
In-Depth Characterization of bZIP Genes in the Context of Endoplasmic Reticulum (ER) Stress in Brassica campestris ssp. chinensis

 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Total 112 BcbZIP Genes Were Identified and Their Physicochemical Properties Were Evaluated
2.2. Phylogenetic Analysis Divided BcbZIP Genes into 10 Sub Families
2.3. The Chromosomal Location and Duplication Events of BcbZIP Genes
2.4. Divergence Analysis Investigated How BcbZIP Genes Have Evolved and Diversified over Time
2.5. Gene Structure, Domains, and Motifs Analysis Provide a Genetic Basis of Biological Processes to Infer the Potential Functions of BcbZIP Genes
2.6. Cis Elements Analysis Indicate the Potential of BcbZIP Genes to Mediate Responses to a Diverse Range of Environmental Stimuli
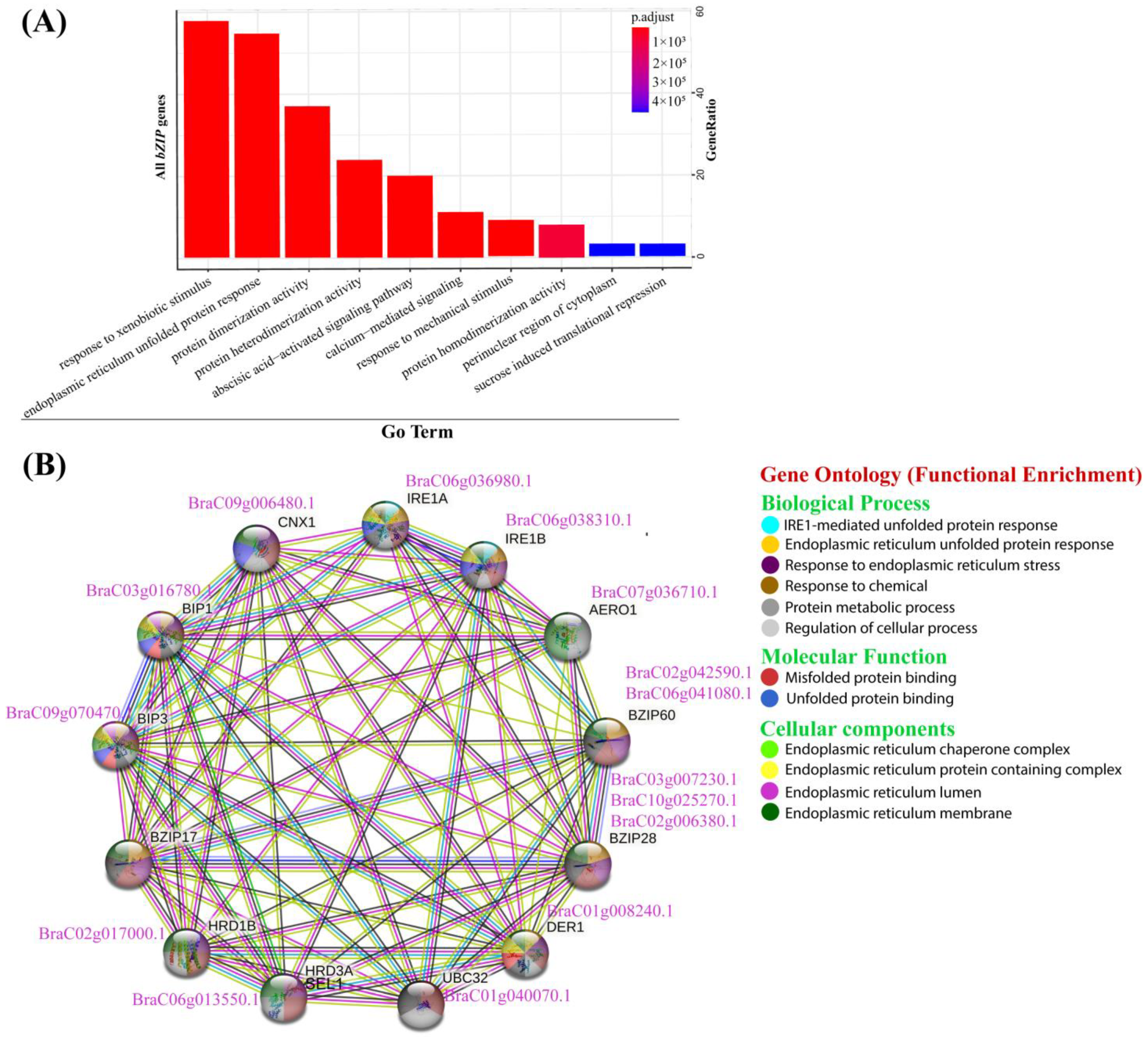
2.7. GO Enrichment Analysis Suggest the Crucial Roles of BcbZIP Genes in Regulating Multiple Cellular Processes
2.8. Co-Expression Network Conveyed a Strong Association between BcbZIP Genes with ER Stress Responsive Genes
2.9. Expression Profiles of BcbZIP Genes during ER Stress in Pak Choi
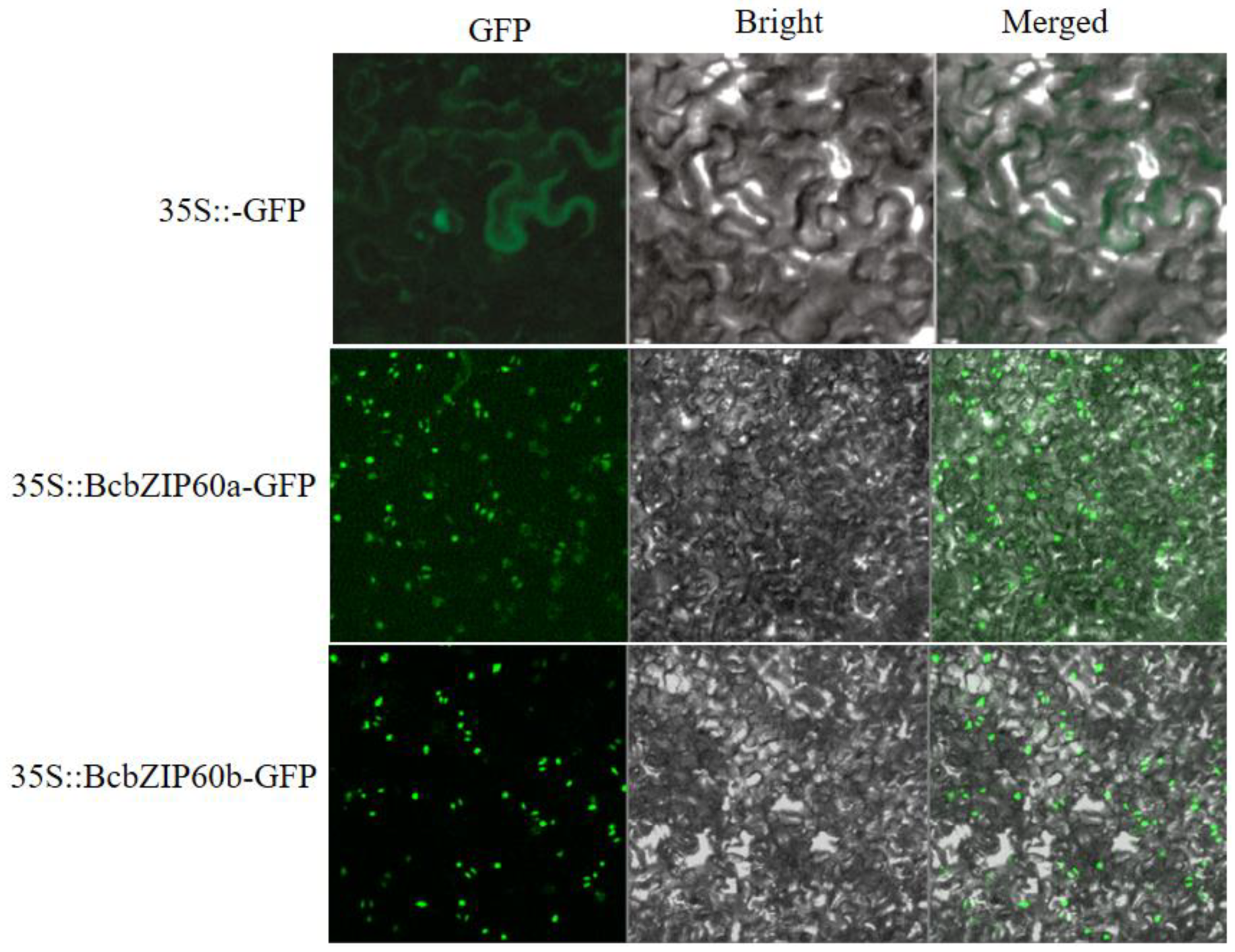
2.10. Subcellular Localization Revealed the Presence of BcbZIP60a and BcbZIP60b in Nucleus
2.11. Y2H Displayed a Strong Protein–Protein Interaction of BcbZIP60a and BcbZIP60b with BcBiP3
2.12. BiFC Assays Indicate the Interaction of BcbZIP60a and BcbZIP60b with BcBiP3 at Multiple Points
3. Discussion
4. Materials and Methods
4.1. Study Layout and Plant Growth Conditions

4.2. Identification and Physicochemical Analysis of bZIP Genes
4.3. Multiple Sequence Alignment and Phylogenetic Analysis of BcbZIP Genes
4.4. Chromosomal Location and Duplication of BcbZIP Genes
4.5. Gene Structure, Domains, and Motifs
4.6. Divergence, Cis Elements, and GO Enrichment Analysis
4.7. Co-Expression Network Analysis of BcbZIP Genes with ER Stress Responsive Genes
4.8. Expression Analysis of BcbZIP in Response to ER Stress
4.9. Yeast Two Hybrid (Y2H) Assay of BcbZIP60 with ER Chaperone BcBiP3
4.10. BiFC Assay of BcbZIP60 with ER Chaperone BcBiP3
4.11. Subcellular Localization of BcbZIP60
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Das, P.; Lakra, N.; Nutan, K.K.; Singla-Pareek, S.L.; Pareek, A. A Unique bZIP Transcription Factor Imparting Multiple Stress Tolerance in Rice. Rice 2019, 12, 58. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Li, W.; Chen, X. Transcription Factor Is Not Just a Transcription Factor. Trends Plant Sci. 2022, 27, 1087–1089. [Google Scholar] [CrossRef]
- Pérez-Rodríguez, P.; Riaño-Pachón, D.M.; Corrêa, L.G.G.; Rensing, S.A.; Kersten, B.; Mueller-Roeber, B. PlnTFDB: Updated Content and New Features of the Plant Transcription Factor Database. Nucleic Acids Res. 2010, 38, D822–D827. [Google Scholar] [CrossRef] [PubMed]
- Duan, L.; Mo, Z.; Fan, Y.; Li, K.; Yang, M.; Li, D.; Ke, Y.; Zhang, Q.; Wang, F.; Fan, Y.; et al. Genome-Wide Identification and Expression Analysis of the bZIP Transcription Factor Family Genes in Response to Abiotic Stress in Nicotiana tabacum L. BMC Genom. 2022, 23, 318. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Tai, B.; Hussain, A.; Jahan, I.; Yang, B.; Xing, F. Genome-Wide Identification and Expression Analysis of the Basic Leucine Zipper (bZIP) Transcription Factor Gene Family in Fusarium graminearum. Genes 2022, 13, 607. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Fujita, Y.; Maruyama, K.; Mogami, J.; Todaka, D.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Four Arabidopsis AREB/ABF Transcription Factors Function Predominantly in Gene Expression Downstream of SnRK2 Kinases in Abscisic Acid Signalling in Response to Osmotic Stress. Plant Cell Environ. 2015, 38, 35–49. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, L.; Ye, T.; Lu, Y.; Chen, X.; Wu, Y. The Inhibitory Effect of ABA on Floral Transition Is Mediated by ABI5 in Arabidopsis. J. Exp. Bot. 2013, 64, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Matiolli, C.C.; Tomaz, J.P.; Duarte, G.T.; Prado, F.M.; Del Bem, L.E.V.; Silveira, A.B.; Gauer, L.; Corrêa, L.G.G.; Drumond, R.D.; Viana, A.J.C.; et al. The Arabidopsis bZIP Gene AtbZIP63 Is a Sensitive Integrator of Transient Abscisic Acid and Glucose Signals. Plant Physiol. 2011, 157, 692–705. [Google Scholar] [CrossRef] [PubMed]
- Stotz, H.U.; Mueller, S.; Zoeller, M.; Mueller, M.J.; Berger, S. TGA Transcription Factors and Jasmonate-Independent COI1 Signalling Regulate Specific Plant Responses to Reactive Oxylipins. J. Exp. Bot. 2013, 64, 963–975. [Google Scholar] [CrossRef] [PubMed]
- Assunção, A.G.L.; Herrero, E.; Lin, Y.-F.; Huettel, B.; Talukdar, S.; Smaczniak, C.; Immink, R.G.H.; van Eldik, M.; Fiers, M.; Schat, H.; et al. Arabidopsis thaliana Transcription Factors bZIP19 and bZIP23 Regulate the Adaptation to Zinc Deficiency. Proc. Natl. Acad. Sci. USA 2010, 107, 10296–10301. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, K.; Weltmeier, F.; Ehlert, A.; Weiste, C.; Stahl, M.; Harter, K.; Dröge-Laser, W. Heterodimers of the Arabidopsis Transcription Factors bZIP1 and bZIP53 Reprogram Amino Acid Metabolism during Low Energy Stress. Plant Cell 2011, 23, 381–395. [Google Scholar] [CrossRef] [PubMed]
- Zander, M.; La Camera, S.; Lamotte, O.; Métraux, J.-P.; Gatz, C. Arabidopsis thaliana Class-II TGA Transcription Factors Are Essential Activators of Jasmonic Acid/Ethylene-Induced Defense Responses. Plant J. 2010, 61, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Yang, H.; Yan, Y.; Wei, Y.; Tie, W.; Ding, Z.; Zuo, J.; Peng, M.; Li, K. Genome-Wide Characterization and Analysis of bZIP Transcription Factor Gene Family Related to Abiotic Stress in Cassava. Sci. Rep. 2016, 6, 22783. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Xiao, L.; Zhao, J.; Hu, Z.; Zhou, Y.; Liu, S.; Wu, H.; Zhou, Y. Comprehensive Genomic Analysis and Expression Profile of Hsp70 Gene Family Related to Abiotic and Biotic Stress in Cucumber. Horticulturae 2023, 9, 1057. [Google Scholar] [CrossRef]
- Du, K.; Huang, J.; Wang, W.; Zeng, Y.; Xuecao, L.; Zhao, F. Monitoring Low-Temperature Stress in Winter Wheat Using TROPOMI Solar-Induced Chlorophyll Fluorescence. IEEE Trans. Geosci. Remote Sens. 2024, 62, 1–11. [Google Scholar] [CrossRef]
- Chambers, J.E.; Marciniak, S.J. Cellular Mechanisms of Endoplasmic Reticulum Stress Signaling in Health and Disease. 2. Protein Misfolding and ER Stress. Am. J. Physiol.-Cell Physiol. 2014, 307, C657–C670. [Google Scholar] [CrossRef] [PubMed]
- Bailey, D.; O’Hare, P. Transmembrane bZIP Transcription Factors in ER Stress Signaling and the Unfolded Protein Response. Antioxid. Redox Signal. 2007, 9, 2305–2322. [Google Scholar] [CrossRef] [PubMed]
- Hetz, C.; Zhang, K.; Kaufman, R.J. Mechanisms, Regulation and Functions of the Unfolded Protein Response. Nat. Rev. Mol. Cell Biol. 2020, 21, 421–438. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-X.; Howell, S.H. Managing the Protein Folding Demands in the Endoplasmic Reticulum of Plants. New Phytol. 2016, 211, 418–428. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, H.; Brandizzi, F.; Verchot, J.; Wang, A. The UPR Branch IRE1-bZIP60 in Plants Plays an Essential Role in Viral Infection and Is Complementary to the Only UPR Pathway in Yeast. PLoS Genet. 2015, 11, e1005164. [Google Scholar] [CrossRef] [PubMed]
- Ruberti, C.; Kim, S.-J.; Stefano, G.; Brandizzi, F. Unfolded Protein Response in Plants: One Master, Many Questions. Curr. Opin. Plant Biol. 2015, 27, 59–66. [Google Scholar] [CrossRef]
- Wan, S.; Jiang, L. Endoplasmic Reticulum (ER) Stress and the Unfolded Protein Response (UPR) in Plants. Protoplasma 2016, 253, 753–764. [Google Scholar] [CrossRef] [PubMed]
- Hollien, J.; Lin, J.H.; Li, H.; Stevens, N.; Walter, P.; Weissman, J.S. Regulated Ire1-Dependent Decay of Messenger RNAs in Mammalian Cells. J. Cell Biol. 2009, 186, 323–331. [Google Scholar] [CrossRef]
- Mishiba, K.; Nagashima, Y.; Suzuki, E.; Hayashi, N.; Ogata, Y.; Shimada, Y.; Koizumi, N. Defects in IRE1 Enhance Cell Death and Fail to Degrade mRNAs Encoding Secretory Pathway Proteins in the Arabidopsis Unfolded Protein Response. Proc. Natl. Acad. Sci. USA 2013, 110, 5713–5718. [Google Scholar] [CrossRef]
- Deng, Y.; Humbert, S.; Liu, J.-X.; Srivastava, R.; Rothstein, S.J.; Howell, S.H. Heat Induces the Splicing by IRE1 of a mRNA Encoding a Transcription Factor Involved in the Unfolded Protein Response in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 7247–7252. [Google Scholar] [CrossRef]
- Liu, J.-X.; Srivastava, R.; Che, P.; Howell, S.H. An Endoplasmic Reticulum Stress Response in Arabidopsis Is Mediated by Proteolytic Processing and Nuclear Relocation of a Membrane-Associated Transcription Factor, bZIP28. Plant Cell 2007, 19, 4111–4119. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, R.; Deng, Y.; Shah, S.; Rao, A.G.; Howell, S.H. BINDING PROTEIN Is a Master Regulator of the Endoplasmic Reticulum Stress Sensor/Transducer bZIP28 in Arabidopsis. Plant Cell 2013, 25, 1416–1429. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-X.; Howell, S.H. bZIP28 and NF-Y Transcription Factors Are Activated by ER Stress and Assemble into a Transcriptional Complex to Regulate Stress Response Genes in Arabidopsis. Plant Cell 2010, 22, 782–796. [Google Scholar] [CrossRef] [PubMed]
- Howell, S.H. Endoplasmic Reticulum Stress Responses in Plants. Annu. Rev. Plant Biol. 2013, 64, 477–499. [Google Scholar] [CrossRef]
- Wang, H.; Niu, H.; Zhai, Y.; Lu, M. Characterization of BiP Genes from Pepper (Capsicum annuum L.) and the Role of CaBiP1 in Response to Endoplasmic Reticulum and Multiple Abiotic Stresses. Front. Plant Sci. 2017, 8, 1122. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Srivastava, R.; Howell, S.H.; Bassham, D.C. Activation of Autophagy by Unfolded Proteins during Endoplasmic Reticulum Stress. Plant J. 2016, 85, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Alvim, F.C.; Carolino, S.M.B.; Cascardo, J.C.M.; Nunes, C.C.; Martinez, C.A.; Otoni, W.C.; Fontes, E.P.B. Enhanced Accumulation of BiP in Transgenic Plants Confers Tolerance to Water Stress. Plant Physiol. 2001, 126, 1042–1054. [Google Scholar] [CrossRef] [PubMed]
- Liebrand, T.W.H.; Kombrink, A.; Zhang, Z.; Sklenar, J.; Jones, A.M.E.; Robatzek, S.; Thomma, B.P.H.J.; Joosten, M.H.A.J. Chaperones of the Endoplasmic Reticulum Are Required for Ve1-Mediated Resistance to Verticillium. Mol. Plant Pathol. 2014, 15, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, H.H.; Brustolini, O.J.B.; Pimenta, M.R.; Mendes, G.C.; Gouveia, B.C.; Silva, P.A.; Silva, J.C.F.; Mota, C.S.; Soares-Ramos, J.R.L.; Fontes, E.P.B. The Molecular Chaperone Binding Protein BiP Prevents Leaf Dehydration-Induced Cellular Homeostasis Disruption. PLoS ONE 2014, 9, e86661. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, G.-F.; Ma, L.-M.; Liu, T.-K.; Zhang, C.-W.; Xiao, D.; Zheng, H.-K.; Chen, F.; Hou, X.-L. A Chromosome-Level Reference Genome of Non-Heading Chinese Cabbage [Brassica campestris (syn. Brassica rapa) ssp. chinensis]. Hortic. Res. 2020, 7, 212. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Li, Y.; Liu, T.; Duan, W.; Huang, Z.; Wang, L.; Tan, H.; Hou, X. Genes Associated with Agronomic Traits in Non-Heading Chinese Cabbage Identified by Expression Profiling. BMC Plant Biol. 2014, 14, 71. [Google Scholar] [CrossRef] [PubMed]
- Ayaz, A.; Jalal, A.; Qian, Z.; Khan, K.A.; Liu, L.; Hu, C.; Li, Y.; Hou, X. Investigating the Effects of Tauroursodeoxycholic Acid (TUDCA) in Mitigating Endoplasmic Reticulum Stress and Cellular Responses in Pak Choi. Physiol. Plant. 2024, 176, e14246. [Google Scholar] [CrossRef] [PubMed]
- Jalal, A.; Ali, Q.; Manghwar, H.; Zhu, D. Identification, Phylogeny, Divergence, Structure, and Expression Analysis of A20/AN1 Zinc Finger Domain Containing Stress-Associated Proteins (SAPs) Genes in Jatropha curcas L. Genes 2022, 13, 1766. [Google Scholar] [CrossRef] [PubMed]
- Jalal, A.; Sun, J.; Chen, Y.; Fan, C.; Liu, J.; Wang, C. Evolutionary Analysis and Functional Identification of Clock-Associated PSEUDO-RESPONSE REGULATOR (PRRs) Genes in the Flowering Regulation of Roses. Int. J. Mol. Sci. 2022, 23, 7335. [Google Scholar] [CrossRef]
- Dong, Y.; Lu, J.; Liu, J.; Jalal, A.; Wang, C. Genome-Wide Identification and Functional Analysis of JmjC Domain-Containing Genes in Flower Development of Rosa chinensis. Plant Mol. Biol. 2020, 102, 417–430. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Chai, F.; Wang, Y.; Jiang, J.; Duan, W.; Wang, Y.; Wang, F.; Li, S.; Wang, L. Genome-Wide Identification and Classification of HSF Family in Grape, and Their Transcriptional Analysis under Heat Acclimation and Heat Stress. Hortic. Plant J. 2018, 4, 133–143. [Google Scholar] [CrossRef]
- Bailey, T.L.; Williams, N.; Misleh, C.; Li, W.W. MEME: Discovering and Analyzing DNA and Protein Sequence Motifs. Nucleic Acids Res. 2006, 34, W369–W373. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhang, Y.; Zhang, Z.; Zhu, J.; Yu, J. KaKs_Calculator 2.0: A Toolkit Incorporating Gamma-Series Methods and Sliding Window Strategies. Genom. Proteom. Bioinform. 2010, 8, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a Database of Plant Cis-Acting Regulatory Elements and a Portal to Tools for in Silico Analysis of Promoter Sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING V11: Protein–Protein Association Networks with Increased Coverage, Supporting Functional Discovery in Genome-Wide Experimental Datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Xia, J.; Jiang, Y.; Bao, Y.; Chen, H.; Wang, D.; Zhang, D.; Yu, J.; Cang, J. Genome-Wide Identification and Analysis of bZIP Gene Family and Resistance of TaABI5 (TabZIP96) under Freezing Stress in Wheat (Triticum aestivum). Int. J. Mol. Sci. 2022, 23, 2351. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Cao, H.; Huang, B.; Zheng, X.; Liang, K.; Wang, G.-L.; Sun, X. The WRKY10-VQ8 Module Safely and Effectively Regulates Rice Thermotolerance. Plant Cell Environ. 2022, 45, 2126–2144. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ma, B.; Wang, C.; Chen, X.; Ruan, Y.-L.; Yuan, Y.; Ma, F.; Li, M. MdWRKY126 Modulates Malate Accumulation in Apple Fruit by Regulating Cytosolic Malate Dehydrogenase (MdMDH5). Plant Physiol. 2022, 188, 2059–2072. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Sun, J.; Chen, Y.; Zhu, P.; Zhang, L.; Wu, S.; Ma, D.; Cao, Q.; Li, Z.; Xu, T. Genome-Wide Identification, Structural and Gene Expression Analysis of the bZIP Transcription Factor Family in Sweet Potato Wild Relative Ipomoea Trifida. BMC Genet. 2019, 20, 41. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Gao, S.; Tang, Y.; Li, L.; Zhang, F.; Feng, B.; Fang, Z.; Ma, L.; Zhao, C. Genome-Wide Identification and Evolutionary Analyses of bZIP Transcription Factors in Wheat and Its Relatives and Expression Profiles of Anther Development Related TabZIP Genes. BMC Genom. 2015, 16, 976. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhou, J.; Zhang, B.; Vanitha, J.; Ramachandran, S.; Jiang, S.-Y. Genome-Wide Expansion and Expression Divergence of the Basic Leucine Zipper Transcription Factors in Higher Plants with an Emphasis on SorghumF. J. Integr. Plant Biol. 2011, 53, 212–231. [Google Scholar] [CrossRef] [PubMed]
- Nijhawan, A.; Jain, M.; Tyagi, A.K.; Khurana, J.P. Genomic Survey and Gene Expression Analysis of the Basic Leucine Zipper Transcription Factor Family in Rice. Plant Physiol. 2008, 146, 333–350. [Google Scholar] [CrossRef] [PubMed]
- Wei, K.; Chen, J.; Wang, Y.; Chen, Y.; Chen, S.; Lin, Y.; Pan, S.; Zhong, X.; Xie, D. Genome-Wide Analysis of bZIP-Encoding Genes in Maize. DNA Res. 2012, 19, 463–476. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chu, Z. Genome-Wide Evolutionary Characterization and Analysis of bZIP Transcription Factors and Their Expression Profiles in Response to Multiple Abiotic Stresses in Brachypodium Distachyon. BMC Genom. 2015, 16, 227. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Xu, D.; Jia, L.; Huang, X.; Ma, G.; Wang, S.; Zhu, M.; Zhang, A.; Guan, M.; Lu, K.; et al. Genome-Wide Identification and Structural Analysis of bZIP Transcription Factor Genes in Brassica napus. Genes 2017, 8, 288. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Zhu, W.; Hu, X.; Sun, C.; Li, Y.; Wang, D.; Wang, Q.; Pei, G.; Zhang, Y.; Guo, A.; et al. Genome-Wide Analysis of the bZIP Gene Family Identifies Two ABI5-Like bZIP Transcription Factors, BrABI5a and BrABI5b, as Positive Modulators of ABA Signalling in Chinese Cabbage. PLoS ONE 2016, 11, e0158966. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.; Manoharan, R.K.; Kang, J.-G.; Chung, M.-Y.; Kim, Y.-W.; Nou, I.-S. Genome-Wide Identification and Characterization of bZIP Transcription Factors in Brassica Oleracea under Cold Stress. Biomed. Res. Int. 2016, 2016, 4376598. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.; Jung, H.-J.; Park, J.-I.; Yang, T.-J.; Nou, I.-S. Transcriptome Analysis of Newly Classified bZIP Transcription Factors of Brassica Rapa in Cold Stress Response. Genomics 2014, 104, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Jakoby, M.; Weisshaar, B.; Dröge-Laser, W.; Vicente-Carbajosa, J.; Tiedemann, J.; Kroj, T.; Parcy, F. bZIP Transcription Factors in Arabidopsis. Trends Plant Sci. 2002, 7, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.-T.; Ma, L.; Duan, W.; Wang, B.-C.; Li, J.-H.; Xu, H.-G.; Yan, X.-Q.; Yan, B.-F.; Li, S.-H.; Wang, L.-J. Differential Proteomic Analysis of Grapevine Leaves by iTRAQ Reveals Responses to Heat Stress and Subsequent Recovery. BMC Plant Biol. 2014, 14, 110. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Xu, F.; Li, Y.; Yu, L.; Fu, M.; Liao, Y.; Yang, X.; Zhang, W.; Ye, J. Genome-Wide Characterization of bZIP Gene Family Identifies Potential Members Involved in Flavonoids Biosynthesis in Ginkgo biloba L. Sci. Rep. 2021, 11, 23420. [Google Scholar] [CrossRef] [PubMed]
- Dröge-Laser, W.; Snoek, B.L.; Snel, B.; Weiste, C. The Arabidopsis bZIP Transcription Factor Family-an Update. Curr. Opin. Plant Biol. 2018, 45, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Chen, S.; Yao, W.; Cheng, Z.; Zhou, B.; Jiang, T. Genome-Wide Analysis and Expression Profile of the bZIP Gene Family in Poplar. BMC Plant Biol. 2021, 21, 122. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Xu, L.; Wang, Y.; Tang, M.; Liu, L. Genome- and Transcriptome-Wide Characterization of bZIP Gene Family Identifies Potential Members Involved in Abiotic Stress Response and Anthocyanin Biosynthesis in Radish (Raphanus sativus L.). Int. J. Mol. Sci. 2019, 20, 6334. [Google Scholar] [CrossRef] [PubMed]
- Rong, S.; Wu, Z.; Cheng, Z.; Zhang, S.; Liu, H.; Huang, Q. Genome-Wide Identification, Evolutionary Patterns, and Expression Analysis of bZIP Gene Family in Olive (Olea europaea L.). Genes 2020, 11, 510. [Google Scholar] [CrossRef]
- Boudet, N.; Aubourg, S.; Toffano-Nioche, C.; Kreis, M.; Lecharny, A. Evolution of Intron/Exon Structure of DEAD Helicase Family Genes in Arabidopsis, Caenorhabditis, and Drosophila. Genome Res. 2001, 11, 2101–2114. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Zheng, Y.; Xin, H.; Fang, L.; Li, S. Comprehensive Analysis of NAC Domain Transcription Factor Gene Family in Vitis Vinifera. Plant Cell Rep. 2013, 32, 61–75. [Google Scholar] [CrossRef]
- Azeem, F.; Tahir, H.; Ijaz, U.; Shaheen, T. A Genome-Wide Comparative Analysis of bZIP Transcription Factors in G. arboreum and G. raimondii (Diploid Ancestors of Present-Day Cotton). Physiol. Mol. Biol. Plants 2020, 26, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Manzoor, M.A.; Manzoor, M.M.; Li, G.; Abdullah, M.; Han, W.; Wenlong, H.; Shakoor, A.; Riaz, M.W.; Rehman, S.; Cai, Y. Genome-Wide Identification and Characterization of bZIP Transcription Factors and Their Expression Profile under Abiotic Stresses in Chinese Pear (Pyrus bretschneideri). BMC Plant Biol. 2021, 21, 413. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Guo, C.; Li, Z.; Sun, J.; Wang, D.; Xu, L.; Li, X.; Guo, Y. Identification and Analysis of bZIP Family Genes in Potato and Their Potential Roles in Stress Responses. Front. Plant Sci. 2021, 12, 637343. [Google Scholar] [CrossRef] [PubMed]
- Rathour, M.; Shumayla; Alok, A.; Upadhyay, S.K. Investigation of Roles of TaTALE Genes during Development and Stress Response in Bread Wheat. Plants 2022, 11, 587. [Google Scholar] [CrossRef] [PubMed]
- Sharma, H.; Sharma, A.; Rajput, R.; Sidhu, S.; Dhillon, H.; Verma, P.C.; Pandey, A.; Upadhyay, S.K. Molecular Characterization, Evolutionary Analysis, and Expression Profiling of BOR Genes in Important Cereals. Plants 2022, 11, 911. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Moreno, A.; López-Domínguez, R.; Villatoro-García, J.A.; Ramirez-Mena, A.; Aparicio-Puerta, E.; Hackenberg, M.; Pascual-Montano, A.; Carmona-Saez, P. Functional Enrichment Analysis of Regulatory Elements. Biomedicines 2022, 10, 590. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Chen, Q.; Dong, H.; Zhang, S.; Huang, X. Genome-Wide Identification and Expression Analysis of the bZIP Transcription Factors, and Functional Analysis in Response to Drought and Cold Stresses in Pear (Pyrus breschneideri). BMC Plant Biol. 2021, 21, 583. [Google Scholar] [CrossRef]
- Choi, J.-W.; Kim, H.-E.; Kim, S. Two Different Domain Architectures Generate Structural and Functional Diversity among bZIP Genes in the Solanaceae Family. Front. Plant Sci. 2022, 13, 967546. [Google Scholar] [CrossRef] [PubMed]
- Bertolotti, A.; Zhang, Y.; Hendershot, L.M.; Harding, H.P.; Ron, D. Dynamic Interaction of BiP and ER Stress Transducers in the Unfolded-Protein Response. Nat. Cell Biol. 2000, 2, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Noh, S.-J.; Kwon, C.S.; Oh, D.-H.; Moon, J.S.; Chung, W.-I. Expression of an Evolutionarily Distinct Novel BiP Gene during the Unfolded Protein Response in Arabidopsis thaliana. Gene 2003, 311, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, R.; Deng, Y.; Howell, S. Stress Sensing in Plants by an ER Stress Sensor/Transducer, bZIP28. Front. Plant Sci. 2014, 5, 59. [Google Scholar] [CrossRef] [PubMed]
- Ghemrawi, R.; Khair, M. Endoplasmic Reticulum Stress and Unfolded Protein Response in Neurodegenerative Diseases. Int. J. Mol. Sci. 2020, 21, 6127. [Google Scholar] [CrossRef] [PubMed]
- Trachootham, D.; Lu, W.; Ogasawara, M.A.; Valle, N.R.-D.; Huang, P. Redox Regulation of Cell Survival. Antioxid. Redox Signal. 2008, 10, 1343–1374. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S/No. | Paralogue Gene Pairs | Ka | Ks | Ka/Ks | Time (MYA) | |
|---|---|---|---|---|---|---|
| 1 | BraC06g036350.1 | BraC06g036450.1 | 0.002779 | 0.009631 | 0.288556 | 00.73 |
| 2 | BraC07g002710.1 | BraC09g010600.1 | 0.042735 | 0.242491 | 0.176234 | 18.48 |
| 3 | BraC03g061850.1 | BraC08g016590.1 | 0.073727 | 0.422454 | 0.174521 | 32.20 |
| 4 | BraC02g023630.1 | BraC07g027810.1 | 0.132831 | 0.455651 | 0.29152 | 34.73 |
| 5 | BraC07g025140.1 | BraC09g059910.1 | 0.069034 | 0.401945 | 0.171749 | 30.64 |
| 6 | BraC01g001260.1 | BraC03g064330.1 | 0.153618 | 0.31279 | 0.491122 | 23.84 |
| 7 | BraC02g047730.1 | BraC02g048410.1 | 0.021165 | 0.039781 | 0.532035 | 3.03 |
| 8 | BraC03g007230.1 | BraC10g025270.1 | 0.090611 | 0.280135 | 0.323456 | 21.35 |
| 9 | BraC02g041500.1 | BraC06g042940.1 | 0.052051 | 0.251405 | 0.207041 | 19.16 |
| 10 | BraC02g038580.1 | BraC06g046020.1 | 0.074368 | 0.354414 | 0.209835 | 27.01 |
| 11 | BraC06g009370.1 | BraC09g068000.1 | 0.072984 | 0.223345 | 0.326779 | 17.02 |
| 12 | BraC02g045000.1 | BraC09g006060.1 | 0.101614 | 0.413633 | 0.245661 | 31.53 |
| 13 | BraC02g042590.1 | BraC06g041080.1 | 0.069065 | 0.319646 | 0.216069 | 24.36 |
| 14 | BraC02g044260.1 | BraC06g038750.1 | 0.089566 | 0.244669 | 0.36607 | 18.65 |
| 15 | BraC04g005820.1 | BraC09g053140.1 | 0.298996 | 0.668932 | 0.446976 | 50.99 |
| 16 | BraC07g022530.1 | BraC09g055620.1 | 0.077406 | 0.337156 | 0.229585 | 25.70 |
| 17 | BraC09g055600.1 | BraC09g055640.1 | 0.001699 | 0.01187 | 0.143158 | 0.90 |
| 18 | BraC04g031300.1 | BraC05g002400.1 | 0.110415 | 0.253654 | 0.435296 | 19.33 |
| 19 | BraC04g027890.1 | BraC05g009380.1 | 0.044586 | 0.338605 | 0.131674 | 25.81 |
| 20 | BraC06g050540.1 | BraC10g002520.1 | 0.308732 | 1.058908 | 0.291557 | 80.71 |
| 21 | BraC03g039400.1 | BraC03g039750.1 | 0.017427 | 0.030102 | 0.578933 | 2.29 |
| 22 | BraC01g039580.1 | BraC05g036280.1 | 0.046399 | 0.22363 | 0.207481 | 17.04 |
| 23 | BraC02g004000.1 | BraC10g028400.1 | 0.045096 | 0.284274 | 0.158634 | 21.67 |
| 24 | BraC09g038170.1 | BraC09g038420.1 | 0.01737 | 0.017292 | 1.004532 | 1.32 |
| 25 | BraC02g003360.1 | BraC10g029390.1 | 0.050306 | 0.424763 | 0.118432 | 32.38 |
| 26 | BraC09g001020.1 | BraC09g007910.1 | 0.172372 | 0.35456 | 0.486157 | 27.02 |
| 27 | BraC07g026920.1 | BraC07g041060.1 | 0.09098 | 0.484017 | 0.187969 | 36.89 |
| 28 | BraC02g002200.1 | BraC10g031480.1 | 0.119591 | 0.952579 | 0.125544 | 72.61 |
| 29 | BraC01g044020.1 | BraC05g041370.1 | 0.024088 | 0.381676 | 0.06311 | 29.09 |
| 30 | BraC06g005390.3 | BraC09g070850.1 | 0.04399 | 0.278365 | 0.158029 | 21.22 |
| 31 | BraC03g000260.1 | BraC03g002970.1 | 0.015199 | 0.01931 | 0.787134 | 1.47 |
| 32 | BraC07g023180.1 | BraC09g056730.1 | 0.055246 | 0.382918 | 0.144277 | 29.19 |
| 33 | BraC04g017540.2 | BraC09g063730.1 | 0.213386 | 0.537255 | 0.397179 | 40.95 |
| 34 | BraC06g051940.3 | BraC08g023460.1 | 0.110492 | 0.554996 | 0.199086 | 42.30 |
| 35 | BraC05g022350.1 | BraC08g007780.1 | 0.063807 | 0.348792 | 0.182937 | 26.58 |
| 36 | BraC08g034790.1 | BraC09g071660.1 | 0.064297 | 0.446193 | 0.144102 | 34.01 |
| 37 | BraC04g030930.1 | BraC05g002220.1 | 0.042663 | 0.557869 | 0.076475 | 42.52 |
| 38 | BraC08g035030.1 | BraC10g004690.1 | 0.069088 | 0.593214 | 0.116463 | 45.21 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ayaz, A.; Jalal, A.; Zhang, X.; Khan, K.A.; Hu, C.; Li, Y.; Hou, X. In-Depth Characterization of bZIP Genes in the Context of Endoplasmic Reticulum (ER) Stress in Brassica campestris ssp. chinensis. Plants 2024, 13, 1160. https://doi.org/10.3390/plants13081160
Ayaz A, Jalal A, Zhang X, Khan KA, Hu C, Li Y, Hou X. In-Depth Characterization of bZIP Genes in the Context of Endoplasmic Reticulum (ER) Stress in Brassica campestris ssp. chinensis. Plants. 2024; 13(8):1160. https://doi.org/10.3390/plants13081160
Chicago/Turabian StyleAyaz, Aliya, Abdul Jalal, Xiaoli Zhang, Khalid Ali Khan, Chunmei Hu, Ying Li, and Xilin Hou. 2024. "In-Depth Characterization of bZIP Genes in the Context of Endoplasmic Reticulum (ER) Stress in Brassica campestris ssp. chinensis" Plants 13, no. 8: 1160. https://doi.org/10.3390/plants13081160