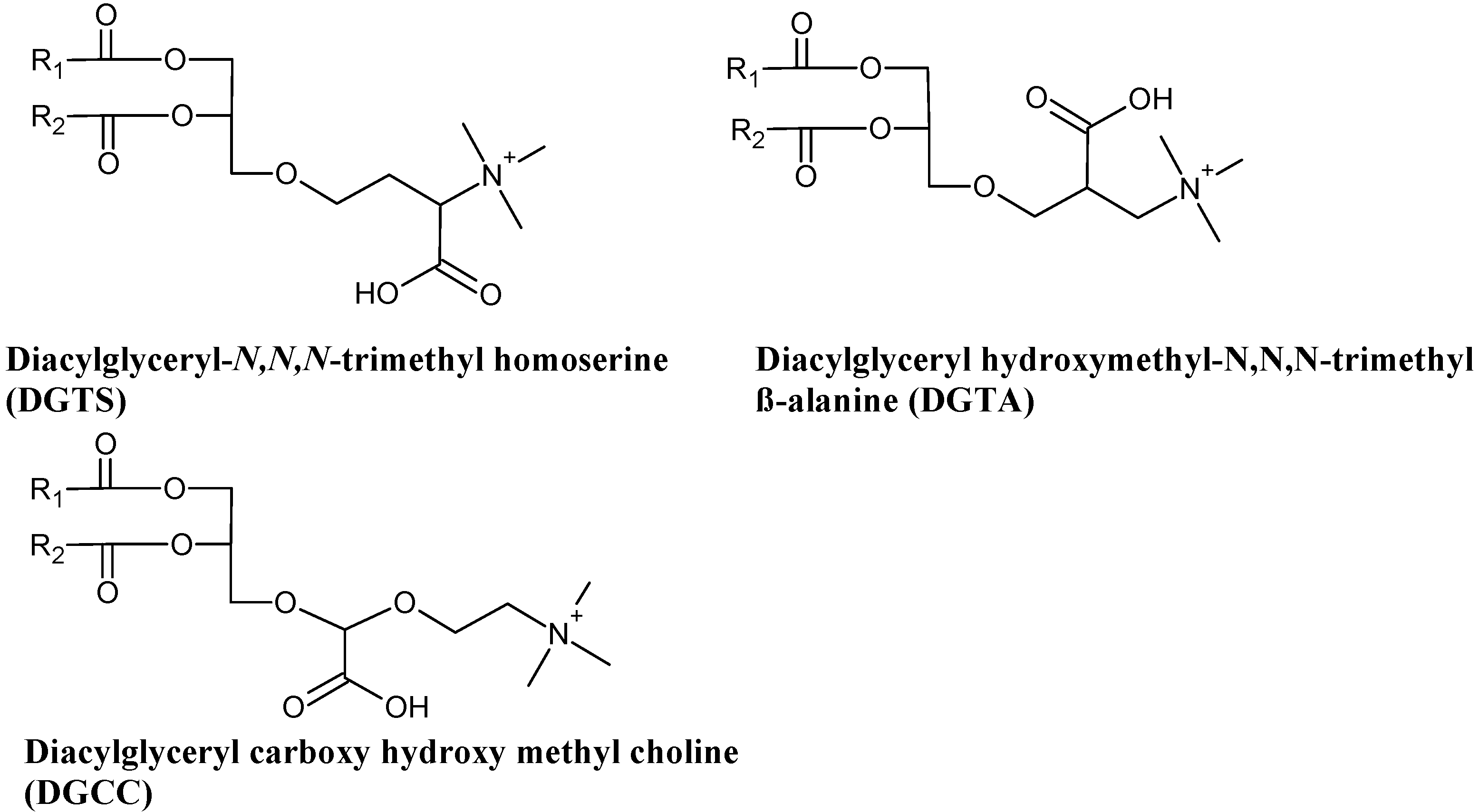
3.4. Betaine Lipids
3.5. Sterols
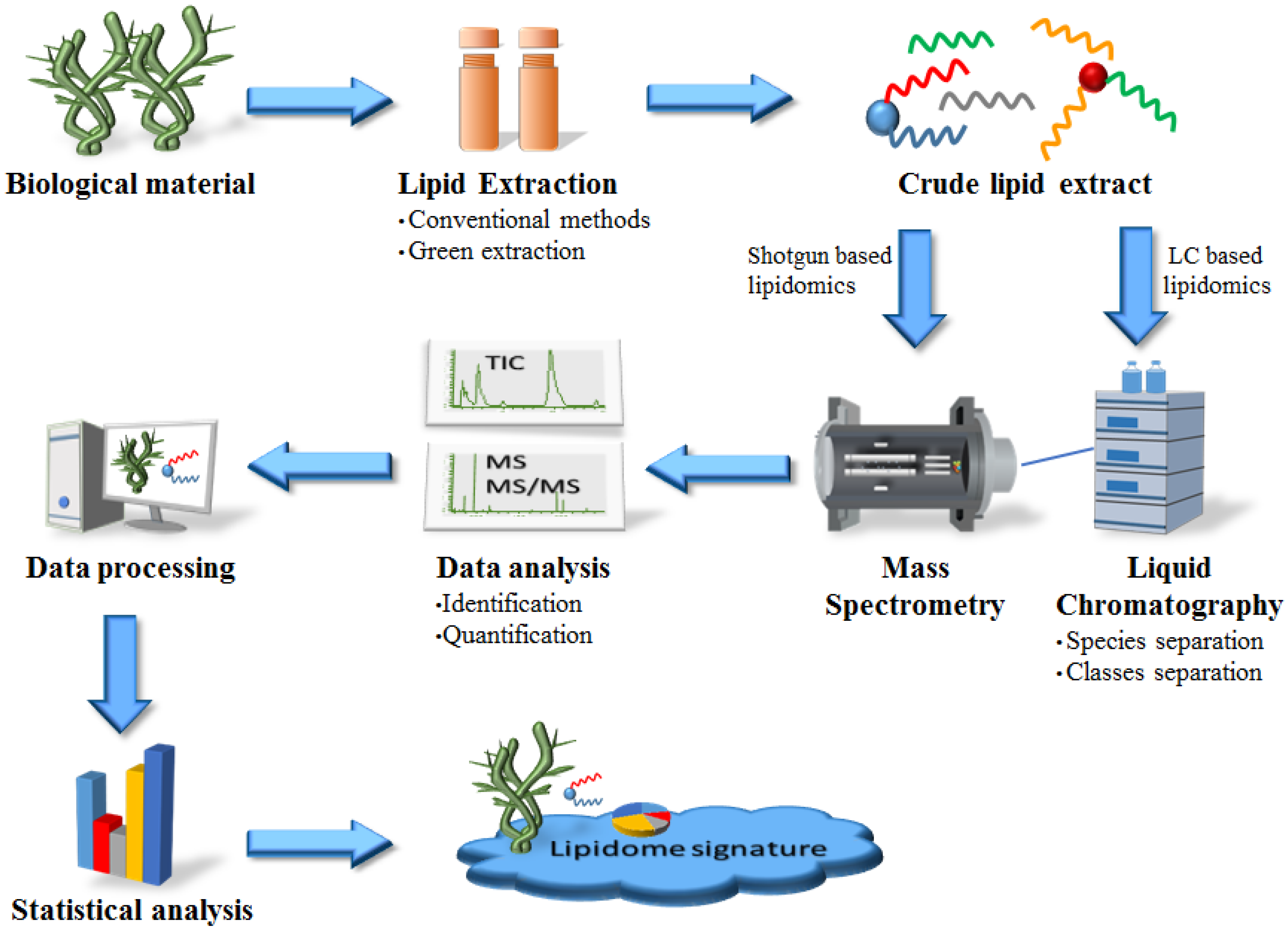
4. Strategies for Lipid Analysis from Marine Macrophytes: From Extraction to Structural Characterization
4.1. Methods of Lipid Extraction from Marine Macrophytes: Conventional vs. New Green Methods
4.1.1. Conventional Methods for Lipid Extraction
4.1.2. Green Extraction of Bioactive Compounds from Marine Macrophytes
4.2. Methods to Analyze Lipids Extracts from Marine Macrophytes
4.2.1. Thin Layer Chromatography (TLC)
4.2.2. Gas Chromatography (GC)
4.2.3. Liquid Chromatography (LC)
4.3. Mass Spectrometry-Based Lipidomics as a Valuable Tool to Find New Bioactive Lipids from Marine Macrophytes
4.4. Highlights of Mass Spectrometry-Based Lipidomics in Marine Macrophytes
5. Lipidomics Bioinformatics: Lipid Databases and Software
6. Future Perspectives
Acknowledgments
Conflicts of Interest
References
- Leal, M.C.; Puga, J.; Serôdio, J.; Gomes, N.C.M.; Calado, R. Trends in the discovery of new marine natural products from invertebrates over the last two decades—Where and what are we bioprospecting? PLoS ONE 2012, 7, e30580. [Google Scholar] [CrossRef] [PubMed]
- Ksouri, R.; Ksouri, W.M.; Jallali, I.; Debez, A.; Magné, C.; Hiroko, I.; Abdelly, C. Medicinal halophytes: Potent source of health promoting biomolecules with medical, nutraceutical and food applications. Crit. Rev. Biotechnol. 2012, 32, 289–326. [Google Scholar] [CrossRef] [PubMed]
- Leal, M.C.; Munro, M.H.G.; Blunt, J.W.; Puga, J.; Jesus, B.; Calado, R.; Rosa, R.; Madeira, C. Biogeography and biodiscovery hotspots of macroalgal marine natural products. Nat. Prod. Rep. 2013, 30, 1380–1390. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.J.; Gangadhar, K.N.; Vizetto-Duarte, C.; Wubshet, S.G.; Nyberg, N.T.; Barreira, L.; Varela, J.; Custódio, L. Maritime halophyte species from southern Portugal as sources of bioactive molecules. Mar. Drugs 2014, 12, 2228–2244. [Google Scholar] [CrossRef] [PubMed]
- Stengel, D.B.; Connan, S.; Popper, Z.A. Algal chemodiversity and bioactivity: Sources of natural variability and implications for commercial application. Biotechnol. Adv. 2011, 29, 483–501. [Google Scholar] [CrossRef] [PubMed]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Cortés-Sánchez, A.D.J.; Hernández-Sánchez, H.; Jaramillo-Flores, M.E. Biological activity of glycolipids produced by microorganisms: New trends and possible therapeutic alternatives. Microbiol. Res. 2013, 168, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Plouguerné, E.; da Gama, B.A. P.; Pereira, R.C.; Barreto-Bergter, E. Glycolipids from seaweeds and their potential biotechnological applications. Front. Cell. Infect. Microbiol. 2014, 4, 174. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.; Santarius, K.A. Changes in chloroplast membrane lipids during adaptation of barley to extreme salinity. Plant Physiol. 1978, 62, 326–329. [Google Scholar] [CrossRef] [PubMed]
- Costa, C.S.B.; Vicenti, J.R.M.; Moron-Villarreyes, J.A.; Caldas, S.; Cardoso, L.V.; Freitas, R.F.; D’Oca, M.G.M. Extraction and characterization of lipids from Sarcocornia ambigua meal: A halophyte biomass produced with shrimp farm effluent irrigation. An. Acad. Bras. Cienc. 2014, 86, 935–943. [Google Scholar] [CrossRef]
- D’Oca, M.G.M.; Moron-Villarreyes, J.A.; Lemoes, J.S.; Costa, C.S.B. Fatty acids composition in seeds of the South American glasswort Sarcocornia ambigua. An. Acad. Bras. Cienc. 2012, 84, 865–870. [Google Scholar] [CrossRef] [PubMed]
- Leal, M.C.; Calado, R. Marine natural products: Biodiscovery, biodiversity, and bioproduction. In Bioactive Natural Products; Wiley-VCH Verlag: Weinheim, Germany, 2014; pp. 473–490. [Google Scholar]
- Royal Society of Chemistry. MarinLit: A Database of the Marine Natural Products Literature. Available online: http://pubs.rsc.org/marinlit/ (accessed on 19 July 2015).
- Aronson, J. eHaloph Database. University of Sussex. Available online: http://www.sussex.ac.uk/affiliates/halophytes/index.php?content=about (accessed on 1 November 2015).
- Tsai, C.J.; Sun Pan, B. Identification of sulfoglycolipid bioactivities and characteristic fatty acids of marine macroalgae. J. Agric. Food Chem. 2012, 60, 8404–8410. [Google Scholar] [CrossRef] [PubMed]
- Tsydendambaev, V.D.; Ivanova, T.V.; Khalilova, L.A.; Kurkova, E.B.; Myasoedov, N.A.; Balnokin, Y.V. Fatty acid composition of lipids in vegetative organs of the halophyte Suaeda altissima under different levels of salinity. Russ. J. Plant Physiol. 2013, 60, 661–671. [Google Scholar] [CrossRef]
- Wu, J.; Seliskar, D.M.; Gallagher, J.L. The response of plasma membrane lipid composition in callus of the halophyte Spartina patens (Poaceae) to salinity stress. Am. J. Bot. 2005, 92, 852–858. [Google Scholar] [CrossRef] [PubMed]
- Eckardt, N.A. Oxylipin signaling in plant stress responses. Plant Cell 2008, 20, 495–497. [Google Scholar] [CrossRef] [PubMed]
- Floreto, E.A.T.; Hirata, H.; Yamasaki, S.; Castro, S.C. Effect of salinity on the growth and fatty acid composition of Ulva pertusa Kjellman (Chlorophyta). Bot. Mar. 1994, 37, 151–156. [Google Scholar] [CrossRef]
- Kumari, P.; Kumar, M.; Reddy, C.R.K.; Jha, B. 3—Algal lipids, fatty acids and sterols A2—Domínguez, Herminia. In Functional Ingredients from Algae for Foods and Nutraceuticals; Woodhead Publishing: Cambridge, UK, 2013; pp. 87–134. [Google Scholar]
- Sanina, N.M.; Goncharova, S.N.; Kostetsky, E.Y. Fatty acid composition of individual polar lipid classes from marine macrophytes. Phytochemistry 2004, 65, 721–730. [Google Scholar] [CrossRef] [PubMed]
- Goncharova, S.N.; Kostetsky, E.Y.; Sanina, N.M. The effect of seasonal shifts in temperature on the lipid composition of marine macrophytes. Russ. J. Plant Physiol. 2004, 51, 169–175. [Google Scholar] [CrossRef]
- Kim, M.K.; Dubacq, J.P.; Thomas, J.C.; Giraud, G. Seasonal variations of triacylglycerols and fatty acids in Fucus serratus. Phytochemistry 1996, 43, 49–55. [Google Scholar] [CrossRef]
- Kostetsky, E.Y.; Goncharova, S.N.; Sanina, N.M.; Shnyrov, V.L. Season influence on lipid composition of marine macrophytes. Bot. Mar. 2004, 47, 134–139. [Google Scholar] [CrossRef]
- Nelson, M.M.; Phleger, C.F.; Nichols, P.D. Seasonal lipid composition in macroalgae of the northeastern Pacific Ocean. Bot. Mar. 2002, 45, 58–65. [Google Scholar] [CrossRef]
- Sanina, N.M.; Goncharova, S.N.; Kostetsky, E.Y. Seasonal changes of fatty acid composition and thermotropic behavior of polar lipids from marine macrophytes. Phytochemistry 2008, 69, 1517–1527. [Google Scholar] [CrossRef] [PubMed]
- Leal, M.C.; Calado, R.; Sheridan, C.; Alimonti, A.; Osinga, R. Coral aquaculture to support drug discovery. Trends Biotechnol. 2013, 31, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Sui, N.; Han, G. Salt-induced photoinhibition of PSII is alleviated in halophyte Thellungiella halophila by increases of unsaturated fatty acids in membrane lipids. Acta Physiol. Plant. 2014, 36, 983–992. [Google Scholar] [CrossRef]
- Sui, N.; Han, G. Increases of unsaturated fatty acids in membrane lipids protects photosystem II from photoinhibition under salinity in different halophytes. J. Agric. Sci. 2014, 6. [Google Scholar] [CrossRef]
- Sui, N.; Li, M.; Li, K.; Song, J.; Wang, B.S. Increase in unsaturated fatty acids in membrane lipids of Suaeda salsa L. enhances protection of photosystem II under high salinity. Photosynthetica 2010, 48, 623–629. [Google Scholar] [CrossRef]
- Floreto, E.A.T.; Teshima, S. The fatty acid composition of seaweeds exposed to different levels of light intensity and salinity. Bot. Mar. 1998, 41, 467–481. [Google Scholar] [CrossRef]
- Gerasimenko, N.I.; Skriptsova, A.V.; Busarova, N.G.; Moiseenko, O.P. Effects of the season and growth stage on the contents of lipids and photosynthetic pigments in brown alga Undaria pinnatifida. Russ. J. Plant Physiol. 2011, 58, 885–891. [Google Scholar] [CrossRef]
- Siriwardhana, N.; Kalupahana, N.S.; Moustaid-Moussa, N. Chapter 13—Health Benefits of n-3 Polyunsaturated Fatty Acids: Eicosapentaenoic Acid and Docosahexaenoic Acid. In Advances in Food and Nutrition Research; Se-Kwon, K., Ed.; Academic Press: Waltham, MA, USA, 2012; Volume 65, pp. 211–222. [Google Scholar]
- Rozentsvet, O.A.; Nesterov, V.N.; Bogdanova, E.S. Membrane-forming lipids of wild halophytes growing under the conditions of Prieltonie of South Russia. Phytochemistry 2014, 105, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Khotimchenko, S.V. Distribution of glyceroglycolipids in marine algae and grasses. Chem. Nat. Compd. 2002, 38, 223–229. [Google Scholar] [CrossRef]
- Gigon, A.; Matos, A.R.; Laffray, D.; Zuily-Fodil, Y.; Pham-Thi, A.T. Effect of drought stress on lipid metabolism in the leaves of Arabidopsis thaliana (Ecotype Columbia). Ann. Bot. 2004, 94, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Melo, T.; Alves, E.; Azevedo, V.; Martins, A.S.; Neves, B.; Domingues, P.; Calado, R.; Abreu, M.H.; Domingues, M.R. Lipidomics as a new approach for the bioprospecting of marine macroalgae—Unraveling the polar lipid and fatty acid composition of Chondrus crispus. Algal Res. 2015, 8, 181–191. [Google Scholar] [CrossRef]
- Khotimchenko, S.V. The fatty acid composition of glycolipids of marine macrophytes. Russ. J. Mar. Biol. 2003, 29, 126–128. [Google Scholar] [CrossRef]
- Plouguerné, E.; de Souza, L.M.; Sassaki, G.L.; Cavalcanti, J.F.; Romanos, M.T.V.; da Gama, B.A.P.; Pereira, R.C.; Barreto-Bergter, E. Antiviral sulfoquinovosyldiacylglycerols (SQDGs) from the Brazilian brown seaweed Sargassum vulgare. Mar. Drugs 2013, 11, 4628–4640. [Google Scholar] [CrossRef] [PubMed]
- Boudière, L.; Michaud, M.; Petroutsos, D.; Rébeillé, F.; Falconet, D.; Bastien, O.; Roy, S.; Finazzi, G.; Rolland, N.; Jouhet, J.; Block, M.A.; Maréchal, E. Glycerolipids in photosynthesis: Composition, synthesis and trafficking. Biochim. Biophys. Acta Bioenerg. 2014, 1837, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Bergé, J.P.; Debiton, E.; Dumay, J.; Durand, P.; Barthomeuf, C. In vitro anti-inflammatory and anti-proliferative activity of sulfolipids from the red alga Porphyridium cruentum. J. Agric. Food Chem. 2002, 50, 6227–6232. [Google Scholar] [CrossRef] [PubMed]
- El Baz, F.K.; El Baroty, G.S.; Abd El Baky, H.H.; Abd El-Salam, O.I.; Ibrahim, E.A. Structural characterization and biological activity of sulfolipids from selected marine algae. Grasas Aceites 2013, 64, 561–571. [Google Scholar]
- Al-Fadhli, A.; Wahidulla, S.; D’Souza, L. Glycolipids from the red alga Chondria armata (Kütz.) Okamura. Glycobiology 2006, 16, 902–915. [Google Scholar] [CrossRef] [PubMed]
- De Souza, L.M.; Sassaki, G.L.; Romanos, M.T.V.; Barreto-Bergter, E. Structural characterization and anti-HSV-1 and HSV-2 activity of glycolipids from the marine algae Osmundaria obtusiloba isolated from Southeastern Brazilian coast. Mar. Drugs 2012, 10, 918–931. [Google Scholar] [CrossRef] [PubMed]
- Banskota, A.; Stefanova, R.; Sperker, S.; Lall, S.; Craigie, J.; Hafting, J. Lipids isolated from the cultivated red alga Chondrus crispus inhibit nitric oxide production. J. Appl. Phycol. 2014, 26, 1565–1571. [Google Scholar] [CrossRef]
- Banskota, A.H.; Stefanova, R.; Sperker, S.; Lall, S.P.; Craigie, J.S.; Hafting, J.T.; Critchley, A.T. Polar lipids from the marine macroalga Palmaria palmata inhibit lipopolysaccharide-induced nitric oxide production in RAW264.7 macrophage cells. Phytochemistry 2014, 101, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Lopes, G.; Daletos, G.; Proksch, P.; Andrade, P.B.; Valentão, P. Anti-inflammatory potential of monogalactosyl diacylglycerols and a monoacylglycerol from the edible brown seaweed Fucus spiralis linnaeus. Mar. Drugs 2014, 12, 1406–1418. [Google Scholar] [CrossRef] [PubMed]
- Ksouri, R.; Falleh, H.; Megdiche, W.; Trabelsi, N.; Mhamdi, B.; Chaieb, K.; Bakrouf, A.; Magné, C.; Abdelly, C. Antioxidant and antimicrobial activities of the edible medicinal halophyte Tamarix gallica L. and related polyphenolic constituents. Food Chem. Toxicol. 2009, 47, 2083–2091. [Google Scholar] [CrossRef] [PubMed]
- Ksouri, W.M.; Medini, F.; Mkadmini, K.; Legault, J.; Magné, C.; Abdelly, C.; Ksouri, R. LC-ESI-TOF-MS identification of bioactive secondary metabolites involved in the antioxidant, anti-inflammatory and anticancer activities of the edible halophyte Zygophyllum album Desf. Food Chem. 2013, 139, 1073–1080. [Google Scholar] [CrossRef] [PubMed]
- Kendel, M.; Wielgosz-Collin, G.; Bertrand, S.; Roussakis, C.; Bourgougnon, N.; Bedoux, G. Lipid composition, fatty acids and sterols in the seaweeds Ulva armoricana, and Solieria chordalis from Brittany (France): An analysis from nutritional, chemotaxonomic, and antiproliferative activity perspectives. Mar. Drugs 2015, 13, 5606–5628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossain, Z.; Kurihara, H.; Hosokawa, M.; Takahashi, K. Growth inhibition and induction of differentiation and apoptosis mediated by sodium butyrate in Caco-2 cells with algal glycolipids. In Vitro Cell. Dev. Biol. Anim. 2005, 41, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Kim, E.H.; Lee, C.; Kim, M.H.; Rho, J.R. Two new monogalactosyl diacylglycerols from brown alga Sargassum thunbergii. Lipids 2007, 42, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Arunkumar, K.; Selvapalam, N.; Rengasamy, R. The antibacterial compound sulphoglycerolipid 1-0 palmitoyl-3-0(6’-sulpho-α-quinovopyranosyl)-glycerol from Sargassum wightii Greville (Phaeophyceae). Bot. Mar. 2005, 48, 441–445. [Google Scholar] [CrossRef]
- Kannan, R.R.R.; Arumugam, R.; Anantharaman, P. Antibacterial potential of three seagrasses against human pathogens. Asian Pac. J. Trop. Med. 2010, 3, 890–893. [Google Scholar] [CrossRef]
- Kumar, C.S.; Sarada, D.V.L.; Thomas, P.G.; Rengasamy, R. Antibacterial activity of three South Indian seagrasses, Cymodocea serrulata, Halophila ovalis and Zostera capensis. World J. Microbiol. Biotechnol. 2008, 24, 1989–1992. [Google Scholar] [CrossRef]
- Hua, K.F.; Hsu, H.Y.; Su, Y.C.; Lin, I.F.; Yang, S.S.; Chen, Y.M.; Chao, L.K. Study on the antiinflammatory activity of methanol extract from seagrass Zostera japonica. J. Agric. Food Chem. 2006, 54, 306–311. [Google Scholar] [CrossRef] [PubMed]
- Kumari, P.; Kumar, M.; Reddy, C.R. K.; Jha, B. Nitrate and phosphate regimes induced lipidomic and biochemical changes in the intertidal macroalga Ulva lactuca (Ulvophyceae, Chlorophyta). Plant Cell Physiol. 2014, 55, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Ohlrogge, J.; Browse, J. Lipid biosynthesis. Plant Cell 1995, 7, 957–970. [Google Scholar] [CrossRef] [PubMed]
- Janero, D.R.; Barrnett, R. Cellular and thylakoid-membrane phospholipids of Chlamydomonas reinhardtii 137+. J. Lipid Res. 1981, 22, 1126–1130. [Google Scholar] [PubMed]
- Dembitsky, V.M.; Rozentsvet, O.A. Phospholipid composition of some marine red algae. Phytochemistry 1990, 29, 3149–3152. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Rozentsvet, O.A.; Pechenkina, E.E. Glycolipids, phospholipids and fatty acids of brown algae species. Phytochemistry 1990, 29, 3417–3421. [Google Scholar] [CrossRef]
- Khotimchenko, S.V.; Kulikova, I.V. Lipids of two species of brown algae of the genus Laminaria. Chem. Nat. Compd. 1999, 35, 17–20. [Google Scholar] [CrossRef]
- Vaskovsky, V.E.; Khotimchenko, S.V.; Xia, B.; Hefang, L. Polar lipids and fatty acids of some marine macrophytes from the Yellow Sea. Phytochemistry 1996, 42, 1347–1356. [Google Scholar] [CrossRef]
- Khotimchenko, S.V.; Kulikova, I.V.; Vas’kovskii, V.E. Distribution of ceramidephosphoinositol in red seaweeds. Russ. J. Mar. Biol. 2000, 26, 286–288. [Google Scholar] [CrossRef]
- Khotimchenko, S.V.; Vas’kovsky, V.E. An inositol-containing sphingolipid from the red alga Gracilaria verrucosa. Bioorg. Khim. 2004, 30, 190–194. [Google Scholar] [CrossRef] [PubMed]
- Küllenberg, D.; Taylor, L.A.; Schneider, M.; Massing, U. Health effects of dietary phospholipids. Lipids Health Dis. 2012, 11, 3. [Google Scholar] [CrossRef] [PubMed]
- Burri, L.; Hoem, N.; Banni, S.; Berge, K. Marine Omega-3 phospholipids: Metabolism and biological activities. Int. J. Mol. Sci. 2012, 13, 15401–15419. [Google Scholar] [CrossRef] [PubMed]
- da Costa, E.; Melo, T.; Moreira, A.S.P.; Alves, E.; Domingues, P.; Calado, R.; Abreu, M.H.; Domingues, M.R. Decoding bioactive polar lipid profile of the macroalgae Codium tomentosum from a sustainable IMTA system using a lipidomic approach. Algal Res. 2015, 12, 388–397. [Google Scholar] [CrossRef]
- Dembitsky, V.M. Betaine ether-linked glycerolipids: Chemistry and biology. Prog. Lipid Res. 1996, 35, 1–51. [Google Scholar] [CrossRef]
- Hofmann, M.; Eichenberger, W. Lipid and fatty acid composition of the marine brown alga Dictyopteris membranacea. Plant Cell Physiol. 1997, 38, 1046–1052. [Google Scholar] [CrossRef]
- Makewicz, A.; Gribi, C.; Eichenberger, W. Lipids of Ectocarpus fasciculatus (Phaeophyceae). Incorporation of [l-14C]oleate and the role of TAG and MGDG in lipid metabolism. Plant Cell Physiol. 1997, 38, 952–960. [Google Scholar] [CrossRef]
- Kato, M.; Sakai, M.; Adachi, K.; Ikemoto, H.; Sano, H. Distribution of betaine lipids in marine algae. Phytochemistry 1996, 42, 1341–1345. [Google Scholar] [CrossRef]
- Terasaki, M.; Hirose, A.; Narayan, B.; Baba, Y.; Kawagoe, C.; Yasui, H.; Saga, N.; Hosokawa, M.; Miyashita, K. Evaluation of recoverable functional lipid components of several brown seaweeds (Phaeophyta) from Japan with special reference to fucoxanthin and fucosterol contents. J. Phycol. 2009, 45, 974–980. [Google Scholar] [CrossRef]
- Patterson, G. The distribution of sterols in algae. Lipids 1971, 6, 120–127. [Google Scholar] [CrossRef]
- Ivanova, A.; Khozin-Goldberg, I.; Kamenarska, Z.; Nechev, J.; Cohen, Z.; Popov, S.; Stefanov, K. Lipophylic compounds from Euphorbia peplis L.—A halophytic plant from the Bulgarian Black Sea Coast. Z. Naturforsch. C 2003, 58, 783–788. [Google Scholar] [PubMed]
- Mansour, M.M.F.; Salama, K.H.A.; Al-Mutawa, M.M.; Abou Hadid, A.F. Effect of NaCl and polyamines on plasma membrane lipids of wheat roots. Biol. Plant. 2002, 45, 235–239. [Google Scholar] [CrossRef]
- Normén, L.; Dutta, P.; Lia, A.; Andersson, H. Soy sterol esters and β-sitostanol ester as inhibitors of cholesterol absorption in human small bowel. Am. J. Clin. Nutr. 2000, 71, 908–913. [Google Scholar] [PubMed]
- Grille, S.; Zaslawski, A.; Thiele, S.; Plat, J.; Warnecke, D. The functions of steryl glycosides come to those who wait: Recent advances in plants, fungi, bacteria and animals. Prog. Lipid Res. 2010, 49, 262–288. [Google Scholar] [CrossRef] [PubMed]
- El Shoubaky, G.A.; Salem, E.A. Terpenes and sterols composition of marine brown algae Padina pavonica (Dictyotales) and Hormophysa triquetra (Fucales). Int. J. Pharm. Phytochem. Res. 2014, 6, 894–900. [Google Scholar]
- Hummel, J.; Segu, S.; Li, Y.; Irgang, S.; Jueppner, J.; Giavalisco, P. Ultra performance liquid chromatography and high resolution mass spectrometry for the analysis of plant lipids. Front. Plant Sci. 2011, 2. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, Y.; Kamide, Y.; Hirai, M.Y.; Saito, K. Plant lipidomics based on hydrophilic interaction chromatography coupled to ion trap time-of-flight mass spectrometry. Metabolomics 2013, 9 (Suppl. S1), 121–131. [Google Scholar] [CrossRef] [PubMed]
- Herrero, M.; Simõ, C.; García-Cañas, V.; Ibáñez, E.; Cifuentes, A. Foodomics: MS-based strategies in modern food science and nutrition. Mass Spectrom. Rev. 2012, 31, 49–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smilowitz, J.T.; Zivkovic, A.M.; Wan, Y.J.Y.; Watkins, S.M.; Nording, M.L.; Hammock, B.D.; German, J.B. Nutritional lipidomics: Molecular metabolism, analytics, and diagnostics. Mol. Nutr. Food Res. 2013, 57, 1319–1335. [Google Scholar] [CrossRef] [PubMed]
- Murphy, S.A.; Nicolaou, A. Lipidomics applications in health, disease and nutrition research. Mol. Nutr. Food Res. 2013, 57, 1336–1346. [Google Scholar] [CrossRef] [PubMed]
- Wolf, C.; Quinn, P.J. Lipidomics: Practical aspects and applications. Prog. Lipid Res. 2008, 47, 15–36. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Cequier-Sanchez, E.; Rodriguez, C.; Ravelo, A.G.; Zarate, R. Dichloromethane as a solvent for lipid extraction and assessment of lipid classes and fatty acids from samples of different natures. J. Agric. Food Chem. 2008, 56, 4297–4303. [Google Scholar] [CrossRef] [PubMed]
- Matyash, V.; Liebisch, G.; Kurzchalia, T.V.; Shevchenko, A.; Schwudke, D. Lipid extraction by methyl-tert-butyl ether for high-throughput lipidomics. J. Lipid Res. 2008, 49, 1137–1146. [Google Scholar] [CrossRef] [PubMed]
- Kumari, P.; Reddy, C.R.K.; Jha, B. Comparative evaluation and selection of a method for lipid and fatty acid extraction from macroalgae. Anal. Biochem. 2011, 415, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Ibañez, E.; Herrero, M.; Mendiola, J.A.; Castro-Puyana, M. Extraction and characterization of bioactive compounds with health benefits from marine resources: Macro and micro algae, cyanobacteria, and invertebrates. In Marine Bioactive Compounds: Sources, Characterization and Applications; Springer US: New York, NY, USA, 2014; pp. 55–98. [Google Scholar]
- Grosso, C.; Valentão, P.; Ferreres, F.; Andrade, P. Alternative and efficient extraction methods for marine-derived compounds. Mar. Drugs 2015, 13, 3182–3230. [Google Scholar] [CrossRef] [PubMed]
- Duarte, K.; Justino, C.I.L.; Gomes, A.M.; Rocha-Santos, T.; Duarte, A.C. Chapter 4—Green analytical methodologies for preparation of extracts and analysis of bioactive compounds. In Comprehensive Analytical Chemistry; Teresa, R.S., Armando, C.D., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; Volume 65, pp. 59–78. [Google Scholar]
- Michalak, I.; Chojnacka, K. Algal extracts: Technology and advances. Eng. Life Sci. 2014, 14, 581–591. [Google Scholar] [CrossRef]
- Pettitt, T.R.; Jones, A.L.; Harwood, J.L. Lipid metabolism in the red marine algae Chondrus crispus and Polysiphonia lanosa as modified by temperature. Phytochemistry 1989, 28, 2053–2058. [Google Scholar] [CrossRef]
- Graeve, M.; Janssen, D. Improved separation and quantification of neutral and polar lipid classes by HPLC–ELSD using a monolithic silica phase: Application to exceptional marine lipids. J. Chromatogr. B 2009, 877, 1815–1819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordbäck, J.; Lundberg, E.; Christie, W.W. Separation of lipid classes from marine particulate material by HPLC on a polyvinyl alcohol-bonded stationary phase using dual-channel evaporative light-scattering detection. Mar. Chem. 1998, 60, 165–175. [Google Scholar] [CrossRef]
- Logvinov, S.; Gerasimenko, N.; Esipov, A.; Denisenko, V.A. Examination of the structures of several glycerolipids from marine macroalgae by NMR and GC-MS. J. Phycol. 2015, 51, 1066–1074. [Google Scholar] [CrossRef]
- Gupta, V.; Thakur, R.S.; Reddy, C.R.K.; Jha, B. Central metabolic processes of marine macrophytic algae revealed from NMR based metabolome analysis. RSC Adv. 2013, 3, 7037–7047. [Google Scholar] [CrossRef]
- Sommer, U.; Herscovitz, H.; Welty, F.K.; Costello, C.E. LC-MS-based method for the qualitative and quantitative analysis of complex lipid mixtures. J. Lipid Res. 2006, 47, 804–814. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, B.; Süß, R.; Teuber, K.; Eibisch, M.; Schiller, J. Lipid analysis by thin-layer chromatography—A review of the current state. J. Chromatogr. A 2011, 1218, 2754–2774. [Google Scholar] [CrossRef] [PubMed]
- Olsen, R.E.; Henderson, R.J. The rapid analysis of neutral and polar marine lipids using double-development HPTLC and scanning densitometry. J. Exp. Mar. Biol. Ecol. 1989, 129, 189–197. [Google Scholar] [CrossRef]
- Srivastava, M. High-Performance Thin-Layer Chromatography (HPTLC); Springer: Berlin, Germany, 2011; pp. 1–397. [Google Scholar]
- Bartlett, E.M.; Lewis, D.H. Spectrophotometric determination of phosphate esters in the presence and absence of orthophosphate. Anal. Biochem. 1970, 36, 159–167. [Google Scholar] [CrossRef]
- Momose, T.; Ohkura, Y. Organic analysis—XX: Microestimation of blood sugar with 5-Hydroxy-1-tetralone. Talanta 1959, 3, 151–154. [Google Scholar] [CrossRef]
- Cheng, S.C.; Huang, M.Z.; Shiea, J. Thin layer chromatography/mass spectrometry. J. Chromatogr. A 2011, 1218, 2700–2711. [Google Scholar] [CrossRef] [PubMed]
- Ragonese, C.; Tedone, L.; Beccaria, M.; Torre, G.; Cichello, F.; Cacciola, F.; Dugo, P.; Mondello, L. Characterisation of lipid fraction of marine macroalgae by means of chromatography techniques coupled to mass spectrometry. Food Chem. 2014, 145, 932–940. [Google Scholar] [CrossRef] [PubMed]
- Ramani, B.; Zorn, H.; Papenbrock, J. Quantification and fatty acid profiles of sulfolipids in two halophytes and a glycophyte grown under different salt concentrations. Z. Naturforsch. C 2004, 59, 835–842. [Google Scholar] [CrossRef] [PubMed]
- Seppänen-Laakso, T.; Laakso, I.; Hiltunen, R. Analysis of fatty acids by gas chromatography, and its relevance to research on health and nutrition. Anal. Chim. Acta 2002, 465, 39–62. [Google Scholar] [CrossRef]
- Eder, K. Gas chromatographic analysis of fatty acid methyl esters. J. Chromatogr. B Biomed. Sci. Appl. 1995, 671, 113. [Google Scholar] [CrossRef]
- Bannon, C.D.; Craske, J.D.; Hilliker, A.E. Analysis of fatty acid methyl esters with high accuracy and reliability. IV. Fats with fatty acids containing four or more carbon atoms. J. Am. Oil Chem. Soc. 1985, 62, 1501–1507. [Google Scholar] [CrossRef]
- Gutnikov, G. Fatty acid profiles of lipid samples. J. Chromatogr. B Biomed. Sci. Appl. 1995, 671, 71–89. [Google Scholar] [CrossRef]
- Nouairi, I.; Ghnaya, T.; Ben Youssef, N.; Zarrouk, M.; Habib Ghorbel, M. Changes in content and fatty acid profiles of total lipids of two halophytes: Sesuvium portulacastrum and Mesembryanthemum crystallinum under cadmium stress. J. Plant Physiol. 2006, 163, 1198–1202. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, T.V.; Myasoedov, N.A.; Pchelkin, V.P.; Tsydendambaev, V.D.; Vereshchagin, A.G. Increased content of very-long-chain fatty acids in the lipids of halophyte vegetative organs. Russ. J. Plant Physiol. 2009, 56, 787–794. [Google Scholar] [CrossRef]
- Van Ginneken, V.J.T.; Helsper, J.P.F.G.; de Visser, W.; van Keulen, H.; Brandenburg, W.A. Polyunsaturated fatty acids in various macroalgal species from north Atlantic and tropical seas. Lipids Health Dis. 2011, 10, 104. [Google Scholar] [CrossRef] [PubMed]
- Cajka, T.; Fiehn, O. Comprehensive analysis of lipids in biological systems by liquid chromatography-mass spectrometry. Trends Analyt. Chem. 2014, 61, 192–206. [Google Scholar] [CrossRef] [PubMed]
- Blaas, N.; Humpf, H.U. Structural profiling and quantitation of glycosyl inositol phosphoceramides in plants with Fourier transform mass spectrometry. J. Agric. Food Chem. 2013, 61, 4257–4269. [Google Scholar] [CrossRef] [PubMed]
- Guella, G.; Frassanito, R.; Mancini, I. A new solution for an old problem: The regiochemical distribution of the acyl chains in galactolipids can be established by electrospray ionization tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2003, 17, 1982–1994. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Verma, R.K.; Sahoo, D.; Srivastava, S.K. Reverse-phase HPLC method for the quantification of two antihyperglycemic glycolipids in Oplismenus burmannii. Biomed. Chromatogr. 2015, 29, 1675–1681. [Google Scholar] [CrossRef] [PubMed]
- Zábranská, M.; Vrkoslav, V.; Sobotníková, J.; Cvačka, J. Analysis of plant galactolipids by reversed-phase high-performance liquid chromatography/mass spectrometry with accurate mass measurement. Chem. Phys. Lipids 2012, 165, 601–607. [Google Scholar] [CrossRef] [PubMed]
- Sajiki, J.; Kakimi, H. Identification of eicosanoids in the red algae, Gracilaria asiatica, using high-performance liquid chromatography and electrospray ionization mass spectrometry. J. Chromatogr. A 1998, 795, 227–237. [Google Scholar] [CrossRef]
- Yang, J.; Schmelzer, K.; Georgi, K.; Hammock, B.D. Quantitative profiling method for oxylipin metabolome by liquid chromatography electrospray ionization tandem mass spectrometry. Anal. Chem. 2009, 81, 8085–8093. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Gross, R.W. Global analyses of cellular lipidomes directly from crude extracts of biological samples by ESI mass spectrometry: A bridge to lipidomics. J. Lipid Res. 2003, 44, 1071–1079. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhou, Z.; Nie, H.; Bai, Y.; Liu, H. Recent advances of chromatography and mass spectrometry in lipidomics. Anal. Bioanal. Chem. 2011, 399, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Domingues, M.R.M.; Reis, A.; Domingues, P. Mass spectrometry analysis of oxidized phospholipids. Chem. Phys. Lipids 2008, 156, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Gross, R.W.; Han, X. Shotgun lipidomics of neutral lipids as an enabling technology for elucidation of lipid-related diseases. Am. J. Physiol. Endocrinol. Metab. 2009, 297, E297–E303. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, P.T.; Milne, S.B.; Byrne, M.O.; Xiang, Y.; Brown, H.A. Glycerophospholipid identification and quantitation by electrospray ionization mass spectrometry. Methods Enzymol. 2007, 432, 21–57. [Google Scholar] [PubMed]
- Pulfer, M.; Murphy, R.C. Electrospray mass spectrometry of phospholipids. Mass Spectrom. Rev. 2003, 22, 332–364. [Google Scholar] [CrossRef] [PubMed]
- Harrison, K.A.; Murphy, R.C. Negative electrospray ionization of glycerophosphocholine lipids: Formation of [M – 15]− ions occurs via collisional decomposition of adduct anions. J. Mass Spectrom. 1995, 30, 1772–1773. [Google Scholar] [CrossRef]
- Zhang, X.; Reid, G.E. Multistage tandem mass spectrometry of anionic phosphatidylcholine lipid adducts reveals novel dissociation pathways. Int. J. Mass Spectrom. 2006, 252, 242–255. [Google Scholar] [CrossRef]
- Hsu, F.F.; Turk, J. Characterization of phosphatidylinositol, phosphatidylinositol-4-phosphate, and phosphatidylinositol-4,5-bisphosphate by electrospray ionization tandem mass spectrometry: A mechanistic study. J. Am. Soc. Mass Spectrom. 2000, 11, 986–999. [Google Scholar] [CrossRef]
- Hsu, F.F.; Turk, J. Studies on phosphatidylglycerol with triple quadrupole tandem mass spectrometry with electrospray ionization: Fragmentation processes and structural characterization. J. Am. Soc. Mass Spectrom. 2001, 12, 1036–1043. [Google Scholar] [CrossRef]
- Hsu, F.F.; Turk, J. Studies on phosphatidylserine by tandem quadrupole and multiple stage quadrupole ion-trap mass spectrometry with electrospray ionization: Structural characterization and the fragmentation processes. J. Am. Soc. Mass Spectrom. 2005, 16, 1510–1522. [Google Scholar] [CrossRef] [PubMed]
- Hsu, F.F.; Turk, J. Electrospray ionization with low-energy collisionally activated dissociation tandem mass spectrometry of glycerophospholipids: Mechanisms of fragmentation and structural characterization. J. Chromatogr. B 2009, 877, 2673–2695. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Liu, K.H.; Lee, S.Y.; Hong, S.J.; Cho, B.K.; Lee, H.; Lee, C.G.; Choi, H.K. Effects of light intensity and nitrogen starvation on glycerolipid, glycerophospholipid, and carotenoid composition in Dunaliella tertiolecta culture. PLoS ONE 2013, 8, e72415. [Google Scholar] [CrossRef] [PubMed]
- Lu, N.; Wei, D.; Chen, F.; Yang, S.T. Lipidomic profiling and discovery of lipid biomarkers in snow alga Chlamydomonas nivalis under salt stress. Eur. J. Lipid Sci. Technol. 2012, 114, 253–265. [Google Scholar] [CrossRef]
- Li, H.; Yan, X.; Xu, J.; Zhou, C. Precise identification of photosynthetic glycerolipids in microalga Tetraselmis chuii by UPLC-ESI-Q-TOF-MS. Sci. China C Life Sci. 2008, 51, 1101–1107. [Google Scholar] [CrossRef] [PubMed]
- Benning, C.; Huang, Z.H.; Gage, D.A. Accumulation of a novel glycolipid and a betaine lipid in cells of rhodobacter sphaeroides grown under phosphate limitation. Arch. Biochem. Biophys. 1995, 317, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Kind, T.; Meissen, J.K.; Yang, D.; Nocito, F.; Vaniya, A.; Cheng, Y.S.; VanderGheynst, J.S.; Fiehn, O. Qualitative analysis of algal secretions with multiple mass spectrometric platforms. J. Chromatogr. A 2012, 1244, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Roche, S.A.; Leblond, J.D. Betaine lipids in chlorarachniophytes. Phycol. Res. 2010, 58, 298–305. [Google Scholar] [CrossRef]
- Jung, H.R.; Sylvänne, T.; Koistinen, K.M.; Tarasov, K.; Kauhanen, D.; Ekroos, K. High throughput quantitative molecular lipidomics. Biochim. Biophys. Acta 2011, 1811, 925–934. [Google Scholar] [CrossRef] [PubMed]
- Vu, H.S.; Tamura, P.; Galeva, N.A.; Chaturvedi, R.; Roth, M.R.; Williams, T.D.; Wang, X.; Shah, J.; Welti, R. Direct infusion mass spectrometry of oxylipin-containing arabidopsis membrane lipids reveals varied patterns in different stress responses. Plant Physiol. 2012, 158, 324–339. [Google Scholar] [CrossRef] [PubMed]
- Nylund, G.M.; Weinberger, F.; Rempt, M.; Pohnert, G. Metabolomic assessment of induced and activated chemical defence in the invasive red alga Gracilaria vermiculophylla. PLoS ONE 2011, 6, e29359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumari, P.; Reddy, C.R.K.; Jha, B. Methyl jasmonate-induced lipidomic and biochemical alterations in the intertidal macroalga Gracilaria dura (Gracilariaceae, Rhodophyta). Plant Cell Physiol. 2015, 56, 1877–1889. [Google Scholar] [CrossRef] [PubMed]
- Ma, A.C.; Chen, Z.; Wang, T.; Song, N.; Yan, Q.; Fang, Y.C.; Guan, H.S.; Liu, H.B. Isolation of the molecular species of monogalactosyldiacylglycerols from brown edible seaweed Sargassum horneri and their inhibitory effects on triglyceride accumulation in 3T3-L1 adipocytes. J. Agric. Food Chem. 2014, 62, 11157–11162. [Google Scholar] [CrossRef] [PubMed]
- Oshima, M. The Newly Developed Lipid Database “LIPIDBANK for Web”, its’ contribution to the development of bioinformatics. Oleoscience 2004, 4, 329–334. [Google Scholar] [CrossRef]
- Yetukuri, L.; Ekroos, K.; Vidal-Puig, A.; Oresic, M. Informatics and computational strategies for the study of lipids. Mol. Biosyst. 2008, 4, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Caffrey, M.; Hogan, J. LIPIDAT: A database of lipid phase transition temperatures and enthalpy changes. DMPC data subset analysis. Chem. Phys. Lipids 1992, 61, 1–109. [Google Scholar] [CrossRef]
- Fahy, E.; Subramaniam, S.; Murphy, R.C.; Nishijima, M.; Raetz, C.R.H.; Shimizu, T.; Spener, F.; van Meer, G.; Wakelam, M.J.O.; Dennis, E.A. Update of the LIPID MAPS comprehensive classification system for lipids. J. Lipid Res. 2009, 50, S9–S14. [Google Scholar] [CrossRef] [PubMed]
- Sud, M.; Fahy, E.; Cotter, D.; Brown, A.; Dennis, E.A.; Glass, C.K.; Merrill, A.H.; Murphy, R.C.; Raetz, C.R.H.; Russell, D.W.; Subramaniam, S. LMSD: LIPID MAPS structure database. Nucleic Acids Res. 2007, 35, D527–D532. [Google Scholar] [CrossRef] [PubMed]
- Ejsing, C.S.; Duchoslav, E.; Sampaio, J.; Simons, K.; Bonner, R.; Thiele, C.; Ekroos, K.; Shevchenko, A. Automated identification and quantification of glycerophospholipid molecular species by multiple precursor ion scanning. Anal. Chem. 2006, 78, 6202–6214. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, R.; Nishijima, M.; Shimizu, T. Basic analytical systems for lipidomics by mass spectrometry in Japan. Methods Enzymol. 2007, 432, 185–211. [Google Scholar] [PubMed]
- Herzog, R.; Schuhmann, K.; Schwudke, D.; Sampaio, J.L.; Bornstein, S.R.; Schroeder, M.; Shevchenko, A. LipidXplorer: A software for consensual cross-platform lipidomics. PLoS ONE 2012, 7, e29851. [Google Scholar] [CrossRef] [PubMed]
- Husen, P.; Tarasov, K.; Katafiasz, M.; Sokol, E.; Vogt, J.; Baumgart, J.; Nitsch, R.; Ekroos, K.; Ejsing, C.S. Analysis of lipid experiments (ALEX): A software framework for analysis of high-resolution shotgun lipidomics data. PLoS ONE 2013, 8, e79736. [Google Scholar] [CrossRef] [PubMed]
- Tsugawa, H.; Cajka, T.; Kind, T.; Ma, Y.; Higgins, B.; Ikeda, K.; Kanazawa, M.; VanderGheynst, J.; Fiehn, O.; Arita, M. MS-DIAL: Data independent MS/MS deconvolution for comprehensive metabolome analysis. Nat. Methods 2015, 12, 523–526. [Google Scholar] [CrossRef] [PubMed]
- Kind, T.; Liu, K.H.; Lee, D.Y.; DeFelice, B.; Meissen, J.K.; Fiehn, O. LipidBlast in silico tandem mass spectrometry database for lipid identification. Nat. Methods 2013, 10, 755–758. [Google Scholar] [CrossRef] [PubMed]

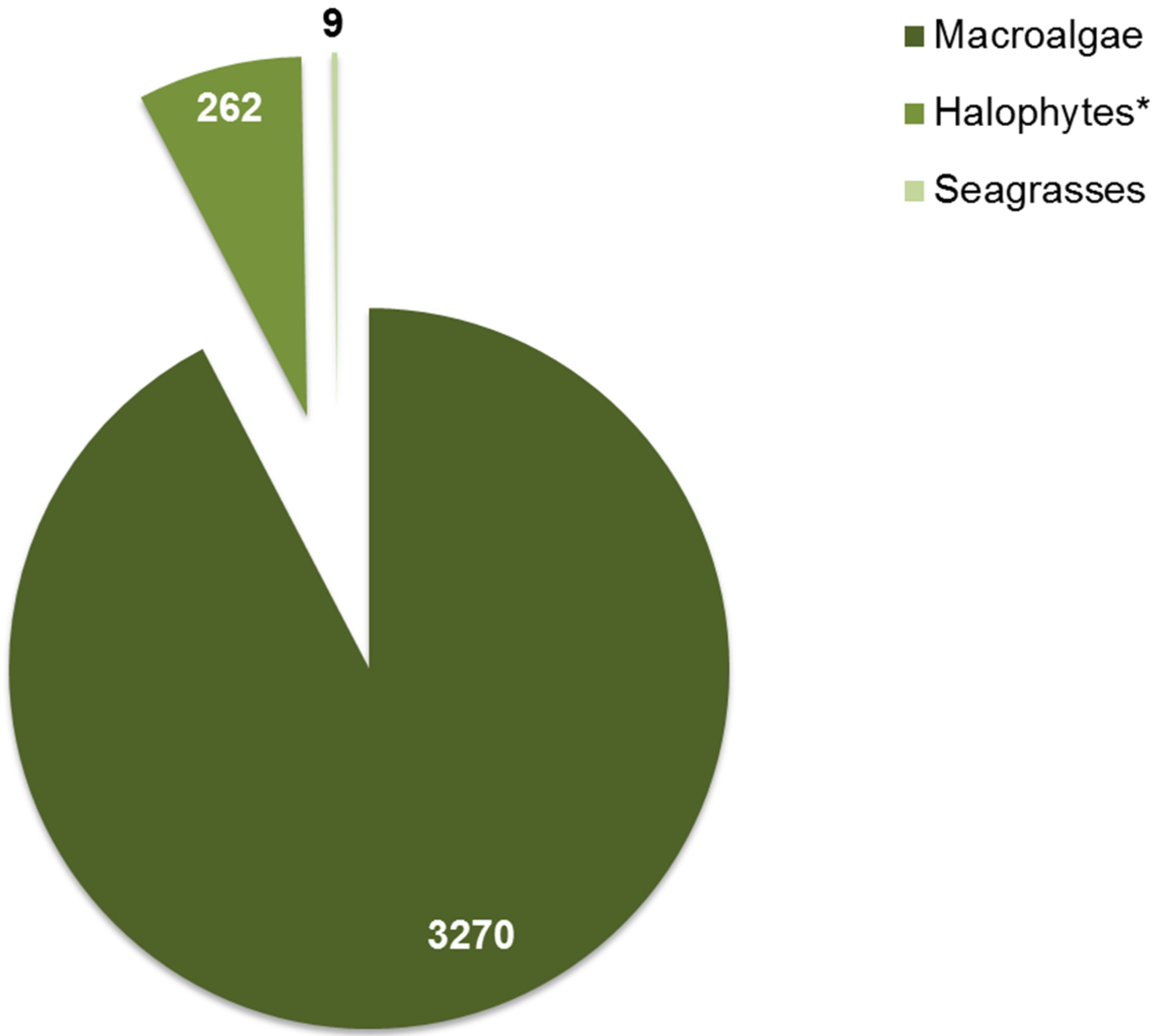





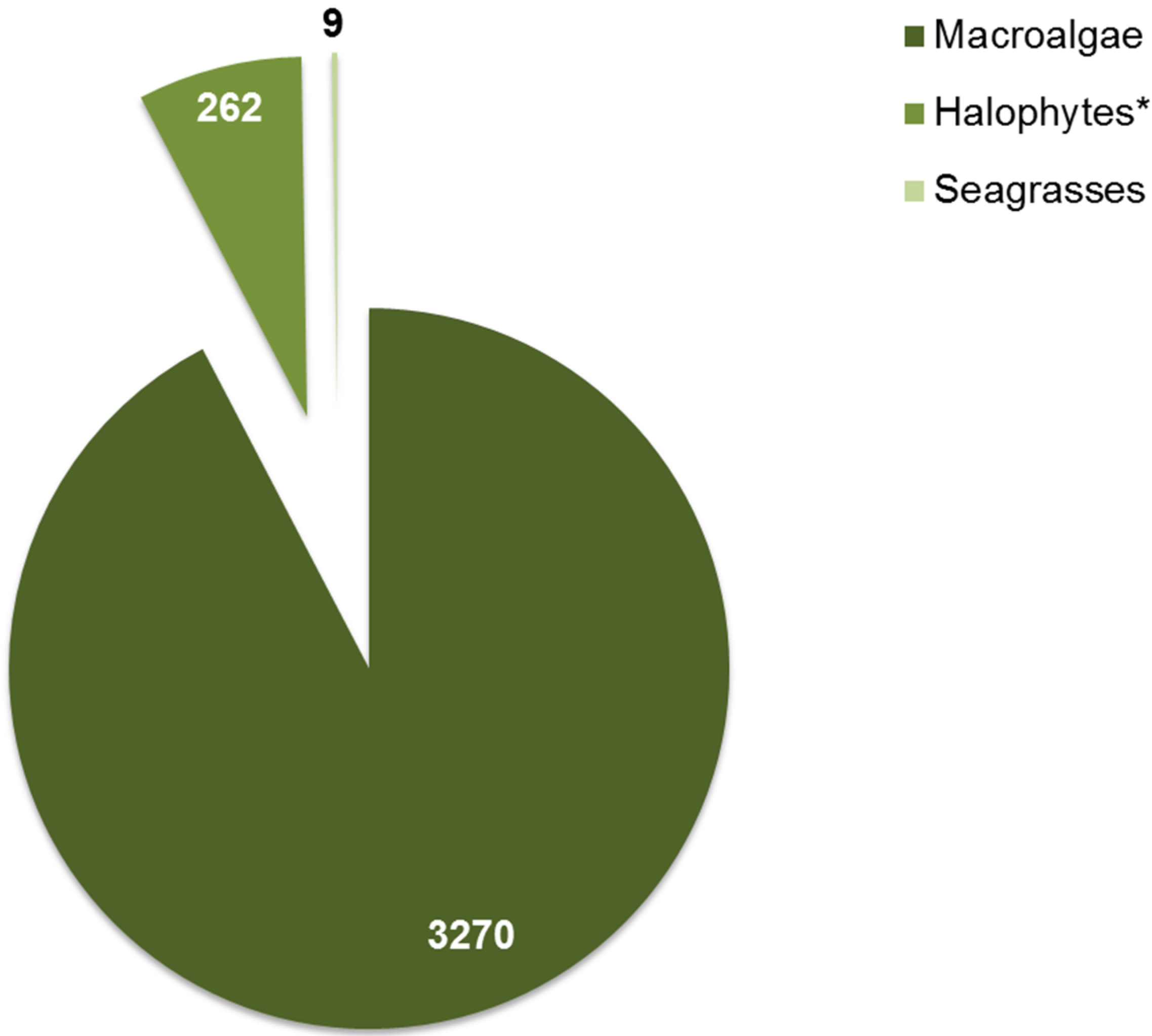
{kind=link}
{kind=link}
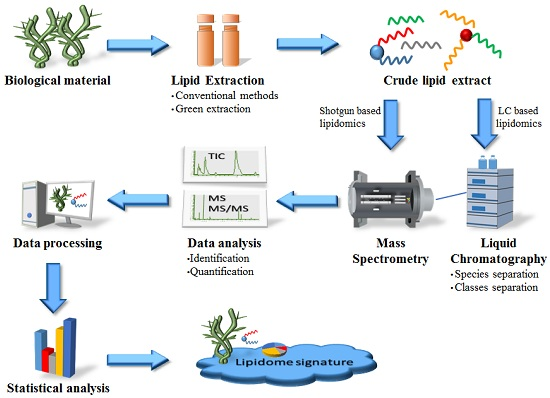
{kind=link}
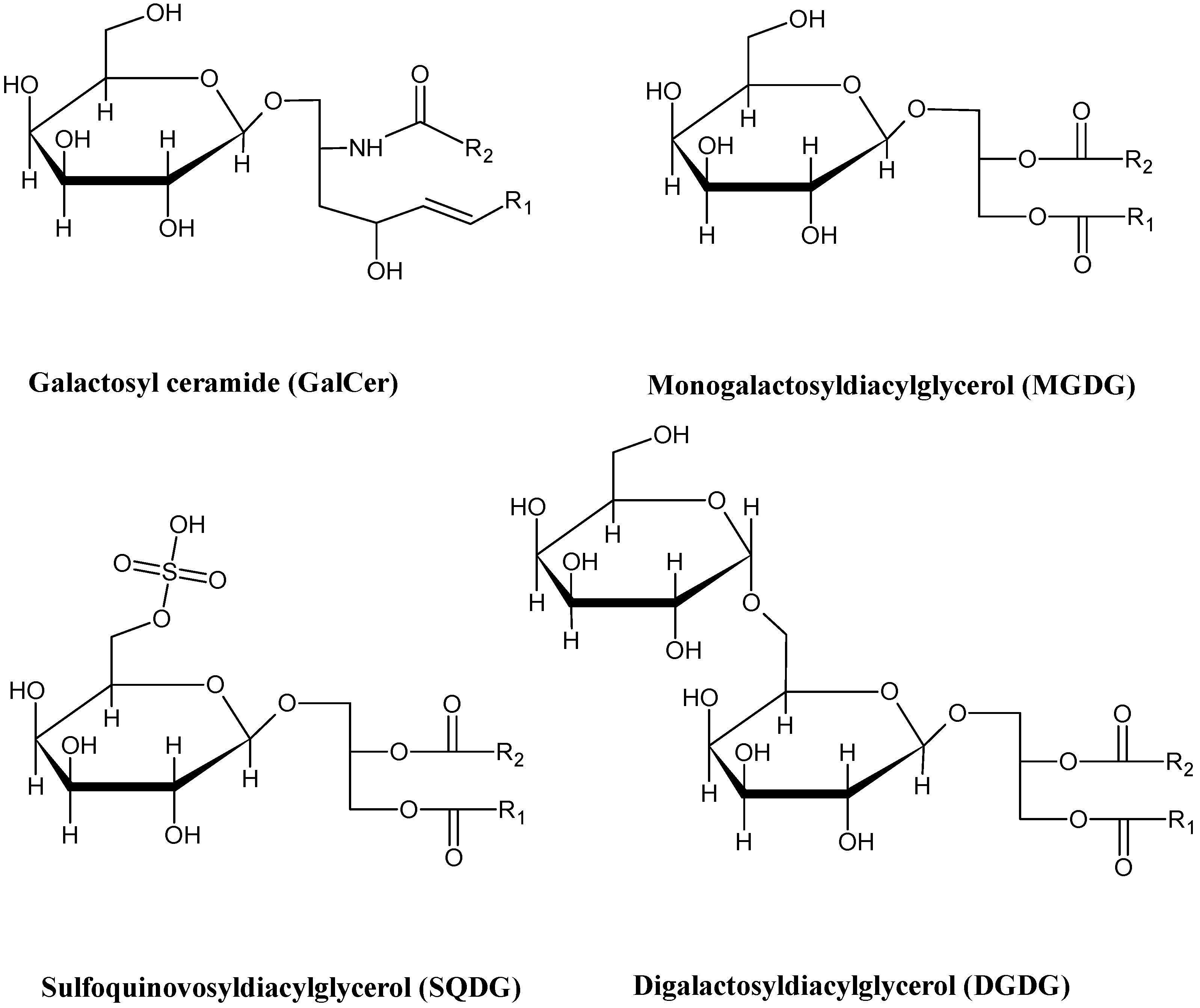{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Lipid Class/Extract | Bioactivity | Ref. |
|---|---|---|---|
| Green macroalgae | |||
| Ulva fasciata | SQDG | Antimicrobial (B. subtilis and E. coli) | [42] |
| Antitumor (MCF-7 and HEPG2 cells) | |||
| Ulva armaricana | DGDG (14:0/18:3) | Antitumor (NSCLC-N6 CELLS) | [50] |
| Red macroalgae | |||
| Chondria armata | MGDG (20:5/16:0) | Antifungal (C. albicans, | [43] |
| Antimicrobial (Klebsiella sp.) | |||
| Chondrus crispus (cultured) | Anti-inflammatory | [45] | |
| Galaxoura cylindriea | SQDG | Antimicrobial (B. subtilis and E. coli) | [42] |
| Antitumor (MCF-7 and HEPG2 cells) | |||
| Laurencia papillose | SQDG | Antimicrobial (B. subtilis and E. coli) | [42] |
| Antitumor (MCF-7 and HEPG2 cells) | |||
| Osmundaria obtusiloba | SQDG | anti-viral (HSV-1 and HSV-2) | [44] |
| Palmaria palmate | SQDG, PG | Anti-inflammatory | [46] |
| Solieria chordalis | MGDG (14:0/16:1) | Anti-tumor (NSCLC-N6 CELLS) | [50] |
| Brown macroalgae | |||
| Dilophys fasciola | SQDG | Antimicrobial (B. subtilis and E. coli) | [42] |
| Antitumor (MCF-7 and HEPG2 cells) | |||
| Fucus spiralis | MGDG | Anti-inflammatory | [47] |
| Sargassum horneri | SQDG, DGDG | Antitumor (Caco-2 cell) | [51] |
| Sargassum thumbergii | MGDG (20:5/18:4) and (18:3/18:4) | Antifungal (Candida albicans) | [52] |
| Sargassum wightii | SQDG | Antimicrobial (X. oryzae pv.) | [53] |
| Taonia atomaria | SQDG | Antimicrobial (B. subtilis and E. coli) | [42] |
| Antitumor (MCF-7 and HEPG2 cells) | |||
| Seagrass | |||
| Cymodocea serrulata | Chloroform extract | Antimicrobial (P. aeruginosa and K. pneumoniae) | [54] |
| Halophila ovalis | Methanolic extract | Antimicrobial (E. coli) | [55] |
| Halophila stipulacea | Methanolic extract | Antimicrobial (V. cholera) | [54] |
| Chloroform extract | Antimicrobial (S. bodii) | ||
| Methanolic extract | Antimicrobial (S. aureus) | [55] | |
| Halodule pinifolia | Methanolic extract | Antimicrobial (S. aureus, K. pneumoniae and S. paratyphi) | [54] |
| Zostera capensis | Methanolic extract | Antimicrobial (S. paratyphi) | [55] |
| Ethyl acetate extract | Antimicrobial (B. cereus) | ||
| Ethyl acetate extract | Antimicrobial (S. typhimurium) | ||
| Zostera japonica | Methanolic extract | Anti-inflammatory | [56] |
| Lipid Class | Detect Ions in MS | Precursor Ion Scan | Neutral Loss Scan | |||
|---|---|---|---|---|---|---|
| Negative | Positive | Negative (m/z) | Positive (m/z) | Negative (Da) | Positive (Da) | |
| Phosphatidylcholine (PC) | [M + Ac-H]− | [M + H]+, [M + Na]+ | - | 184 | - | - |
| Phosphatidylethanolamine (PE) | [M − H]− | [M + H]+, [M + Na]+ | - | - | - | 141 |
| Phosphatidylglycerol (PG) | [M − H]− | [M + NH4]+, [M + Na]+ | - | - | 74 | - |
| Phosphatidylinositol (PI) | [M − H]− | [M + NH4]+ | 241 | - | - | - |
| Phosphatidylserine (PS) | [M − H]− | [M + H]+ | - | - | 87 | 185 |
| Monogalactosyldiacylglycerol (MGDG) | [M − H]− | [M + NH4]+, [M + Na]+ | - | 243 | - | 179 |
| Digalactosyldiacylglycerol (DGDG) | [M − H]− | [M + NH4]+, [M + Na]+ | - | 347 | - | 162 |
| 365 | 341 | |||||
| Sulfoquinovosildiacylglycerol (SQDG) | [M − H]− | [M + NH4]+, [M + Na]+ | 225 | - | - | - |
| Ceramide (Cer) | [M − H]− | [M + H]+, [M + NH4]+, [M + Na]+ | 264 | |||
| Galactosylceramide (GalCer) | [M − H]− | [M + H]+, [M + NH4]+, [M + Na]+ | - | 264 | - | 162 |
| 180 | ||||||
| Inositolphosphoceramide (IPC) | [M − H]− | [M + H]+, [M + NH4]+, [M + Na]+ | - | 223 | - | 162 |
| 241 | 180 | |||||
| 259 | ||||||
| Diacylglyceryl-N,N,N-trimethylhomoserine (DGTS) | [M + H]+ | - | 236 | - | 87 | |
| 74 | ||||||
| 59 | ||||||
| Diacylglycerylhydroxymethyl-N,N,N-trimetyl β-alanine (DGTA) | [M + H]+ | 236 | 87 | |||
| 74 | ||||||
| 59 | ||||||
| Diacylglycerylcarboxyhydroxy methylcholine (DGCC) | [M + H]+ | 104 | ||||
| Species Name | MS Approach | Extraction Method | Glycolipids | Phospholipids | Betaine Lipids | Ref. |
|---|---|---|---|---|---|---|
| Green macroalgae | ||||||
| Codium tomentosum | HILIC LC-MSn ESI-LXQ-IT | CH3OH | SQDG (20), SQMG (4), DGDG (22), MGDG (10) | PG(22), LPG(8) PA(9), PI (13), LPC (11), PC(62) | DGTS (43), MGTS (16) | [68] |
| Enteromorpha intestinalis | HILIC-LC-MS ESI/IT-TOF | Folch | SQDG (1), SQMG (1) | [107] | ||
| Ulva armaricana | LC-IT-TOF | CH3OH:CHCl3 (1:1, v/v) | DGDG | [50] | ||
| Ulva fasciata | LC-MSn ESI-QqQ | CH3OH:CHCl3 (2:1, v/v) | SQDG (1), SQMG (1) | [42] | ||
| Ulva lactuca | ESI-Q-TOF-MSn | Bligh and Dyer | MGDG, DGDG, SQDG | PG, LPG, PC, LPC, PS, PA, PI | DGTS | [57] |
| Ulva rigida | HILIC-LC-MS ESI/IT-TOF | Folch | SQDG (1) | PC (4), LPE (1) | [107] | |
| Red macroalgae | ||||||
| Asparagopsis taxiformis | HILIC-LC-MS ESI/IT-TOF | Folch | SQMG | [107] | ||
| Chondria armata | offline TLC-ESI-QTOF-MSn | CH3OH + isolation in CHCl3 | MGMG (2), MGDG (3), SQMG (2) | PG (4) | [43] | |
| Chondrus crispus (cultured) | HILIC LC-MSn ESI-LXQ-IT | MTBE:CH3OH | DGDG (19), SQDG (14) | PG (18), LPG (2), PC (60), LPC (8), PA (14) | DGTS (14) | [37] |
| Chondrus crispus (cultured) | Off-line LC-Q-MSn | CH3OH and several fraction based on EtOAc blends | MGDG (6), DGDG (2) | [45] | ||
| Galaxoura cylindriea | LC-MSn ESI-QqQ | CH3OH:CHCl3 (2:1, v/v) | SQMG, SQDG | [42] | ||
| Laurencia papillose | LC-MSn ESI-QqQ | CH3OH:CHCl3 (2:1, v/v) | SQMG, SQDG | [42] | ||
| Palmaria palmata | Reverse-phase LC-Q-MSn | CH3OH:CHCl3 (1:1, v/v) | MGDG (2), DGDG (3), SQDG (2) | PG (2), PE (1) | [46] | |
| Pterocodiella capillacea | HILIC-LC-MS-ESI/IT-TOF | Folch | DGDG, SQDG, SQMG | PG, PC, PI, LPI, PS, LPE | [107] | |
| Osmundaria obtusiloba | Off-line API-ESI-QqQ-MSn | MGDG (1), DGDG (1), SQDG (1), SQMG (1) | [44] | |||
| Solieria chordalis | LC-IT-TOF | CH3OH:CHCl3 (1:1, v/v) | MGDG | [50] | ||
| Brown macroalgae | ||||||
| Colpomenia sinuosa | HILIC-LC-MS ESI/IT-TOF | Folch | SQMG | PC, LPE, PI | [107] | |
| Cystoseyra brachicarpa | HILIC-LC-MS ESI/IT-TOF | Folch | SQDG | PG, PC | [107] | |
| Dictoyota dicotoma | HILIC-LC-MS ESI/IT-TOF | Folch | SQDG, SQMG | PC, LPE | [107] | |
| Dilophys fasciola | LC-MSn ESI-QqQ | CH3OH:CHCl3 (2:1, v/v) | SQDG (1), SQMG (1) | [42] | ||
| Fucus sp. | ESI-LTQ-MSn | CH3OH + fractions solvent/solvent partitioning | MGDG (2) | [47] | ||
| Sargassum sp. | Reverse-phase LC-ESI-QIT-MS | EtOAc | MGDG (10) | [145] | ||
| FAB-MSn | CH3OH:nBuOH | MGDG (2) | [52] | |||
| offline TLC-ESI-QTOF-MSn | CH3OH:CHCl3 (1:2 and 2:1, v/v) | SQDG | [39] | |||
| Stypocaulum scoparium | HILIC-LC-MS ESI/IT-TOF | Folch | PG, PC, PS | [107] | ||
| Taonia atomaria | LC-MSn ESI-QqQ | CH3OH:CHCl3 (2:1, v/v) | SQDG, SQMG | [42] | ||
| Halophytes | ||||||
| Aster tripolium | LC-TOF MS | CH3OH/CHCl3/H2O (65:25:4, v/v/v) | SQDG | [108] | ||
| Sesuvium portulacastrum | LC-TOF MS | CH3OH/CHCl3/H2O (65:25:4, v/v/v) | SQDG | [108] | ||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maciel, E.; Leal, M.C.; Lillebø, A.I.; Domingues, P.; Domingues, M.R.; Calado, R. Bioprospecting of Marine Macrophytes Using MS-Based Lipidomics as a New Approach. Mar. Drugs 2016, 14, 49. https://doi.org/10.3390/md14030049
Maciel E, Leal MC, Lillebø AI, Domingues P, Domingues MR, Calado R. Bioprospecting of Marine Macrophytes Using MS-Based Lipidomics as a New Approach. Marine Drugs. 2016; 14(3):49. https://doi.org/10.3390/md14030049
Chicago/Turabian StyleMaciel, Elisabete, Miguel Costa Leal, Ana Isabel Lillebø, Pedro Domingues, Maria Rosário Domingues, and Ricardo Calado. 2016. "Bioprospecting of Marine Macrophytes Using MS-Based Lipidomics as a New Approach" Marine Drugs 14, no. 3: 49. https://doi.org/10.3390/md14030049