

Alpha-Melanocyte-Stimulating Hormone Maintains Retinal Homeostasis after Ischemia/Reperfusion
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Ischemia-Reperfusion Injury and α-MSH Treatment
2.2. Immunohistochemistry
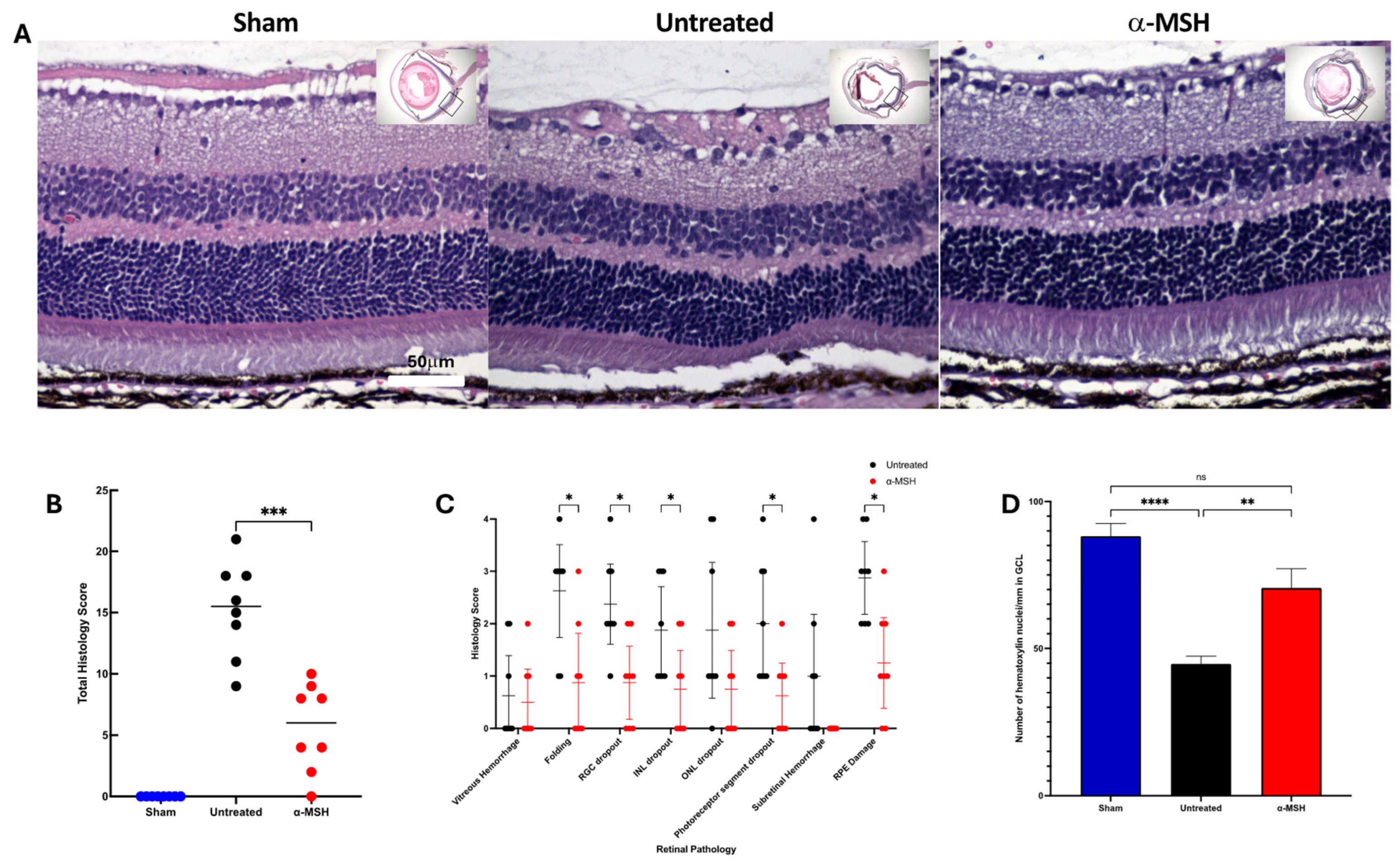
2.3. Histopathological Scoring
2.4. Assay for Apoptosis
2.5. Statistical Analysis
3. Results
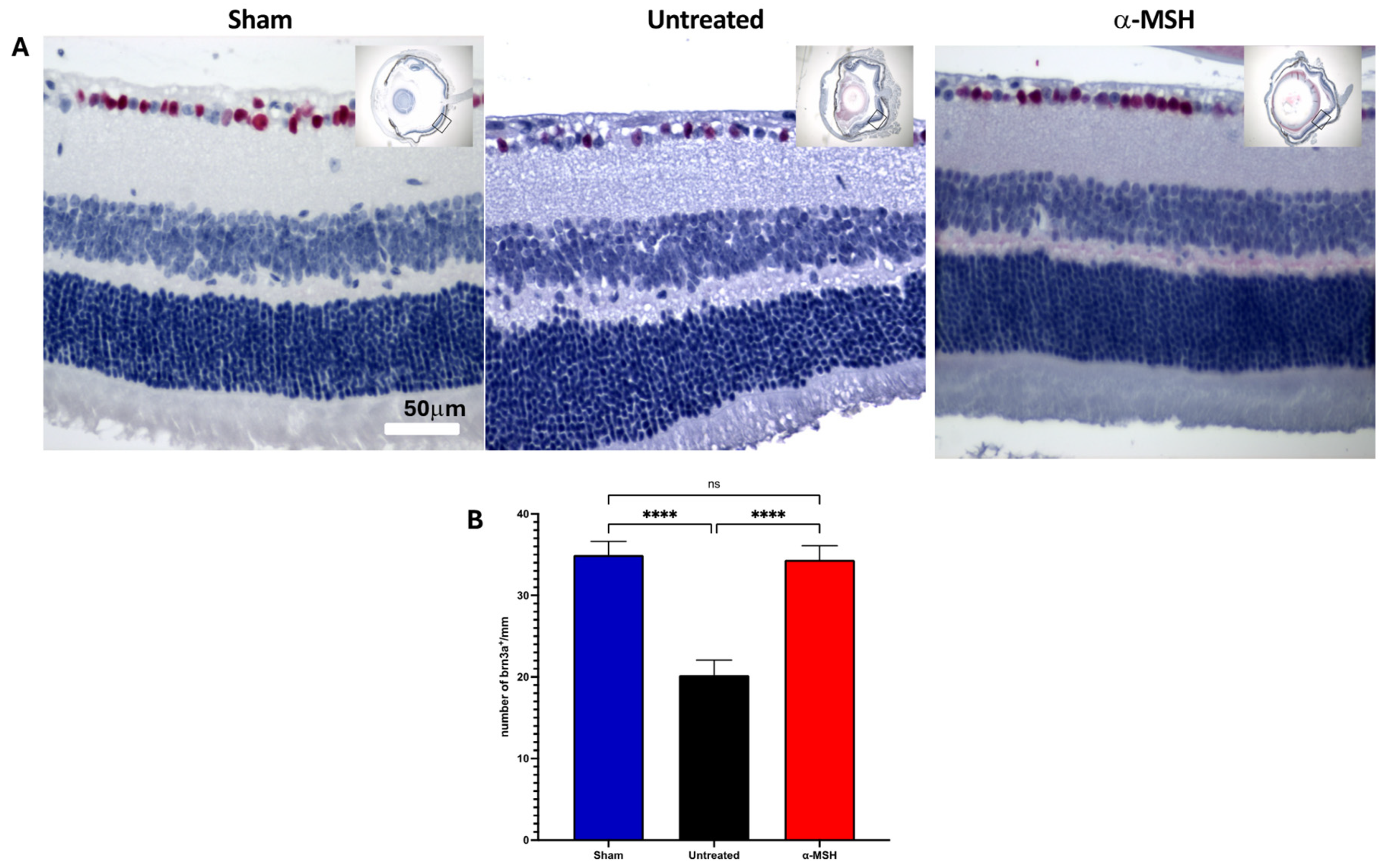
3.1. The Effects of α-MSH Treatment on Retinal Structure and Ganglion Cell Survival following I/R
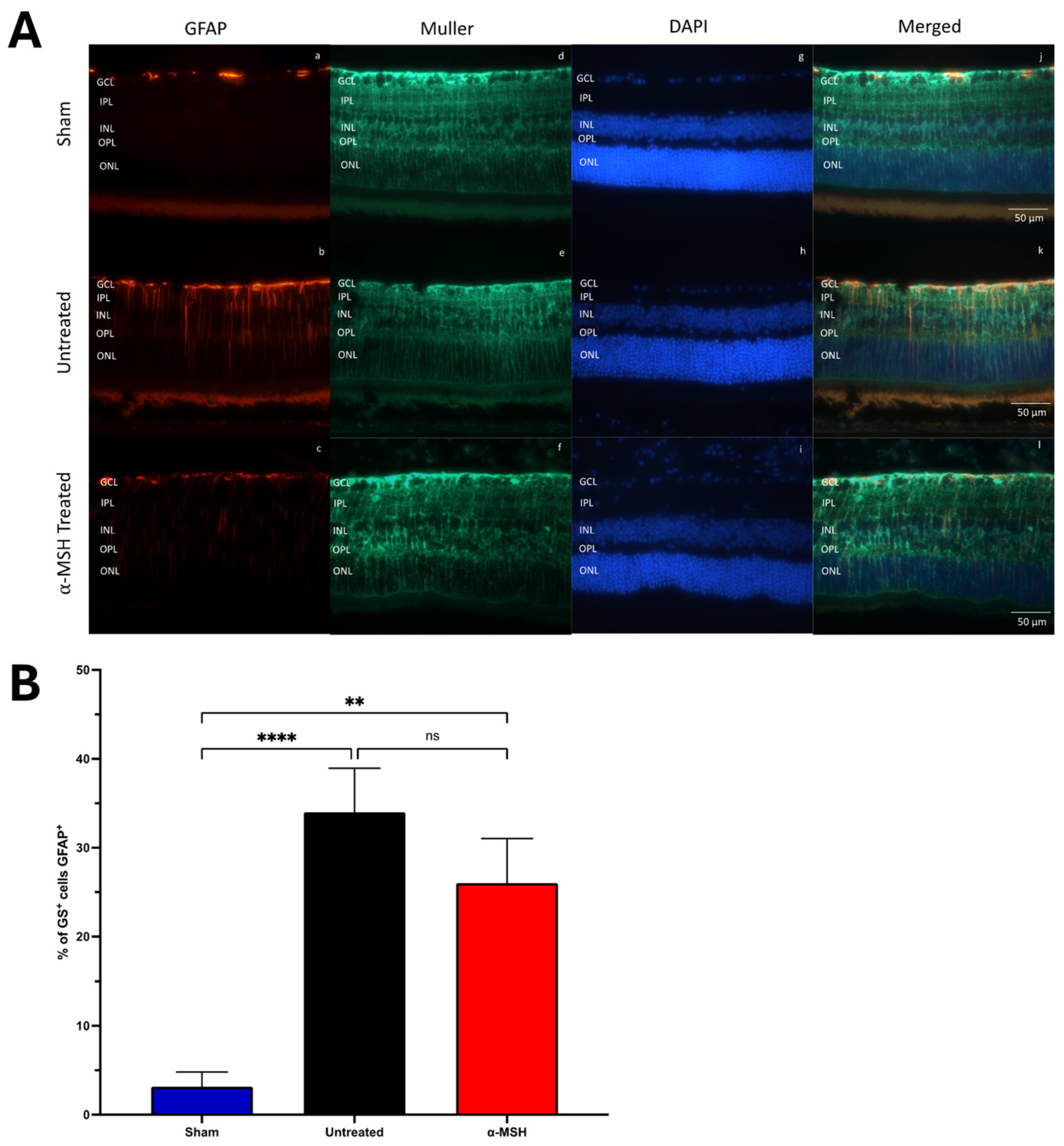
3.2. The Effects of α-MSH Treatment on Muller and Microglial Cell Activation
3.3. The Effects of α-MSH Treatment on Retinal Apoptosis following I/R
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sasaki, C.T.; Vageli, D.P. miR-21, miR-155, miR-192, and miR-375 Deregulations Related to NF-kappaB Activation in Gastroduodenal Fluid-Induced Early Preneoplastic Lesions of Laryngeal Mucosa In Vivo. Neoplasia 2016, 18, 329–338. [Google Scholar] [CrossRef]
- Wong, T.Y.; Cheung, C.M.; Larsen, M.; Sharma, S.; Simo, R. Diabetic retinopathy. Nat. Rev. Dis. Primers 2016, 2, 16012. [Google Scholar] [CrossRef] [PubMed]
- Khanh Vu, T.H.; Chen, H.; Pan, L.; Cho, K.S.; Doesburg, D.; Thee, E.F.; Wu, N.; Arlotti, E.; Jager, M.J.; Chen, D.F. CD4(+) T-Cell Responses Mediate Progressive Neurodegeneration in Experimental Ischemic Retinopathy. Am. J. Pathol. 2020, 190, 1723–1734. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.; Wong, T.Y.; Sabanayagam, C. Epidemiology of diabetic retinopathy, diabetic macular edema and related vision loss. Eye Vis. 2015, 2, 17. [Google Scholar] [CrossRef] [PubMed]
- Strauss, O. Pharmacology of the retinal pigment epithelium, the interface between retina and body system. Eur. J. Pharmacol. 2016, 787, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Kaur, A.; Kumar, R.; Sharma, A. Diabetic Retinopathy Leading to Blindness–A Review. Curr. Diabetes Rev. 2024; online ahead of print. [Google Scholar] [CrossRef]
- Musat, O.; Cernat, C.; Labib, M.; Gheorghe, A.; Toma, O.; Zamfir, M.; Boureanu, A.M. Diabetic Macular Edema. Rom. J. Ophthalmol. 2015, 59, 133–136. [Google Scholar] [PubMed]
- Osborne, N.N.; Casson, R.J.; Wood, J.P.; Chidlow, G.; Graham, M.; Melena, J. Retinal ischemia: Mechanisms of damage and potential therapeutic strategies. Prog. Retin. Eye Res. 2004, 23, 91–147. [Google Scholar] [CrossRef] [PubMed]
- Shahror, R.A.; Morris, C.A.; Mohammed, A.A.; Wild, M.; Zaman, B.; Mitchell, C.D.; Phillips, P.H.; Rusch, N.J.; Shosha, E.; Fouda, A.Y. Role of myeloid cells in ischemic retinopathies: Recent advances and unanswered questions. J. Neuroinflammation 2024, 21, 65. [Google Scholar] [CrossRef]
- Al Zabadi, H.; Taha, I.; Zagha, R. Clinical and Molecular Characteristics of Diabetic Retinopathy and Its Severity Complications among Diabetic Patients: A Multicenter Cross-Sectional Study. J. Clin. Med. 2022, 11, 3945. [Google Scholar] [CrossRef]
- Gantz, I.; Fong, T.M. The melanocortin system. Am. J. Physiol. Endocrinol. Metab. 2003, 284, E468–E474. [Google Scholar] [CrossRef]
- Harno, E.; Gali Ramamoorthy, T.; Coll, A.P.; White, A. POMC: The Physiological Power of Hormone Processing. Physiol. Rev. 2018, 98, 2381–2430. [Google Scholar] [CrossRef] [PubMed]
- Rouault, A.A.J.; Srinivasan, D.K.; Yin, T.C.; Lee, A.A.; Sebag, J.A. Melanocortin Receptor Accessory Proteins (MRAPs): Functions in the melanocortin system and beyond. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 2462–2467. [Google Scholar] [CrossRef] [PubMed]
- Clemson, C.M.; Yost, J.; Taylor, A.W. The Role of Alpha-MSH as a Modulator of Ocular Immunobiology Exemplifies Mechanistic Differences between Melanocortins and Steroids. Ocul. Immunol. Inflamm. 2017, 25, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.W.; Ng, T.F. Negative regulators that mediate ocular immune privilege. J. Leukoc. Biol. 2018, 103, 1179–1187. [Google Scholar] [CrossRef] [PubMed]
- Alemi, H.; Wang, S.; Blanco, T.; Kahale, F.; Singh, R.B.; Ortiz, G.; Musayeva, A.; Yuksel, E.; Pang, K.; Deshpande, N.; et al. The Neuropeptide alpha-Melanocyte-Stimulating Hormone Prevents Persistent Corneal Edema following Injury. Am. J. Pathol. 2024, 194, 150–164. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.B.; Cheng, L.; Bi, H.E.; Zhang, Z.Q.; Yao, J.; Zhou, X.Z.; Jiang, Q. Alpha-melanocyte stimulating hormone protects retinal pigment epithelium cells from oxidative stress through activation of melanocortin 1 receptor-Akt-mTOR signaling. Biochem. Biophys. Res. Commun. 2014, 443, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Jo, S.K.; Cho, W.Y.; Kim, H.K.; Won, N.H. The effect of alpha-melanocyte-stimulating hormone on renal tubular cell apoptosis and tubulointerstitial fibrosis in cyclosporine A nephrotoxicity. Transplantation 2004, 78, 1756–1764. [Google Scholar] [CrossRef]
- Bohm, M.; Wolff, I.; Scholzen, T.E.; Robinson, S.J.; Healy, E.; Luger, T.A.; Schwarz, T.; Schwarz, A. alpha-Melanocyte-stimulating hormone protects from ultraviolet radiation-induced apoptosis and DNA damage. J. Biol. Chem. 2005, 280, 5795–5802. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Kahale, F.; Naderi, A.; Surico, P.L.; Yin, J.; Dohlman, T.; Chen, Y.; Dana, R. Therapeutic Effects of Stimulating the Melanocortin Pathway in Regulating Ocular Inflammation and Cell Death. Biomolecules 2024, 14, 169. [Google Scholar] [CrossRef]
- Cai, S.; Yang, Q.; Hou, M.; Han, Q.; Zhang, H.; Wang, J.; Qi, C.; Bo, Q.; Ru, Y.; Yang, W.; et al. Alpha-Melanocyte-Stimulating Hormone Protects Early Diabetic Retina from Blood-Retinal Barrier Breakdown and Vascular Leakage via MC4R. Cell. Physiol. Biochem. 2018, 45, 505–522. [Google Scholar] [CrossRef]
- Muhammad, F.Y.; Peters, K.; Wang, D.; Lee, D.J. Exacerbation of autoimmune uveitis by obesity occurs through the melanocortin 5 receptor. J. Leukoc. Biol. 2019, 106, 879–887. [Google Scholar] [CrossRef] [PubMed]
- Gesualdo, C.; Balta, C.; Platania, C.B.M.; Trotta, M.C.; Herman, H.; Gharbia, S.; Rosu, M.; Petrillo, F.; Giunta, S.; Della Corte, A.; et al. Fingolimod and Diabetic Retinopathy: A Drug Repurposing Study. Front. Pharmacol. 2021, 12, 718902. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.F.; Taylor, A.W. Stimulating the Melanocortin System in Uveitis and Diabetes Preserves the Structure and Anti-Inflammatory Activity of the Retina. Int. J. Mol. Sci. 2023, 24, 6928. [Google Scholar] [CrossRef] [PubMed]
- Goit, R.K.; Taylor, A.W.; Lo, A.C.Y. Anti-inflammatory alpha-Melanocyte-Stimulating Hormone Protects Retina After Ischemia/Reperfusion Injury in Type I Diabetes. Front. Neurosci. 2022, 16, 799739. [Google Scholar] [CrossRef] [PubMed]
- Hartsock, M.J.; Cho, H.; Wu, L.; Chen, W.J.; Gong, J.; Duh, E.J. A Mouse Model of Retinal Ischemia-Reperfusion Injury Through Elevation of Intraocular Pressure. J. Vis. Exp. 2016, 14, 54065. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Caspi, R.R. Clinical and Functional Evaluation of Ocular Inflammatory Disease Using the Model of Experimental Autoimmune Uveitis. Methods Mol. Biol. 2019, 1899, 211–227. [Google Scholar] [CrossRef]
- Lam, T.T.; Abler, A.S.; Tso, M.O. Apoptosis and caspases after ischemia-reperfusion injury in rat retina. Investig. Ophthalmol. Vis. Sci. 1999, 40, 967–975. [Google Scholar]
- Krishnan, A.; Fei, F.; Jones, A.; Busto, P.; Marshak-Rothstein, A.; Ksander, B.R.; Gregory-Ksander, M. Overexpression of Soluble Fas Ligand following Adeno-Associated Virus Gene Therapy Prevents Retinal Ganglion Cell Death in Chronic and Acute Murine Models of Glaucoma. J. Immunol. 2016, 197, 4626–4638. [Google Scholar] [CrossRef] [PubMed]
- Nadal-Nicolas, F.M.; Galindo-Romero, C.; Lucas-Ruiz, F.; Marsh-Amstrong, N.; Li, W.; Vidal-Sanz, M.; Agudo-Barriuso, M. Pan-retinal ganglion cell markers in mice, rats, and rhesus macaques. Zool. Res. 2023, 44, 226–248. [Google Scholar] [CrossRef]
- Roesch, K.; Jadhav, A.P.; Trimarchi, J.M.; Stadler, M.B.; Roska, B.; Sun, B.B.; Cepko, C.L. The transcriptome of retinal Muller glial cells. J. Comp. Neurol. 2008, 509, 225–238. [Google Scholar] [CrossRef]
- Bringmann, A.; Iandiev, I.; Pannicke, T.; Wurm, A.; Hollborn, M.; Wiedemann, P.; Osborne, N.N.; Reichenbach, A. Cellular signaling and factors involved in Muller cell gliosis: Neuroprotective and detrimental effects. Prog. Retin. Eye Res. 2009, 28, 423–451. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.G.; Lue, L.F. Immune phenotypes of microglia in human neurodegenerative disease: Challenges to detecting microglial polarization in human brains. Alzheimers Res. Ther. 2015, 7, 56. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.J.; Braun, T.A.; Wordinger, R.J.; Clark, A.F. Progressive morphological changes and impaired retinal function associated with temporal regulation of gene expression after retinal ischemia/reperfusion injury in mice. Mol. Neurodegener. 2013, 8, 21. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.W.; Hsu, S.; Ng, T.F. The Role of Retinal Pigment Epithelial Cells in Regulation of Macrophages/Microglial Cells in Retinal Immunobiology. Front. Immunol. 2021, 12, 724601. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.W.; Streilein, J.W.; Cousins, S.W. Identification of alpha-melanocyte stimulating hormone as a potential immunosuppressive factor in aqueous humor. Curr. Eye Res. 1992, 11, 1199–1206. [Google Scholar] [CrossRef] [PubMed]
- Zmijewski, M.A.; Sharma, R.K.; Slominski, A.T. Expression of molecular equivalent of hypothalamic-pituitary-adrenal axis in adult retinal pigment epithelium. J. Endocrinol. 2007, 193, 157–169. [Google Scholar] [CrossRef]
- Naveh, N. Melanocortins applied intravitreally delay retinal dystrophy in Royal College of Surgeons rats. Graefes Arch. Clin. Exp. Ophthalmol. 2003, 241, 1044–1050. [Google Scholar] [CrossRef] [PubMed]
- Teshigawara, K.; Takahashi, S.; Boswell, T.; Li, Q.; Tanaka, S.; Takeuchi, S. Identification of avian alpha-melanocyte-stimulating hormone in the eye: Temporal and spatial regulation of expression in the developing chicken. J. Endocrinol. 2001, 168, 527–537. [Google Scholar] [CrossRef]
- Wu, C.S.; Cioanca, A.V.; Gelmi, M.C.; Wen, L.; Di Girolamo, N.; Zhu, L.; Natoli, R.; Conway, R.M.; Petsoglou, C.; Jager, M.J.; et al. The multifunctional human ocular melanocortin system. Prog. Retin. Eye Res. 2023, 95, 101187. [Google Scholar] [CrossRef]
- Zhang, W.; Wu, Y. Mitogen- and stress-activated protein kinase-1 activation is involved in melanocortin-induced BDNF expression in Neuro2a neuronal cells. Neuroreport 2020, 31, 1007–1014. [Google Scholar] [CrossRef]
- Yu, S.; Doycheva, D.M.; Gamdzyk, M.; Gao, Y.; Guo, Y.; Travis, Z.D.; Tang, J.; Chen, W.X.; Zhang, J.H. BMS-470539 Attenuates Oxidative Stress and Neuronal Apoptosis via MC1R/cAMP/PKA/Nurr1 Signaling Pathway in a Neonatal Hypoxic-Ischemic Rat Model. Oxid. Med. Cell Longev. 2022, 2022, 4054938. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Dong, L.; Liu, X.; Jiang, Y.; Zhang, L.; Zhang, X.; Li, X.; Zhang, Y. Alpha-Melanocyte-stimulating hormone protects retinal vascular endothelial cells from oxidative stress and apoptosis in a rat model of diabetes. PLoS ONE 2014, 9, e93433. [Google Scholar] [CrossRef] [PubMed]
- Edling, A.E.; Gomes, D.; Weeden, T.; Dzuris, J.; Stefano, J.; Pan, C.; Williams, J.; Kaplan, J.; Perricone, M.A. Immunosuppressive activity of a novel peptide analog of alpha-melanocyte stimulating hormone (alpha-MSH) in experimental autoimmune uveitis. J. Neuroimmunol. 2011, 236, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Nishida, T.; Miyata, S.; Itoh, Y.; Mizuki, N.; Ohgami, K.; Shiratori, K.; Ilieva, I.B.; Ohno, S.; Taylor, A.W. Anti-inflammatory effects of alpha-melanocyte-stimulating hormone against rat endotoxin-induced uveitis and the time course of inflammatory agents in aqueous humor. Int. Immunopharmacol. 2004, 4, 1059–1066. [Google Scholar] [CrossRef]
- Shiratori, K.; Ohgami, K.; Ilieva, I.B.; Koyama, Y.; Yoshida, K.; Ohno, S. Inhibition of endotoxin-induced uveitis and potentiation of cyclooxygenase-2 protein expression by alpha-melanocyte-stimulating hormone. Investig. Ophthalmol. Vis. Sci. 2004, 45, 159–164. [Google Scholar] [CrossRef]
- Lipton, J.M.; Catania, A. Anti-inflammatory actions of the neuroimmunomodulator alpha-MSH. Immunol. Today 1997, 18, 140–145. [Google Scholar] [CrossRef]
- Arnason, B.G.; Berkovich, R.; Catania, A.; Lisak, R.P.; Zaidi, M. Mechanisms of action of adrenocorticotropic hormone and other melanocortins relevant to the clinical management of patients with multiple sclerosis. Mult. Scler. 2013, 19, 130–136. [Google Scholar] [CrossRef]
- Bhardwaj, R.S.; Schwarz, A.; Becher, E.; Mahnke, K.; Aragane, Y.; Schwarz, T.; Luger, T.A. Pro-opiomelanocortin-derived peptides induce IL-10 production in human monocytes. J. Immunol. 1996, 156, 2517–2521. [Google Scholar] [CrossRef] [PubMed]
- Mason, M.J.; Van Epps, D. Modulation of IL-1, tumor necrosis factor, and C5a-mediated murine neutrophil migration by alpha-melanocyte-stimulating hormone. J. Immunol. 1989, 142, 1646–1651. [Google Scholar] [CrossRef]
- Catania, A.; Rajora, N.; Capsoni, F.; Minonzio, F.; Star, R.A.; Lipton, J.M. The neuropeptide alpha-MSH has specific receptors on neutrophils and reduces chemotaxis in vitro. Peptides 1996, 17, 675–679. [Google Scholar] [CrossRef]
- Manna, S.K.; Sarkar, A.; Sreenivasan, Y. Alpha-melanocyte-stimulating hormone down-regulates CXC receptors through activation of neutrophil elastase. Eur. J. Immunol. 2006, 36, 754–769. [Google Scholar] [CrossRef]
- Cui, H.S.; Hayasaka, S.; Zhang, X.Y.; Chi, Z.L.; Hayasaka, Y. Effect of alpha-melanocyte-stimulating hormone on interleukin 8 and monocyte chemotactic protein 1 expression in a human retinal pigment epithelial cell line. Ophthalmic Res. 2005, 37, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Maisto, R.; Oltra, M.; Vidal-Gil, L.; Martinez-Gil, N.; Sancho-Pelluz, J.; Filippo, C.D.; Rossi, S.; Amico, M.D.; Barcia, J.M.; Romero, F.J. ARPE-19-derived VEGF-containing exosomes promote neovascularization in HUVEC: The role of the melanocortin receptor 5. Cell Cycle 2019, 18, 413–424. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.W.; Dixit, S.; Yu, J. Retinal Pigment Epithelial Cell Line Suppression of Phagolysosome Activation. Int. J. Ophthalmol. Eye Sci. 2015, (Suppl. S2), 1–6. [Google Scholar]
- Spana, C.; Taylor, A.W.; Yee, D.G.; Makhlina, M.; Yang, W.; Dodd, J. Probing the Role of Melanocortin Type 1 Receptor Agonists in Diverse Immunological Diseases. Front. Pharmacol. 2018, 9, 1535. [Google Scholar] [CrossRef]
- Rossi, S.; Maisto, R.; Gesualdo, C.; Trotta, M.C.; Ferraraccio, F.; Kaneva, M.K.; Getting, S.J.; Surace, E.; Testa, F.; Simonelli, F.; et al. Activation of Melanocortin Receptors MC 1 and MC 5 Attenuates Retinal Damage in Experimental Diabetic Retinopathy. Mediat. Inflamm. 2016, 2016, 7368389. [Google Scholar] [CrossRef]
- Varga, B.; Gesztelyi, R.; Bombicz, M.; Haines, D.; Szabo, A.M.; Kemeny-Beke, A.; Antal, M.; Vecsernyes, M.; Juhasz, B.; Tosaki, A. Protective effect of alpha-melanocyte-stimulating hormone (alpha-MSH) on the recovery of ischemia/reperfusion (I/R)-induced retinal damage in a rat model. J. Mol. Neurosci. 2013, 50, 558–570. [Google Scholar] [CrossRef]
- Patel, H.B.; Montero-Melendez, T.; Greco, K.V.; Perretti, M. Melanocortin receptors as novel effectors of macrophage responses in inflammation. Front. Immunol. 2011, 2, 41. [Google Scholar] [CrossRef] [PubMed]
- Mandrika, I.; Muceniece, R.; Wikberg, J.E. Effects of melanocortin peptides on lipopolysaccharide/interferon-gamma-induced NF-kappaB DNA binding and nitric oxide production in macrophage-like RAW 264.7 cells: Evidence for dual mechanisms of action. Biochem. Pharmacol. 2001, 61, 613–621. [Google Scholar] [CrossRef]
- Cao, W.; Wen, R.; Li, F.; Cheng, T.; Steinberg, R.H. Induction of basic fibroblast growth factor mRNA by basic fibroblast growth factor in Muller cells. Invest. Ophthalmol. Vis. Sci. 1997, 38, 1358–1366. [Google Scholar]
- Harada, T.; Harada, C.; Kohsaka, S.; Wada, E.; Yoshida, K.; Ohno, S.; Mamada, H.; Tanaka, K.; Parada, L.F.; Wada, K. Microglia-Muller glia cell interactions control neurotrophic factor production during light-induced retinal degeneration. J. Neurosci. 2002, 22, 9228–9236. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, M.C.; Chiabrando, G.A. Multitarget Activities of Muller Glial Cells and Low-Density Lipoprotein Receptor-Related Protein 1 in Proliferative Retinopathies. ASN Neuro 2022, 14, 17590914221136365. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Jin, X.; Zhou, P.; Zhu, H.; Xie, K.; Jin, B.; Du, L. Relationship between Uveitis and the Differential Reactivity of Retinal Microglia. Ophthalmic Res. 2023, 66, 1206–1212. [Google Scholar] [CrossRef] [PubMed]
- O’Koren, E.G.; Yu, C.; Klingeborn, M.; Wong, A.Y.W.; Prigge, C.L.; Mathew, R.; Kalnitsky, J.; Msallam, R.A.; Silvin, A.; Kay, J.N.; et al. Microglial Function Is Distinct in Different Anatomical Locations during Retinal Homeostasis and Degeneration. Immunity 2019, 50, 723–737.e727. [Google Scholar] [CrossRef]
- Yu, C.; Lad, E.M.; Mathew, R.; Shiraki, N.; Littleton, S.; Chen, Y.; Hou, J.; Schlepckow, K.; Degan, S.; Chew, L.; et al. Microglia at sites of atrophy restrict the progression of retinal degeneration via galectin-3 and Trem2. J. Exp. Med. 2024, 221, e20231011. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Scores | |
|---|---|---|
| 1 | RPE Damage | 0: None |
| 1: Trace/Mild | ||
| 2: Moderate | ||
| 3: Severe | ||
| 4: Very severe | ||
| 2 | Retinal folds | 0: None |
| 1: One to two folds | ||
| 2: Three to four folds | ||
| 3: Five to six folds | ||
| 4: Seven or more folds | ||
| Cells Loss | ||
| 3 | •Ganglion Cell Layer | 0: <1% |
| 4 | •Inner Nuclear Layer | 1: 1–24% |
| 5 | •Outer Nuclear Layer | 2: 25–49% |
| 6 | •Photoreceptor | 3: 50–74% |
| 4: 75–100% | ||
| Hemorrhage | ||
| 7 | •Vitreous | 0: None |
| 8 | •Subretinal | 1: Trace |
| 2: Mild (<5% area) | ||
| 3: Moderate (6–10% area) | ||
| 4: Severe (>10% area) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ng, T.F.; Cho, J.Y.; Zhao, J.L.; Gardiner, J.R.; Wang, E.S.; Leung, E.; Xu, Z.; Fineman, S.L.; Lituchy, M.; Lo, A.C.; et al. Alpha-Melanocyte-Stimulating Hormone Maintains Retinal Homeostasis after Ischemia/Reperfusion. Biomolecules 2024, 14, 525. https://doi.org/10.3390/biom14050525
Ng TF, Cho JY, Zhao JL, Gardiner JR, Wang ES, Leung E, Xu Z, Fineman SL, Lituchy M, Lo AC, et al. Alpha-Melanocyte-Stimulating Hormone Maintains Retinal Homeostasis after Ischemia/Reperfusion. Biomolecules. 2024; 14(5):525. https://doi.org/10.3390/biom14050525
Chicago/Turabian StyleNg, Tat Fong, Jenna Y. Cho, John L. Zhao, John R. Gardiner, Eric S. Wang, Elman Leung, Ziqian Xu, Samantha L. Fineman, Melinda Lituchy, Amy C. Lo, and et al. 2024. "Alpha-Melanocyte-Stimulating Hormone Maintains Retinal Homeostasis after Ischemia/Reperfusion" Biomolecules 14, no. 5: 525. https://doi.org/10.3390/biom14050525