

Lemon Peel Water Extract: A Novel Material for Retinal Health, Protecting Retinal Pigment Epithelial Cells against Dynamin-Related Protein 1-Mediated Mitochondrial Fission by Blocking ROS-Stimulated Mitogen-Activated Protein Kinase/Extracellular Signal-Regulated Kinase Pathway
 , , and
, , and
Abstract
:1. Introduction
2. Material and Methods
2.1. Chemicals
2.2. Preparation of LUWE
2.3. Trolox Equivalent Antioxidant Capacity Assay (TEAC)
2.4. DPPH Radical Scavenging Activity
2.5. Analysis of Flavonoid Content
2.6. Analysis of Polyphenol Content
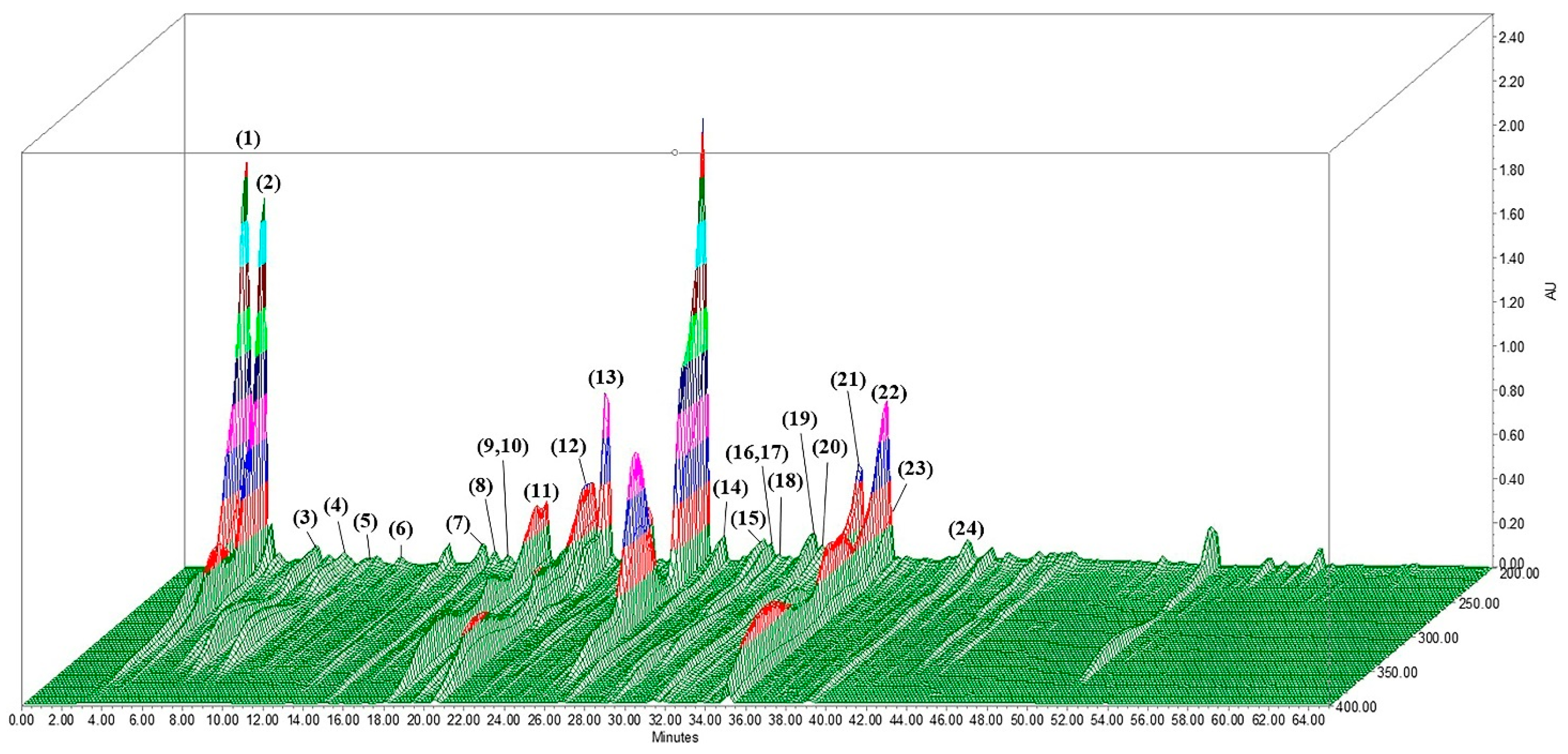
2.7. Analysis of LUWE Components
2.8. Animal Model
2.9. Tissue Sections and Thickness Measurement
2.10. Human RPE Cell Model
2.11. Cell Viability Assay
2.12. Cell Morphology and Apoptosis Analysis
2.13. Mitochondrial ROS Level Analysis
2.14. Western Blotting
2.15. Mitochondrial Dimension Measurements
2.16. Statistical Analysis
3. Results
3.1. Analysis of Antioxidant Activity in LUWE
3.2. LUWE Mitigates Retinal Degeneration in Morphology and Thickness Induced by NaIO3
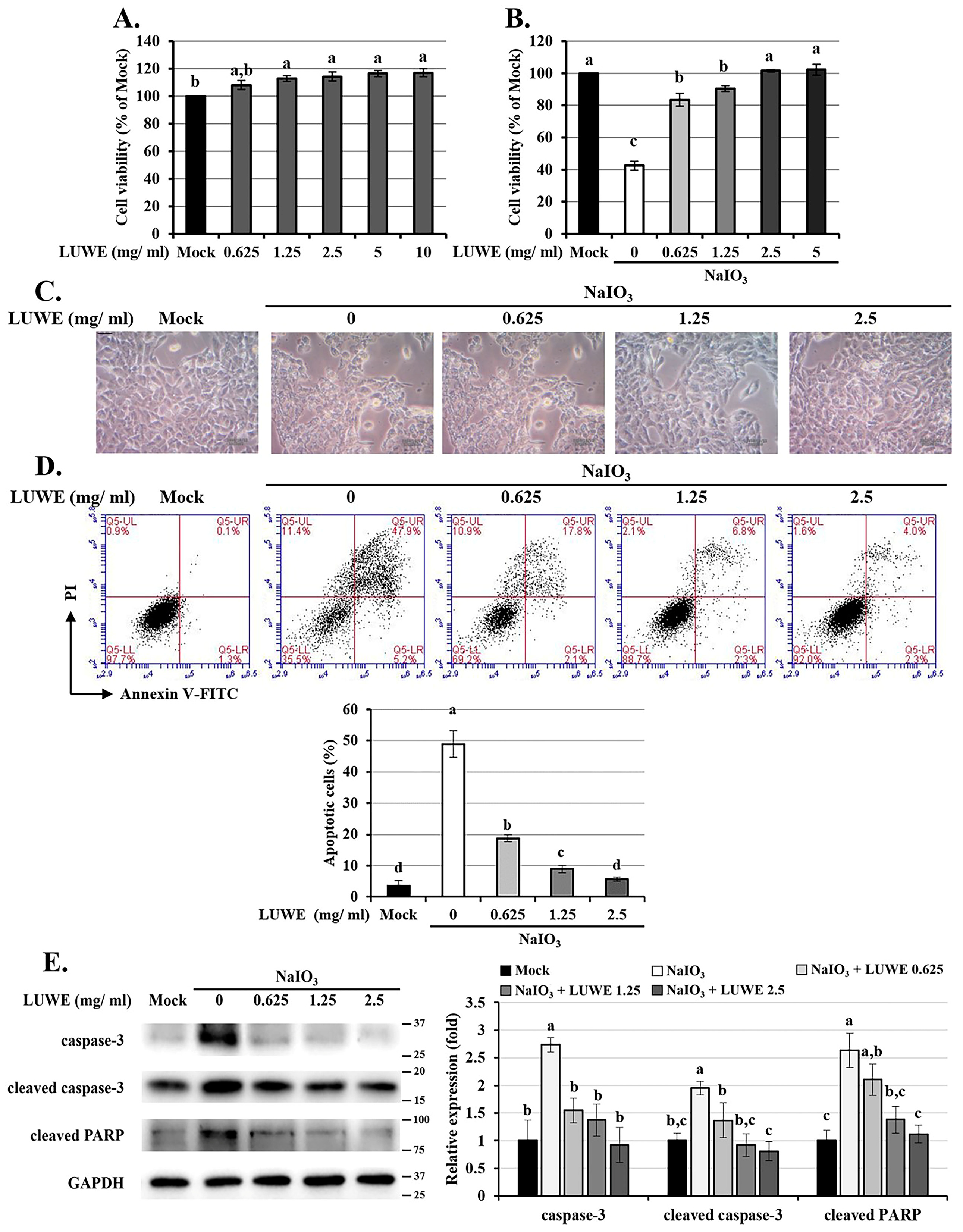
3.3. LUWE Attenuates Cell Death in NaIO3-Treated ARPE-19 Cells
3.4. LUWE Attenuates the Mitochondrial ROS-Stimulated MEK/ERK Signaling Pathway in NaIO3-Treated ARPE-19 Cells
3.5. LUWE Inhibits the Intrinsic Apoptosis Induced by NaIO3 through the MEK/ERK Signaling Pathway
3.6. LUWE Reduces Mitochondrial Fission Regulated by the MEK/ERK Signaling Pathway in NaIO3-Treated ARPE-19 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guymer, R.H.; Campbell, T.G. Age-related macular degeneration. Lancet 2023, 10386, 1459–1472. [Google Scholar] [CrossRef]
- Khachigian, L.M.; Liew, G.; Teo, K.Y.; Wong, T.Y.; Mitchell, P. Emerging therapeutic strategies for unmet need in neovascular age-related macular degeneration. J. Transl. Med. 2023, 21, 133. [Google Scholar] [CrossRef]
- Eshaq, R.S.; Wright, W.S.; Harris, N.R. Oxygen delivery, consumption, and conversion to reactive oxygen species in experimental models of diabetic retinopathy. Redox Biol. 2014, 2, 661–666. [Google Scholar] [CrossRef]
- Chan, C.-M.; Huang, D.-Y.; Sekar, P.; Hsu, S.-H.; Lin, W.-W. Reactive oxygen species-dependent mitochondrial dynamics and autophagy confer protective effects in retinal pigment epithelial cells against sodium iodate-induced cell death. J. Biomed. Sci. 2019, 26, 40. [Google Scholar] [CrossRef]
- Park, C.; Cha, H.-J.; Kim, M.Y.; Bang, E.; Moon, S.-K.; Yun, S.J.; Kim, W.-J.; Noh, J.S.; Kim, G.-Y.; Cho, S.; et al. Phloroglucinol Attenuates DNA Damage and Apoptosis Induced by Oxidative Stress in Human Retinal Pigment Epithelium ARPE-19 Cells by Blocking the Production of Mitochondrial ROS. Antioxidants 2022, 11, 2353. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-Y.; Zhao, Y.; Kim, H.-L.; Oh, Y.; Xu, Q. Sodium iodate-induced retina degeneration observed in non-separate sclerochoroid/retina pigment epithelium/retina whole mounts. Ann. Eye Sci. 2022, 3, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Balmer, J.; Zulliger, R.; Roberti, S.; Enzmann, V. Retinal cell death caused by sodium iodate involves multiple caspase-dependent and caspase-independent cell-death pathways. Int. J. Mol. Sci. 2015, 7, 15086–15103. [Google Scholar] [CrossRef]
- Enzbrenner, A.; Zulliger, R.; Biber, J.; Pousa, A.M.Q.; Schäfer, N.; Stucki, C.; Giroud, N.; Berrera, M.; Kortvely, E.; Schmucki, R.; et al. Sodium iodate-induced degeneration results in local complement changes and inflammatory processes in murine retina. Int. J. Mol. Sci. 2021, 22, 9218. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Wang, T.; Sun, G.-F.; Xiao, J.-X.; Jiang, L.-P.; Tou, F.-F.; Qu, X.-H.; Han, X.-J. Metformin protects against retinal ischemia/reperfusion injury through AMPK-mediated mitochondrial fusion. Free Radic. Biol. Med. 2023, 205, 47–61. [Google Scholar] [CrossRef]
- Zhang, S.-M.; Fan, B.; Li, Y.-L.; Zuo, Z.-Y.; Li, G.-Y. Oxidative Stress-Involved Mitophagy of Retinal Pigment Epithelium and Retinal Degenerative Diseases. Cell. Mol. Neurobiol. 2023, 43, 3265–3276. [Google Scholar] [CrossRef]
- McCubrey, J.A.; LaHair, M.M.; Franklin, R.A. Reactive oxygen species-induced activation of the MAP kinase signaling pathways. Antioxid. Redox Signal. 2006, 8, 1775–1789. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-C.; Chien, Y.; Yarmishyn, A.A.; Lim, L.-Y.; Tsai, H.-Y.; Kuo, W.-C.; Tsai, P.-H.; Yang, S.-H.; Hong, S.-I.; Chen, S.-J.; et al. Inhibition of oxidative stress-induced epithelial-mesenchymal transition in retinal pigment epithelial cells of age-related macular degeneration model by suppressing ERK activation. J. Adv. Res. 2023, 23, 00167-4. [Google Scholar] [CrossRef] [PubMed]
- Flaxel, C.J.; Adelman, R.A.; Bailey, S.T.; Fawzi, A.; Lim, J.I.; Vemulakonda, G.A.; Ying, G.-s. Age-related macular degeneration preferred practice pattern®. Ophthalmology 2020, 127, P1–P65. [Google Scholar] [CrossRef] [PubMed]
- Gardner, T.W.; Antonetti, D.A.; Barber, A.J.; Lieth, E.; Tarbell, J.A.; The Penn State Retina Research Group. The molecular structure and function of the inner blood-retinal barrier. Doc. Ophthalmol. 1999, 97, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Zema, D.; Calabrò, P.; Folino, A.; Tamburino, V.; Zappia, G.; Zimbone, S. Valorisation of citrus processing waste: A review. Waste Manag. 2018, 80, 252–273. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.-S.; Kim, I.-d.; Dhungana, S.K.; Park, E.-J.; Park, J.-J.; Kim, J.-H.; Shin, D.-H. Quality Characteristics and Antioxidant Potential of Lemon (Citrus limon Burm. f.) seed oil extracted by different methods. Front. Nutr. 2021, 8, 644406. [Google Scholar] [CrossRef]
- Saini, R.K.; Ranjit, A.; Sharma, K.; Prasad, P.; Shang, X.; Gowda, K.G.M.; Keum, Y.-S. Bioactive compounds of citrus fruits: A review of composition and health benefits of carotenoids, flavonoids, limonoids, and terpenes. Antioxidants 2022, 11, 239. [Google Scholar] [CrossRef]
- Youdim, K.A.; Qaiser, M.Z.; Begley, D.J.; Rice-Evans, C.A.; Abbott, N.J. Flavonoid permeability across an in situ model of the blood–brain barrier. Free Radic. Biol. Med. 2004, 36, 592–604. [Google Scholar] [CrossRef]
- Muhammad, T.; Ikram, M.; Ullah, R.; Rehman, S.U.; Kim, M.O. Hesperetin, a citrus flavonoid, attenuates LPS-induced neuroinflammation, apoptosis and memory impairments by modulating TLR4/NF-κB signaling. Nutrients 2019, 11, 648. [Google Scholar] [CrossRef]
- Chang, Y.-Y.; Lee, Y.-J.; Hsu, M.-Y.; Wang, M.; Tsou, S.-C.; Chen, C.-C.; Lin, J.-A.; Hsiao, Y.-P.; Lin, H.-W. Protective effect of quercetin on sodium iodate-induced retinal apoptosis through the reactive oxygen species-mediated mitochondrion-dependent pathway. Int. J. Mol. Sci. 2021, 22, 4056. [Google Scholar] [CrossRef]
- Tungmunnithum, D.; Thongboonyou, A.; Pholboon, A.; Yangsabai, A. Flavonoids and other phenolic compounds from medicinal plants for pharmaceutical and medical aspects: An overview. Medicines 2018, 5, 93. [Google Scholar] [CrossRef]
- Costa, R.; Costa Lima, S.A.; Gameiro, P.; Reis, S. On the development of a cutaneous flavonoid delivery system: Advances and limitations. Antioxidants 2021, 10, 1376. [Google Scholar] [CrossRef] [PubMed]
- Song, M.-H.; Shin, E.-C.; Hwang, D.-Y.; Jang, I.-S. Effects of lutein or lutein in combination with vitamin C on mRNA expression and activity of antioxidant enzymes and status of the antioxidant system in SD rats. Lab. Anim. Res. 2015, 31, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Karbuz, P.; Tugrul, N. Microwave and ultrasound assisted extraction of pectin from various fruits peel. J. Food Sci. Technol. 2021, 58, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Papoutsis, K.; Pristijono, P.; Golding, J.B.; Stathopoulos, C.E.; Bowyer, M.C.; Scarlett, C.J.; Vuong, Q.V. Optimizing a sustainable ultrasound-assisted extraction method for the recovery of polyphenols from lemon by-products: Comparison with hot water and organic solvent extractions. Eur. Food Res. Technol. 2018, 244, 1353–1365. [Google Scholar] [CrossRef]
- Jagannath, A.; Biradar, R. Comparative evaluation of soxhlet and ultrasonics on the structural morphology and extraction of bioactive compounds of lemon (Citrus limon L.) peel. J. Food Chem. Nanotechnol. 2019, 5, 56–64. [Google Scholar] [CrossRef]
- Konan, K.; Le Tien, C.; Mateescu, M. Electrolysis-induced fast activation of the ABTS reagent for an antioxidant capacity assay. Anal. Methods. 2016, 8, 5638–5644. [Google Scholar] [CrossRef]
- Anggraini, T.; Nanda, R.F.; Syukri, D. Effect of processing on green and black tea DPPH radical scavenging activity, IC50 value, total polyphenols, catechin and epigallocatechin gallate content. In Proceedings of the 7th International Conference on Sustainable Agriculture, Food and Energy, Phuket, Thailand, 18–21 October 2019; IOP Conference Series: Earth and Environmental Science. IOP Publishing: Bristol, UK, 2021; p. 012017. [Google Scholar]
- Shraim, A.M.; Ahmed, T.A.; Rahman, M.M.; Hijji, Y.M. Determination of total flavonoid content by aluminum chloride assay: A critical evaluation. LWT-Food Sci. Technol. 2021, 150, 111932. [Google Scholar] [CrossRef]
- Carmona-Hernandez, J.C.; Taborda-Ocampo, G.; González-Correa, C.H. Folin-Ciocalteu reaction alternatives for higher polyphenol quantitation in Colombian passion fruits. Int. J. Food Sci. 2021, 2021, 8871301. [Google Scholar] [CrossRef]
- Hsu, M.-Y.; Hsiao, Y.-P.; Lin, Y.-T.; Chen, C.; Lee, C.-M.; Liao, W.-C.; Tsou, S.-C.; Lin, H.-W.; Chang, Y.-Y. Quercetin alleviates the accumulation of superoxide in sodium iodate-induced retinal autophagy by regulating mitochondrial reactive oxygen species homeostasis through enhanced deacetyl-SOD2 via the Nrf2-PGC-1α-Sirt1 pathway. Antioxidants 2021, 10, 1125. [Google Scholar] [CrossRef]
- Tzur, A.; Moore, J.K.; Jorgensen, P.; Shapiro, H.M.; Kirschner, M.W. Optimizing optical flow cytometry for cell volume-based sorting and analysis. PLoS ONE 2011, 1, e16053. [Google Scholar] [CrossRef] [PubMed]
- Stern, A.D.; Rahman, A.H.; Birtwistle, M.R. Cell size assays for mass cytometry. Cytom. Part A 2017, 91, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Kushwah, N.; Bora, K.; Maurya, M.; Pavlovich, M.C.; Chen, J. Oxidative stress and antioxidants in age-related macular degeneration. Antioxidants 2023, 7, 1379. [Google Scholar] [CrossRef] [PubMed]
- Ledesma-Escobar, C.; Priego-Capote, F.; Luque de Castro, M. Characterization of lemon (Citrus limon) polar extract by liquid chromatography–tandem mass spectrometry in high resolution mode. J. Mass Spectrom. 2015, 50, 1196–1205. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, M.A.; Al-Gendy, A.A.; Hamdan, D.I.; El-Shazly, A.M. Phytoconstituents, LC-ESI-MS profile, antioxidant and antimicrobial activities of Citrus x limon L. Burm. f. Cultivar Variegated Pink Lemon. Int. J. Pharm. Sci. 2017, 9, 375. [Google Scholar]
- Wang, Z.; Jablonski, J.E. Targeted and non-targeted detection of lemon juice adulteration by LC-MS and chemometrics. Food Addit. Contam. Part A 2016, 33, 560–573. [Google Scholar] [CrossRef] [PubMed]
- Gargouri, B.; Ammar, S.; Verardo, V.; Besbes, S.; Segura-Carretero, A.; Bouaziz, M. RP-HPLC–DAD-ESI-TOF–MS based strategy for new insights into the qualitative and quantitative phenolic profile in Tunisian industrial Citrus Limon by-product and their antioxidant activity. Eur. Food Res. Technol. 2017, 243, 2011–2024. [Google Scholar] [CrossRef]
- Brito, A.; Ramirez, J.E.; Areche, C.; Sepúlveda, B.; Simirgiotis, M.J. HPLC-UV-MS profiles of phenolic compounds and antioxidant activity of fruits from three citrus species consumed in Northern Chile. Molecules 2014, 19, 17400–17421. [Google Scholar] [CrossRef] [PubMed]
- García-Salas, P.; Gómez-Caravaca, A.M.; Arráez-Román, D.; Segura-Carretero, A.; Guerra-Hernández, E.; García-Villanova, B.; Fernández-Gutiérrez, A. Influence of technological processes on phenolic compounds, organic acids, furanic derivatives, and antioxidant activity of whole-lemon powder. Food Chem. 2013, 141, 869–878. [Google Scholar] [CrossRef]
- Guccione, C.; Bergonzi, M.C.; Piazzini, V.; Bilia, A.R. A simple and rapid HPLC-PDA MS method for the profiling of citrus peels and traditional Italian liquors. Planta Med. 2016, 82, 1039–1045. [Google Scholar] [CrossRef]
- Sanches, V.L.; Cunha, T.A.; Viganó, J.; de Souza Mesquita, L.M.; Faccioli, L.H.; Breitkreitz, M.C.; Rostagno, M.A. Comprehensive analysis of phenolics compounds in citrus fruits peels by UPLC-PDA and UPLC-Q/TOF MS using a fused-core column. Food Chem. X 2022, 14, 100262. [Google Scholar] [CrossRef]
- Ledesma-Escobar, C.A.; Priego-Capote, F.; Luque de Castro, M.D. Comparative study of the effect of sample pretreatment and extraction on the determination of flavonoids from lemon (Citrus limon). PLoS ONE 2016, 11, e0148056. [Google Scholar] [CrossRef]
- Castro-Vázquez, L.; Lozano, M.V.; Rodríguez-Robledo, V.; González-Fuentes, J.; Marcos, P.; Villaseca, N.; Arroyo-Jiménez, M.M.; Santander-Ortega, M.J. Pressurized Extraction as an Opportunity to Recover Antioxidants from Orange Peels: Heat treatment and Nanoemulsion Design for Modulating Oxidative Stress. Molecules 2021, 26, 5928. [Google Scholar] [CrossRef]
- Miura, M.; Makita, S.; Sugiyama, S.; Hong, Y.-J.; Yasuno, Y.; Elsner, A.E.; Tamiya, S.; Tsukahara, R.; Iwasaki, T.; Goto, H. Evaluation of intraretinal migration of retinal pigment epithelial cells in age-related macular degeneration using polarimetric imaging. Sci. Rep. 2017, 7, 3150. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, E.; Mammadzada, P.; André, H. The different facades of retinal and choroidal endothelial cells in response to hypoxia. Int. J. Mol. Sci. 2018, 19, 3846. [Google Scholar] [CrossRef] [PubMed]
- Godwin, W.C.; Hoffmann, G.F.; Gray, T.J.; Hughes, R.M. Imaging of morphological and biochemical hallmarks of apoptosis with optimized optogenetic tools. J. Biol. Chem. 2019, 294, 16918–16929. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.R.; Lawrenson, J.G. Antioxidant vitamin and mineral supplements for slowing the progression of age-related macular degeneration. Cochrane Database Syst. Rev. 2017, 7, CD000254. [Google Scholar] [CrossRef]
- Chan, C.-M.; Sekar, P.; Huang, D.-Y.; Hsu, S.-H.; Lin, W.-W. Different effects of metformin and A769662 on sodium iodate-induced cytotoxicity in retinal pigment epithelial cells: Distinct actions on mitochondrial fission and respiration. Antioxidants 2020, 9, 1057. [Google Scholar] [CrossRef]
- McCubrey, J.A.; Steelman, L.S.; Chappell, W.H.; Abrams, S.L.; Wong, E.W.; Chang, F.; Lehmann, B.; Terrian, D.M.; Milella, M.; Tafuri, A.; et al. Roles of the Raf/MEK/ERK pathway in cell growth, malignant transformation and drug resistance. Biochim. Biophys. Acta Mol. Cell Res. 2007, 1773, 1263–1284. [Google Scholar] [CrossRef]
- Wang, J.-Z.; Long, C.; Li, K.-Y.; Xu, H.-T.; Yuan, L.-L.; Wu, G.-Y. Potent block of potassium channels by MEK inhibitor U0126 in primary cultures and brain slices. Sci. Rep. 2018, 8, 8808. [Google Scholar] [CrossRef]
- Wu, Y.; Zhao, D.; Zhuang, J.; Zhang, F.; Xu, C. Caspase-8 and caspase-9 functioned differently at different stages of the cyclic stretch-induced apoptosis in human periodontal ligament cells. PLoS ONE 2016, 11, e0168268. [Google Scholar] [CrossRef]
- Sivandzade, F.; Bhalerao, A.; Cucullo, L. Analysis of the mitochondrial membrane potential using the cationic JC-1 dye as a sensitive fluorescent probe. Bio-Protocol 2019, 9, e3128. [Google Scholar] [CrossRef]
- Yako, T.; Nakamura, M.; Nakamura, S.; Hara, H.; Shimazawa, M. Pharmacological inhibition of mitochondrial fission attenuates oxidative stress-induced damage of retinal pigmented epithelial cells. J. Pharmacol. Sci. 2021, 146, 149–159. [Google Scholar] [CrossRef]
- Suri, S.; Singh, A.; Nema, P.K. Current applications of citrus fruit processing waste: A scientific outlook. Appl. Food Res. 2022, 2, 100050. [Google Scholar] [CrossRef]
- Rekha, C.; Poornima, G.; Manasa, M.; Abhipsa, V.; Devi, J.P.; Kumar, H.T.V.; Kekuda, T.R.P. Ascorbic acid, total phenol content and antioxidant activity of fresh juices of four ripe and unripe citrus fruits. Chem. Sci. Trans. 2012, 1, 303–310. [Google Scholar] [CrossRef]
- Perez-Perez, J.; Castillo, I.P.; Garcia-Lidon, A.; Botia, P.; Garcia-Sanchez, F. Fino lemon clones compared with the lemon varieties Eureka and Lisbon on two rootstocks in Murcia (Spain). Sci. Hortic. 2005, 106, 530–538. [Google Scholar] [CrossRef]
- Durmus, N.; Kilic-Akyilmaz, M. Bioactivity of non-extractable phenolics from lemon peel obtained by enzyme and ultrasound assisted extractions. Food Biosci. 2023, 53, 102571. [Google Scholar] [CrossRef]
- M’hiri, N.; Ghali, R.; Nasr, I.B.; Boudhrioua, N. Effect of different drying processes on functional properties of industrial lemon byproduct. Process Saf. Environ. Prot. 2018, 116, 450–460. [Google Scholar] [CrossRef]
- Peng, J.; Zheng, T.-t.; Liang, Y.; Duan, L.-f.; Zhang, Y.-d.; Wang, L.-J.; He, G.-m.; Xiao, H.-t. p-Coumaric acid protects human lens epithelial cells against oxidative stress-induced apoptosis by MAPK signaling. Oxid. Med. Cell. Longev. 2018, 2018, 8549052. [Google Scholar] [CrossRef]
- Huang, W.-C.; Liou, C.-J.; Shen, S.-C.; Hu, S.; Hsiao, C.-Y.; Wu, S.-J. Luteolin attenuates IL-1β-induced THP-1 adhesion to ARPE-19 cells via suppression of NF-κB and MAPK pathways. Mediators Inflamm. 2020, 2020, e9421340. [Google Scholar] [CrossRef]
- Chowers, G.; Cohen, M.; Marks-Ohana, D.; Stika, S.; Eijzenberg, A.; Banin, E.; Obolensky, A. Course of sodium iodate–induced retinal degeneration in albino and pigmented mice. Investig. Ophthalmol. Vis. Sci. 2017, 58, 2239–2249. [Google Scholar] [CrossRef]
- Wang, J.; Iacovelli, J.; Spencer, C.; Saint-Geniez, M. Direct effect of sodium iodate on neurosensory retina. Investig. Ophthalmol. Vis. Sci. 2014, 55, 1941–1953. [Google Scholar] [CrossRef]
- Zhou, P.; Kannan, R.; Spee, C.; Sreekumar, P.G.; Dou, G.; Hinton, D.R. Protection of retina by αB crystallin in sodium iodate induced retinal degeneration. PLoS ONE 2014, 9, e98275. [Google Scholar] [CrossRef]
- Kim, J.; Lee, Y.J.; Won, J.Y. Molecular mechanisms of retinal pigment epithelium dysfunction in age-related macular degeneration. Int. J. Mol. Sci. 2021, 22, 12298. [Google Scholar] [CrossRef]
- Chang, Y.-Y.; Wang, M.; Yeh, J.-H.; Tsou, S.-C.; Chen, T.-C.; Hsu, M.-Y.; Lee, Y.-J.; Wang, I.; Lin, H.-W.J.F. The protective effects of beta-mangostin against sodium iodate-induced retinal ROS-mediated apoptosis through MEK/ERK and p53 signaling pathways. Food Funct. 2023, 14, 10896–10909. [Google Scholar] [CrossRef]
- Crowley, L.C.; Marfell, B.J.; Scott, A.P.; Waterhouse, N.J. Quantitation of apoptosis and necrosis by annexin V binding, propidium iodide uptake, and flow cytometry. Cold Spring Harb. Protoc. 2016, 2016, 953–957. [Google Scholar] [CrossRef]
- Luo, X.; Gu, S.; Zhang, Y.; Zhang, J. Kinsenoside ameliorates oxidative stress-induced RPE cell apoptosis and inhibits angiogenesis via Erk/p38/NF-κB/VEGF signaling. Front. Pharmacol. 2018, 9, 240. [Google Scholar] [CrossRef]
- Tsai, H.-Y.; Lai, H.I.A.M.; Chen, Z.-Y.; Lin, T.-C.; Khor, W.; Kuo, W.-C.; Syu, J.-P.; Tsai, P.-H.; Yang, Y.-P.; Chien, Y.; et al. Inhibition of DUSP6 activates autophagy and rescues the retinal pigment epithelium in sodium iodate-induced retinal degeneration models in vivo and in vitro. Biomedicines 2022, 10, 159. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Content | |
|---|---|
| ABTS+ scavenging activity (mmole TE/g sample) | 5.65 ± 0.06 |
| DPPH scavenging activity (mmole CE/g sample) | 2.51 ± 0.03 |
| Polyphenol content (mmole GAE/g sample) | 10.24 ± 0.09 |
| Flavonoids content (µmole CE/g sample) | 964 ± 46.61 |
| No. | Compound | Neutral Mass | R.T. (min) | Adduct | Main m/z Fragments | References |
|---|---|---|---|---|---|---|
| 1 | Isocitric acid | 192.12 | 2.95 | [M+H]− | 111.00, 390.90 | [35,36] |
| 2 | Citric acid | 192.12 | 3.83 | [M+H]− | 111.00, 390.90 | [36,37,38] |
| 3 | Gentiopicroside | 356.10 | 6.53 | [M+H]− | 91.00, 191.00, 719.05 | [37] |
| 4 | Ferulic acid hexoside | 356.32 | 7.57 | [M+H]− | 91.00, 191.00, 615.15 | [38] |
| 5 | Quercetin-3-O-rutinoside-7-O-glucoside | 772.66 | 10.12 | [M+H]− | 365.05 | [39] |
| 6 | Quinic acid | 192.06 | 10.55 | [M+H]+ | 83.05, 354.95, 523.05 | [35] |
| 7 | Limocitrol-O-Glu | 538.46 | 14.72 | [M+H]+ | 83.05 | [38] |
| 8 | Chrysoeriol-6,8-di-C-Glu/stellarin-2 | 624.50 | 15.20 | [M+H]− | 91.00, 757.10 | [38,40,41,42] |
| 9 | p-Coumaroyl quinic acid | 338.10 | 15.73 | [M+H]− | 391.05, 577.05, 755.10 | [36] |
| 10 | Apigenin-7-O-Neo/rhoifolin | 578.16 | 15.73 | [M+H]− | 336.95, 391.05, 755.10 | [38,40,42] |
| 11 | Limocitrin-Glu-HMG-Glu | 816.22 | 18.10 | [M+H]− | 391.05 | [35,38,43] |
| 12 | Sinapoyl-O-glucoside | 386.35 | 20.23 | [M+H]− | 91.00, 336.90 | [36,38] |
| 13 | Eriodictyol-7-O-Rut/neoeriocitrin | 596.17 | 21.67 | [M+H]− | 659.15 | [35,43] |
| 14 | Naringenin-7-O-Neo/naringin | 580.18 | 26.30 | [M+H]− | 91.00, 501.00 | [35,39,41,43] |
| 15 | Hesperetin | 302.08 | 29.17 | [M+H]− | 471.05, 681.15 | [35,36,39,43] |
| 16 | Hesperetin-7-O-Rut/hesperidin | 610.19 | 29.32 | [M+H]− | 300.95, 471.05 | [35,39,41,43,44] |
| 17 | Hesperetin-7-O-Neo/neohesperidin | 610.19 | 29.32 | [M+H]− | 301.10, 471.05 | [35,36,43] |
| 18 | Limonexic acid/limonexin | 502.52 | 30.98 | [M+H]+ | 83.05, 687.10 | [36] |
| 19 | Limocitrol-O-Glu-HMG | 682.17 | 31.38 | [M+H]− | 579.05 | [35,36,43] |
| 20 | Naringenin-7-O-Neo/naringin | 580.18 | 32.10 | [M+H]− | 91.05, 409.10, 543.10 | [35,39,41,43] |
| 21 | p-Coumatric acid | 164.05 | 32.95 | [M+H]− | 119.05, 350.95 | [36] |
| 22 | Luteolin | 286.24 | 34.52 | [M+H]+ | 83.05 | [36,39] |
| 23 | Limocitrin | 346.29 | 34.92 | [M+H]+ | 83.05, 797.10 | [36] |
| 24 | Limocitrol | 376.31 | 39.40 | [M+H]− | 91.05, 435.05, 841.15, | [36] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsou, S.-C.; Chuang, C.-J.; Wang, I.; Chen, T.-C.; Yeh, J.-H.; Hsu, C.-L.; Hung, Y.-C.; Lee, M.-C.; Chang, Y.-Y.; Lin, H.-W. Lemon Peel Water Extract: A Novel Material for Retinal Health, Protecting Retinal Pigment Epithelial Cells against Dynamin-Related Protein 1-Mediated Mitochondrial Fission by Blocking ROS-Stimulated Mitogen-Activated Protein Kinase/Extracellular Signal-Regulated Kinase Pathway. Antioxidants 2024, 13, 538. https://doi.org/10.3390/antiox13050538
Tsou S-C, Chuang C-J, Wang I, Chen T-C, Yeh J-H, Hsu C-L, Hung Y-C, Lee M-C, Chang Y-Y, Lin H-W. Lemon Peel Water Extract: A Novel Material for Retinal Health, Protecting Retinal Pigment Epithelial Cells against Dynamin-Related Protein 1-Mediated Mitochondrial Fission by Blocking ROS-Stimulated Mitogen-Activated Protein Kinase/Extracellular Signal-Regulated Kinase Pathway. Antioxidants. 2024; 13(5):538. https://doi.org/10.3390/antiox13050538
Chicago/Turabian StyleTsou, Shang-Chun, Chen-Ju Chuang, Inga Wang, Tzu-Chun Chen, Jui-Hsuan Yeh, Chin-Lin Hsu, Yu-Chien Hung, Ming-Chung Lee, Yuan-Yen Chang, and Hui-Wen Lin. 2024. "Lemon Peel Water Extract: A Novel Material for Retinal Health, Protecting Retinal Pigment Epithelial Cells against Dynamin-Related Protein 1-Mediated Mitochondrial Fission by Blocking ROS-Stimulated Mitogen-Activated Protein Kinase/Extracellular Signal-Regulated Kinase Pathway" Antioxidants 13, no. 5: 538. https://doi.org/10.3390/antiox13050538