Suppression of Contraction Raises Calcium Ion Levels in the Heart of Zebrafish Larvae

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Fish Husbandry and Morpholino Microinjection
2.2. Mounting of Larvae for Fluorescence Microscopy
2.3. Fluorescence Imaging
2.4. Bioluminescence Imaging
2.5. Image Processing and Data Analysis
2.6. Statistics
3. Results
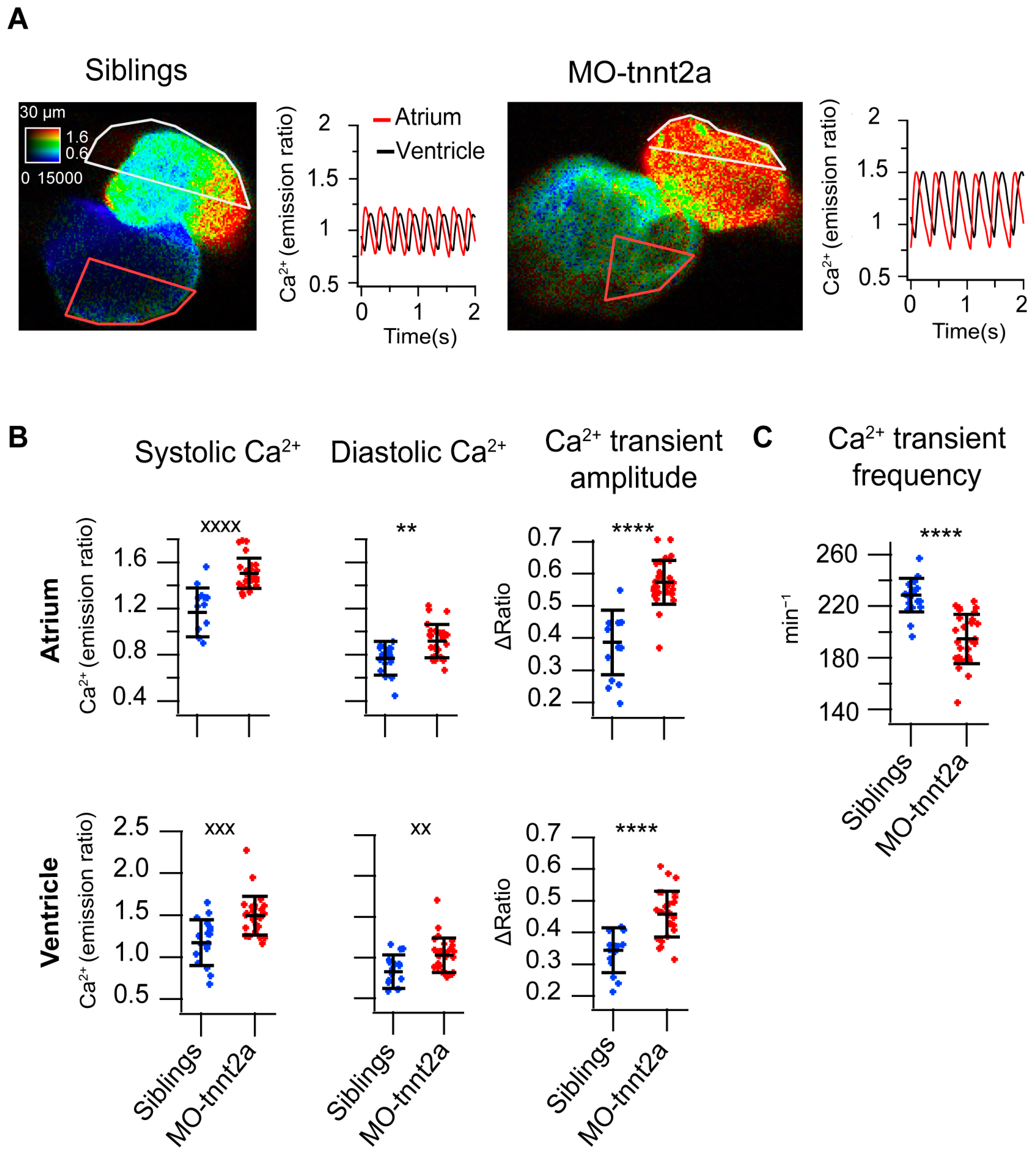
3.1. Morphological and Functional Alterations in tnnt2a Morphant Larvae
3.2. Downregulation of tnnt2a Induced Aberrant Ca2+ Dynamics in 3 dpf Larvae
3.3. Pharmacological Myosin Inhibition with Para-Aminoblebbistatin Alters Ca2+ Dynamics in 3 dpf Larvae
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baillie, J.S.; Stoyek, M.R.; Quinn, T.A. Seeing the Light: The Use of Zebrafish for Optogenetic Studies of the Heart. Front. Physiol. 2021, 12, 748570. [Google Scholar] [CrossRef] [PubMed]
- Sehnert, A.J.; Huq, A.; Weinstein, B.M.; Walker, C.; Fishman, M.; Stainier, D.Y. Cardiac troponin T is essential in sarcomere assembly and cardiac contractility. Nat. Genet. 2002, 31, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Kovács, M.; Tóth, J.; Hetényi, C.; Málnási-Csizmadia, A.; Sellers, J.R. Mechanism of blebbistatin inhibition of myosin II. J. Biol. Chem. 2004, 279, 35557–35563. [Google Scholar] [CrossRef] [PubMed]
- Várkuti, B.H.; Képiró, M.; Horváth, I.; Végner, L.; Ráti, S.; Zsigmond, Á.; Hegyi, G.; Lenkei, Z.; Varga, M.; Málnási-Csizmadia, A. A highly soluble, non-phototoxic, non-fluorescent blebbistatin derivative. Sci. Rep. 2016, 6, 26141. [Google Scholar] [CrossRef] [PubMed]
- Lin, E.; Shafaattalab, S.; Gill, J.; Al-Zeer, B.; Craig, C.; Lamothe, M.; Rayani, K.; Gunawan, M.; Li, A.Y.; Hove-Madsen, L.; et al. Physiological phenotyping of the adult zebrafish heart. Mar. Genom. 2020, 49, 100701. [Google Scholar] [CrossRef] [PubMed]
- van Opbergen, C.J.M.; Koopman, C.D.; Kok, B.J.M.; Knöpfel, T.; Renninger, S.L.; Orger, M.B.; Vos, M.A.; van Veen, T.A.B.; Bakkers, J.; de Boer, T.P. Optogenetic sensors in the zebrafish heart: A novel in vivo electrophysiological tool to study cardiac arrhythmogenesis. Theranostics 2018, 8, 4750–4764. [Google Scholar] [CrossRef] [PubMed]
- Baillie, J.S.; Gendernalik, A.; Garrity, D.M.; Bark, D., Jr.; Quinn, T.A. The in vivo study of cardiac mechano-electric and mechano-mechanical coupling during heart development in zebrafish. Front. Physiol. 2023, 14, 1086050. [Google Scholar] [CrossRef] [PubMed]
- van Opbergen, C.J.M.; van der Voorn, S.M.; Vos, M.A.; de Boer, T.P.; van Veen, T.A.B. Cardiac Ca(2+) signalling in zebrafish: Translation of findings to man. Prog. Biophys. Mol. Biol. 2018, 138, 45–58. [Google Scholar] [CrossRef]
- Salgado-Almario, J.; Vicente, M.; Molina, Y.; Martinez-Sielva, A.; Vincent, P.; Domingo, B.; Llopis, J. Simultaneous imaging of calcium and contraction in the beating heart of zebrafish larvae. Theranostics 2022, 12, 1012–1029. [Google Scholar] [CrossRef]
- Vicente, M.; Salgado-Almario, J.; Valiente-Gabioud, A.A.; Collins, M.M.; Vincent, P.; Domingo, B.; Llopis, J. Early calcium and cardiac contraction defects in a model of phospholamban R9C mutation in zebrafish. J. Mol. Cell Cardiol. 2022, 173, 127–140. [Google Scholar] [CrossRef]
- Salgado-Almario, J.; Vicente, M.; Vincent, P.; Domingo, B.; Llopis, J. Mapping Calcium Dynamics in the Heart of Zebrafish Embryos with Ratiometric Genetically Encoded Calcium Indicators. Int. J. Mol. Sci. 2020, 21, 6610. [Google Scholar] [CrossRef] [PubMed]
- Vicente, M.; Salgado-Almario, J.; Collins, M.M.; Martínez-Sielva, A.; Minoshima, M.; Kikuchi, K.; Domingo, B.; Llopis, J. Cardioluminescence in Transgenic Zebrafish Larvae: A Calcium Imaging Tool to Study Drug Effects and Pathological Modeling. Biomedicines 2021, 9, 1294. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.H.; Cho, M.J.; Kole, R. Up-regulation of luciferase gene expression with antisense oligonucleotides: Implications and applications in functional assay development. Biochemistry 1998, 37, 6235–6239. [Google Scholar] [CrossRef] [PubMed]
- Mota, É.; Bompierre, S.; Betolngar, D.; Castro, L.R.V.; Vincent, P. Pivotal role of phosphodiesterase 10A in the integration of dopamine signals in mice striatal D(1) and D(2) medium-sized spiny neurones. Br. J. Pharmacol. 2021, 178, 4873–4890. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.H.; Kralj, J.M.; Douglass, A.D.; Engert, F.; Cohen, A.E. Simultaneous mapping of membrane voltage and calcium in zebrafish heart in vivo reveals chamber-specific developmental transitions in ionic currents. Front. Physiol. 2014, 5, 344. [Google Scholar] [CrossRef] [PubMed]
- Jou, C.J.; Spitzer, K.W.; Tristani-Firouzi, M. Blebbistatin effectively uncouples the excitation-contraction process in zebrafish embryonic heart. Cell Physiol. Biochem. 2010, 25, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Kappadan, V.; Telele, S.; Uzelac, I.; Fenton, F.; Parlitz, U.; Luther, S.; Christoph, J. High-Resolution Optical Measurement of Cardiac Restitution, Contraction, and Fibrillation Dynamics in Beating vs. Blebbistatin-Uncoupled Isolated Rabbit Hearts. Front. Physiol. 2020, 11, 464. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.; Quintanilla, J.G.; Alfonso-Almazán, J.M.; Galán-Arriola, C.; Yan, P.; Sánchez-González, J.; Pérez-Castellano, N.; Pérez-Villacastín, J.; Ibañez, B.; Loew, L.M.; et al. In vivo ratiometric optical mapping enables high-resolution cardiac electrophysiology in pig models. Cardiovasc. Res. 2019, 115, 1659–1671. [Google Scholar] [CrossRef]
- Thestrup, T.; Litzlbauer, J.; Bartholomäus, I.; Mues, M.; Russo, L.; Dana, H.; Kovalchuk, Y.; Liang, Y.; Kalamakis, G.; Laukat, Y.; et al. Optimized ratiometric calcium sensors for functional in vivo imaging of neurons and T lymphocytes. Nat. Methods 2014, 11, 175–182. [Google Scholar] [CrossRef]
- Baubet, V.; Le Mouellic, H.; Campbell, A.K.; Lucas-Meunier, E.; Fossier, P.; Brúlet, P. Chimeric green fluorescent protein-aequorin as bioluminescent Ca2+ reporters at the single-cell level. Proc. Natl. Acad. Sci. USA 2000, 97, 7260–7265. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.W.; Wardill, T.J.; Sun, Y.; Pulver, S.R.; Renninger, S.L.; Baohan, A.; Schreiter, E.R.; Kerr, R.A.; Orger, M.B.; Jayaraman, V.; et al. Ultrasensitive fluorescent proteins for imaging neuronal activity. Nature 2013, 499, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Laviv, T.; Kim, B.B.; Chu, J.; Lam, A.J.; Lin, M.Z.; Yasuda, R. Simultaneous dual-color fluorescence lifetime imaging with novel red-shifted fluorescent proteins. Nat. Methods 2016, 13, 989–992. [Google Scholar] [CrossRef] [PubMed]
- Umemoto, N.; Nishimura, Y.; Shimada, Y.; Yamanaka, Y.; Kishi, S.; Ito, S.; Okamori, K.; Nakamura, Y.; Kuroyanagi, J.; Zhang, Z.; et al. Fluorescent-based methods for gene knockdown and functional cardiac imaging in zebrafish. Mol. Biotechnol. 2013, 55, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Quinn, T.A.; Kohl, P. Cardiac Mechano-Electric Coupling: Acute Effects of Mechanical Stimulation on Heart Rate and Rhythm. Physiol. Rev. 2021, 101, 37–92. [Google Scholar] [CrossRef]
- Quinn, T.A.; Kohl, P. The Bainbridge effect: Stretching our understanding of cardiac pacemaking for more than a century. J. Physiol. 2022, 600, 4377–4379. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, E.A.; Stoyek, M.R.; Rose, R.A.; Quinn, T.A. Intrinsic regulation of sinoatrial node function and the zebrafish as a model of stretch effects on pacemaking. Prog. Biophys. Mol. Biol. 2017, 130, 198–211. [Google Scholar] [CrossRef]
- Armelin, V.A.; Braga, V.; Teixeira, M.T.; Guagnoni, I.N.; Wang, T.; Florindo, L.H. The baroreflex in aquatic and amphibious teleosts: Does terrestriality represent a significant driving force for the evolution of a more effective baroreflex in vertebrates? Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2021, 255, 110916. [Google Scholar] [CrossRef]
- Eisner, D.A. Ups and downs of calcium in the heart. J. Physiol. 2018, 596, 19–30. [Google Scholar] [CrossRef]
- Eisner, D.A.; Caldwell, J.L.; Trafford, A.W.; Hutchings, D.C. The Control of Diastolic Calcium in the Heart: Basic Mechanisms and Functional Implications. Circ. Res. 2020, 126, 395–412. [Google Scholar] [CrossRef]
- Bartman, T.; Walsh, E.C.; Wen, K.K.; McKane, M.; Ren, J.; Alexander, J.; Rubenstein, P.A.; Stainier, D.Y. Early myocardial function affects endocardial cushion development in zebrafish. PLoS Biol. 2004, 2, E129. [Google Scholar] [CrossRef] [PubMed]
- Samsa, L.A.; Givens, C.; Tzima, E.; Stainier, D.Y.; Qian, L.; Liu, J. Cardiac contraction activates endocardial Notch signaling to modulate chamber maturation in zebrafish. Development 2015, 142, 4080–4091. [Google Scholar] [CrossRef] [PubMed]
- Salgado-Almario, J.; Molina, Y.; Vicente, M.; Martínez-Sielva, A.; Rodríguez-García, R.; Vincent, P.; Domingo, B.; Llopis, J. ERG potassium channels and T-type calcium channels contribute to the pacemaker and atrioventricular conduction in zebrafish larvae. Acta Physiol. 2024, 240, e14075. [Google Scholar] [CrossRef]
- Garrott, K.; Kuzmiak-Glancy, S.; Wengrowski, A.; Zhang, H.; Rogers, J.; Kay, M.W. K(ATP) channel inhibition blunts electromechanical decline during hypoxia in left ventricular working rabbit hearts. J. Physiol. 2017, 595, 3799–3813. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martinez-Sielva, A.; Vicente, M.; Salgado-Almario, J.; Garcia-Blazquez, A.; Domingo, B.; Llopis, J. Suppression of Contraction Raises Calcium Ion Levels in the Heart of Zebrafish Larvae. Biosensors 2024, 14, 219. https://doi.org/10.3390/bios14050219
Martinez-Sielva A, Vicente M, Salgado-Almario J, Garcia-Blazquez A, Domingo B, Llopis J. Suppression of Contraction Raises Calcium Ion Levels in the Heart of Zebrafish Larvae. Biosensors. 2024; 14(5):219. https://doi.org/10.3390/bios14050219
Chicago/Turabian StyleMartinez-Sielva, Antonio, Manuel Vicente, Jussep Salgado-Almario, Aarón Garcia-Blazquez, Beatriz Domingo, and Juan Llopis. 2024. "Suppression of Contraction Raises Calcium Ion Levels in the Heart of Zebrafish Larvae" Biosensors 14, no. 5: 219. https://doi.org/10.3390/bios14050219