
Short- and Long-Term Effects of Subchronic Stress Exposure in Male and Female Brain-Derived Neurotrophic Factor Knock-In Val66Met Mice
 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Forced Swimming Stress (FSS)
2.3. Measurement of the Corticosterone Serum Level
2.4. Splash Test
2.5. Novelty-Suppressed Feeding Test
2.6. Sucrose Preference
2.7. RNA Isolation and Reverse Transcription
2.8. Real-Time PCR
2.9. Western Blot Analysis
2.10. Statistical Analysis
3. Results
3.1. The Immobility Time Is Higher in Male BDNFV/M Mice during Exposure to Subchronic FSS
3.2. Subchronic Forced Swimming Stress Induced a Short-Term Anxiety-like Phenotype in Mice
3.3. Subchronic Forced Swimming Stress Induced a Long-Term Anxiety-like Phenotype Only in Male BDNFV/M Mice
3.4. Subchronic Forced Swimming Stress Induced a Long-Term Change of BDNF Exons in the Hippocampus in a Sex- and Genotype-Depending Manner
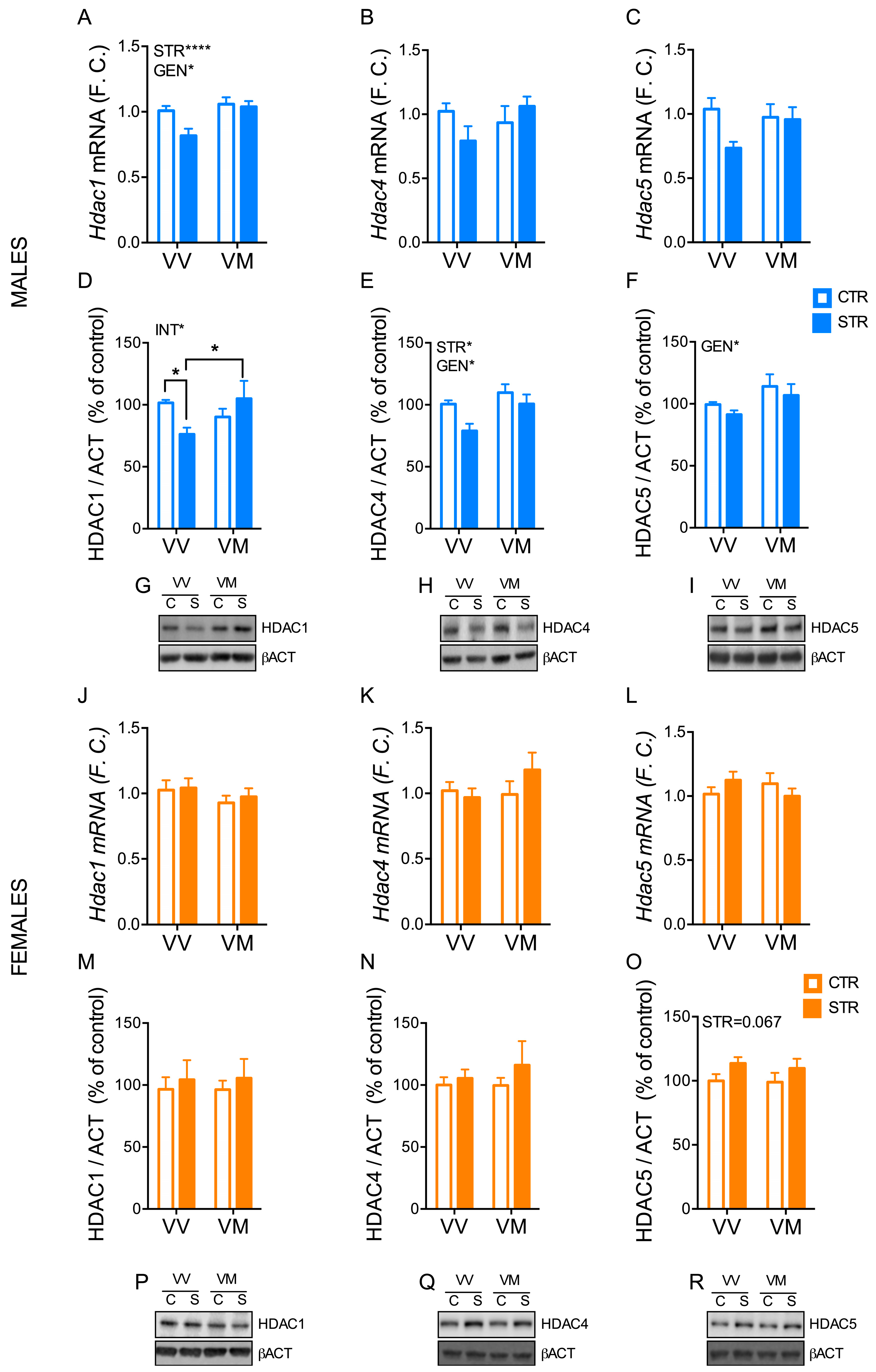
3.5. Subchronic Forced Swimming Stress Induced a Long-Term Change of HDACs in the Hippocampus in a Sex- and Genotype-Depending Manner
3.6. Subchronic Forced Swimming Stress Induced a Long-Term Change of Dnmt in the Hippocampus in a Sex- and Genotype-Depending Manner
3.7. Subchronic Forced Swimming Stress Induced a Long-Term Change of CREB Phosphorylation in the Hippocampus in a Sex- and Genotype-Depending Manner
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Leonardo, E.D.; Hen, R. Anxiety as a developmental disorder. Neuropsychopharmacology 2008, 33, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Kessler, R.C.; Aguilar-Gaxiola, S.; Alonso, J.; Chatterji, S.; Lee, S.; Ormel, J.; Üstün, T.B.; Wang, P.S. The global burden of mental disorders: An update from the WHO World Mental Health (WMH) surveys. Epidemiol. Psichiatr. Soc. 2009, 18, 23–33. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S.; Bowles, N.P.; Gray, J.D.; Hill, M.N.; Hunter, R.G.; Karatsoreos, I.N.; Nasca, C. Mechanisms of stress in the brain. Nat. Neurosci. 2015, 18, 1353–1363. [Google Scholar] [CrossRef]
- Daviu, N.; Bruchas, M.R.; Moghaddam, B.; Sandi, C.; Beyeler, A. Neurobiological links between stress and anxiety. Neurobiol. Stress 2019, 11, 100191. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S.; Eiland, L.; Hunter, R.G.; Miller, M.M. Stress and anxiety: Structural plasticity and epigenetic regulation as a consequence of stress. Neuropharmacology 2012, 62, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Zhao, W.; Zhou, Q.; Yang, Y.; Chen, S.; Hu, J.; Jiang, W.; Zhang, W.; Cai, J.; Qiu, J. Sex differences in depression, anxiety and health-promoting lifestyles among community residents: A network approach. J. Affect. Disord. 2023, 340, 369–378. [Google Scholar] [CrossRef] [PubMed]
- McLean, C.P.; Asnaani, A.; Litz, B.T.; Hofmann, S.G. Gender differences in anxiety disorders: Prevalence, course of illness, comorbidity and burden of illness. J. Psychiatr. Res. 2011, 45, 1027–1035. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Ping, S.; Liu, X. Gender differences in depression, anxiety, and stress among college students: A longitudinal study from China. J. Affect. Disord. 2020, 263, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; He, L.; Ma, R.; Ding, W.; Zhou, C.; Lin, S.; Zhang, J.C.; Chen, J.; Yao, W. The role of MeCP2 and the BDNF/TrkB signaling pathway in the stress resilience of mice subjected to CSDS. Psychopharmacology 2022, 239, 2921–2929. [Google Scholar] [CrossRef]
- Baj, G.; D’Alessandro, V.; Musazzi, L.; Mallei, A.; Sartori, C.R.; Sciancalepore, M.; Tardito, D.; Langone, F.; Popoli, M.; Tongiorgi, E. Physical Exercise and Antidepressants Enhance BDNF Targeting in Hippocampal CA3 Dendrites: Further Evidence of a Spatial Code for BDNF Splice Variants. Neuropsychopharmacology 2012, 37, 1600–1611. [Google Scholar] [CrossRef]
- Bazzari, A.H.; Bazzari, F.H. BDNF Therapeutic Mechanisms in Neuropsychiatric Disorders. Int. J. Mol. Sci. 2022, 23, 8417. [Google Scholar] [CrossRef]
- Notaras, M.; van den Buuse, M. Neurobiology of BDNF in fear memory, sensitivity to stress, and stress-related disorders. Mol. Psychiatry 2020, 25, 2251–2274. [Google Scholar] [CrossRef] [PubMed]
- Egan, M.F.; Kojima, M.; Callicott, J.H.; Goldberg, T.E.; Kolachana, B.S.; Bertolino, A.; Zaitsev, E.; Gold, B.; Goldman, D.; Dean, M.; et al. The BDNF val66met polymorphism affects activity-dependent secretion of BDNF and human memory and hippocampal function. Cell 2003, 112, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Y.; Jing, D.; Bath, K.G.; Ieraci, A.; Khan, T.; Siao, C.J.; Herrera, D.G.; Toth, M.; Yang, C.; McEwen, B.S.; et al. Genetic variant BDNF (Val66Met) polymorphism alters anxiety-related behavior. Science 2006, 314, 140–143. [Google Scholar] [CrossRef] [PubMed]
- Hosang, G.M.; Shiles, C.; Tansey, K.E.; McGuffin, P.; Uher, R. Interaction between stress and the BDNF Val66Met polymorphism in depression: A systematic review and meta-analysis. BMC Med. 2014, 12, 8417. [Google Scholar] [CrossRef] [PubMed]
- Notaras, M.; Hill, R.; van den Buuse, M. The BDNF gene Val66Met polymorphism as a modifier of psychiatric disorder susceptibility: Progress and controversy. Mol. Psychiatry 2015, 20, 916–930. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Wang, D.-D.; Wang, Y.; Liu, T.; Lee, F.S.; Chen, Z.-Y. Variant Brain-Derived Neurotrophic Factor Val66Met Polymorphism Alters Vulnerability to Stress and Response to Antidepressants. J. Neurosci. 2012, 32, 4092–4101. [Google Scholar] [CrossRef] [PubMed]
- Sandrini, L.; Ieraci, A.; Amadio, P.; Veglia, F.; Popoli, M.; Lee, F.S.; Tremoli, E.; Barbieri, S.S. Sub-chronic stress exacerbates the pro-thrombotic phenotype in BDNFVal/Met mice: Gene-environment interaction in the modulation of arterial thrombosis. Int. J. Mol. Sci. 2018, 19, 3235. [Google Scholar] [CrossRef] [PubMed]
- Musazzi, L.; Tornese, P.; Sala, N.; Lee, F.S.; Popoli, M.; Ieraci, A. Acute stress induces an aberrant increase of presynaptic release of glutamate and cellular activation in the hippocampus of BDNFVal/Met mice. J. Cell. Physiol. 2022, 237, 3834–3844. [Google Scholar] [CrossRef]
- Lin, E.; Tsai, S.J. Gene-environment interactions and role of epigenetics in anxiety disorders. In Advances in Experimental Medicine and Biology; Springer: Berlin/Heidelberg, Germany, 2020; Volume 1191, pp. 93–102. [Google Scholar]
- Rudenko, A.; Tsai, L.H. Epigenetic regulation in memory and cognitive disorders. Neuroscience 2014, 264, 51–63. [Google Scholar] [CrossRef]
- Biswas, S.; Rao, C.M. Epigenetic tools (The Writers, The Readers and The Erasers) and their implications in cancer therapy. Eur. J. Pharmacol. 2018, 837, 8–24. [Google Scholar] [CrossRef] [PubMed]
- Hyun, K.; Jeon, J.; Park, K.; Kim, J. Writing, erasing and reading histone lysine methylations. Exp. Mol. Med. 2017, 49, e324. [Google Scholar] [CrossRef] [PubMed]
- Covington, H.E.; Maze, I.; LaPlant, Q.C.; Vialou, V.F.; Ohnishi, Y.N.; Berton, O.; Fass, D.M.; Renthal, W.; Rush, A.J.; Wu, E.Y.; et al. Antidepressant Actions of Histone Deacetylase Inhibitors. J. Neurosci. 2009, 29, 11451–11460. [Google Scholar] [CrossRef] [PubMed]
- Persaud, N.S.; Cates, H.M. The Epigenetics of Anxiety Pathophysiology: A DNA Methylation and Histone Modification Focused Review. eNeuro 2023, 10. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, S.S.; Sandrini, L.; Musazzi, L.; Popoli, M.; Ieraci, A. Apocynin Prevents Anxiety-Like Behavior and Histone Deacetylases Overexpression Induced by Sub-Chronic Stress in Mice. Biomolecules 2021, 11, 885. [Google Scholar] [CrossRef] [PubMed]
- Mallei, A.; Ieraci, A.; Popoli, M. Chronic social defeat stress differentially regulates the expression of BDNF transcripts and epigenetic modifying enzymes in susceptible and resilient mice. World J. Biol. Psychiatry 2019, 20, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Fan, W.; Zhang, X.; Dong, E. Gestational stress induces depressive-like and anxiety-like phenotypes through epigenetic regulation of BDNF expression in offspring hippocampus. Epigenetics 2016, 11, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Sailaja, B.S.; Cohen-Carmon, D.; Zimmerman, G.; Soreq, H.; Meshorer, E. Stress-induced epigenetic transcriptional memory of acetylcholinesterase by HDAC4. Proc. Natl. Acad. Sci. USA 2012, 109, E3687–E3695. [Google Scholar] [CrossRef]
- Wei, J.; Cheng, J.; Waddell, N.J.; Wang, Z.J.; Pang, X.; Cao, Q.; Liu, A.; Chitaman, J.M.; Abreu, K.; Jasrotia, R.S.; et al. DNA Methyltransferase 3A Is Involved in the Sustained Effects of Chronic Stress on Synaptic Functions and Behaviors. Cereb. Cortex 2021, 31, 1998–2012. [Google Scholar] [CrossRef]
- Hodes, G.E.; Pfau, M.L.; Purushothaman, I.; Ahn, H.F.; Golden, S.A.; Christoffel, D.J.; Magida, J.; Brancato, A.; Takahashi, A.; Flanigan, M.E.; et al. Sex Differences in Nucleus Accumbens Transcriptome Profiles Associated with Susceptibility versus Resilience to Subchronic Variable Stress. J. Neurosci. 2015, 35, 16362–16376. [Google Scholar] [CrossRef]
- Ravanelli, F.; Musazzi, L.; Barbieri, S.S.; Rovati, G.; Popoli, M.; Barbon, A.; Ieraci, A. Differential Epigenetic Changes in the Dorsal Hippocampus of Male and Female SAMP8 Mice: A Preliminary Study. Int. J. Mol. Sci. 2023, 24, 13084. [Google Scholar] [CrossRef]
- Overstreet, D.H. Modeling Depression in Animal Models. In Methods in Molecular Biology (Clifton, N.J.); Springer: Berlin/Heidelberg, Germany, 2012; Volume 829, pp. 125–144. [Google Scholar]
- Baugher, B.J.; Buckhaults, K.; Case, J.; Sullivan, A.; Huq, S.N.; Sachs, B.D. Sub-chronic stress induces similar behavioral effects in male and female mice despite sex-specific molecular adaptations in the nucleus accumbens. Behav. Brain Res. 2022, 425, 113811. [Google Scholar] [CrossRef]
- Mitra, R.; Jadhav, S.; McEwen, B.S.; Vyas, A.; Chattarji, S. Stress duration modulates the spatiotemporal patterns of spine formation in the basolateral amygdala. Proc. Natl. Acad. Sci. USA 2005, 102, 9371–9376. [Google Scholar] [CrossRef]
- Mitra, R.; Sapolsky, R.M. Acute corticosterone treatment is sufficient to induce anxiety and amygdaloid dendritic hypertrophy. Proc. Natl. Acad. Sci. USA 2008, 105, 5573–5578. [Google Scholar] [CrossRef] [PubMed]
- Caradonna, S.G.; Paul, M.R.; Marrocco, J. An allostatic epigenetic memory on chromatin footprints after double-hit acute stress. Neurobiol. Stress 2022, 20, 100475. [Google Scholar] [CrossRef]
- Bonifacino, T.; Mingardi, J.; Facchinetti, R.; Sala, N.; Frumento, G.; Ndoj, E.; Valenza, M.; Paoli, C.; Ieraci, A.; Torazza, C.; et al. Changes at glutamate tripartite synapses in the prefrontal cortex of a new animal model of resilience/vulnerability to acute stress. Transl. Psychiatry 2023, 13, 62. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.; Chattarji, S. Sex differences in the delayed impact of acute stress on the amygdala. Neurobiol. Stress 2021, 14, 100292. [Google Scholar] [CrossRef] [PubMed]
- Torrisi, S.A.; Lavanco, G.; Maurel, O.M.; Gulisano, W.; Laudani, S.; Geraci, F.; Grasso, M.; Barbagallo, C.; Caraci, F.; Bucolo, C.; et al. A novel arousal-based individual screening reveals susceptibility and resilience to PTSD-like phenotypes in mice. Neurobiol. Stress 2020, 14, 100286. [Google Scholar] [CrossRef] [PubMed]
- Tsankova, N.M.; Berton, O.; Renthal, W.; Kumar, A.; Neve, R.L.; Nestler, E.J. Sustained hippocampal chromatin regulation in a mouse model of depression and antidepressant action. Nat. Neurosci. 2006, 9, 519–525. [Google Scholar] [CrossRef]
- Palmisano, M.; Pandey, S.C. Epigenetic mechanisms of alcoholism and stress-related disorders. Alcohol 2017, 60, 7–18. [Google Scholar] [CrossRef]
- Li, Y.F.; Huang, Y.; Amsdell, S.L.; Xiao, L.; O’Donnell, J.M.; Zhang, H.T. Antidepressant- and anxiolytic-like effects of the phosphodiesterase-4 inhibitor rolipram on behavior depend on cyclic AMP response element binding protein-mediated neurogenesis in the hippocampus. Neuropsychopharmacology 2009, 34, 2404–2419. [Google Scholar] [CrossRef] [PubMed]
- Das, S.K.; Baitharu, I.; Barhwal, K.; Hota, S.K.; Singh, S.B. Early mood behavioral changes following exposure to monotonous environment during isolation stress is associated with altered hippocampal synaptic plasticity in male rats. Neurosci. Lett. 2016, 612, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.C.; Roy, A.; Zhang, H.; Xu, T. Partial deletion of the cAMP response element-binding protein gene promotes alcohol-drinking behaviors. J. Neurosci. 2004, 24, 5022–5030. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, H.D.; Duman, R.S. Peripheral BDNF produces antidepressant-like effects in cellular and behavioral models. Neuropsychopharmacology 2010, 35, 2378–2391. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Wang, W.; Zhu, Z.; Zhao, M.; Li, H.; Liu, D.; Pan, F. Involvement of brain-derived neurotrophic factor methylation in the prefrontal cortex and hippocampus induced by chronic unpredictable mild stress in male mice. J. Neurochem. 2023, 164, 624–642. [Google Scholar] [CrossRef] [PubMed]
- Sales, A.J.; Joca, S.R.L. Antidepressant administration modulates stress-induced DNA methylation and DNA methyltransferase expression in rat prefrontal cortex and hippocampus. Behav. Brain Res. 2018, 343, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Sales, A.J.; Maciel, I.S.; Suavinha, A.C.D.R.; Joca, S.R.L. Modulation of DNA Methylation and Gene Expression in Rodent Cortical Neuroplasticity Pathways Exerts Rapid Antidepressant-Like Effects. Mol. Neurobiol. 2021, 58, 777–794. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.B.; Ye, K. Sex differences in brain-derived neurotrophic factor signaling and functions. J. Neurosci. Res. 2017, 95, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Bath, K.G.; Chuang, J.; Spencer-Segal, J.L.; Amso, D.; Altemus, M.; McEwen, B.S.; Lee, F.S. Variant brain-derived neurotrophic factor (Valine66Methionine) polymorphism contributes to developmental and estrous stage-specific expression of anxiety-like behavior in female mice. Biol. Psychiatry 2012, 72, 499–504. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Forward | Primer Reverse |
|---|---|---|
| Bdnf | TCGTTCCTTTCGAGTTAGCC | TTGGTAAACGGCACAAAAC |
| Bdnf4 | CAGAGCAGCTGCCTTGATGTTT | CGCCTTCATGCAACCGAAGTAT |
| Bdnf6 | ACAATGTGACTCCACTGCCGG | CGCCTTCATGCAACCGAAGTAT |
| Hdac1 | GAGTTCTGTCAGTTGTCCACGG | TTCAGACTTCTTTGCATGGTGC |
| Hdac4 | CAATCCCACAGTCTCCGTGT | CAGCACCCCACTAAGGTTCA |
| Hdac5 | TGTCACCGCCAGATGTTTTG | TGAGCAGAGCCGAGACACAG |
| Dnmt1 | GGACACAGGTGCCCGCGA | ATGAACCCCAGATGTTGACCA |
| Dnmt3a | AGATCATGTACGTCGGGGAC | CAATCACCAGGTCGAATGGG |
| Rps18 | TGGAGCGAGTGATCACCATCA | CCTCACGCAGCTTGTTGTCTA |
| bAct | GCCAGAGCAGTAATCTCCTTCT | AGTGTGACGTTGACATCCGTA |
| Gapdh | CGTGCCGCCTGGAGAAACC | CGTGCCGCCTGGAGAAACC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xavier, F.A.C.; Barbieri, S.S.; Popoli, M.; Ieraci, A. Short- and Long-Term Effects of Subchronic Stress Exposure in Male and Female Brain-Derived Neurotrophic Factor Knock-In Val66Met Mice. Biology 2024, 13, 303. https://doi.org/10.3390/biology13050303
Xavier FAC, Barbieri SS, Popoli M, Ieraci A. Short- and Long-Term Effects of Subchronic Stress Exposure in Male and Female Brain-Derived Neurotrophic Factor Knock-In Val66Met Mice. Biology. 2024; 13(5):303. https://doi.org/10.3390/biology13050303
Chicago/Turabian StyleXavier, Fernando Antonio Costa, Silvia Stella Barbieri, Maurizio Popoli, and Alessandro Ieraci. 2024. "Short- and Long-Term Effects of Subchronic Stress Exposure in Male and Female Brain-Derived Neurotrophic Factor Knock-In Val66Met Mice" Biology 13, no. 5: 303. https://doi.org/10.3390/biology13050303