
Screening of Antioxidative and Antiproliferative Activities of Crude Polysaccharides Extracted from Six Different Plants
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Selected Plants and Chemicals
2.2. Preparation of Polysaccharide Extracts from the Selected Plants
2.3. Chemical Composition of the Plant Polysaccharide Extracts
2.4. Assessment of Free-Radical-Scavenging Activities of Plant Polysaccharides
2.4.1. Determination of DPPH Radical-Scavenging Activity
2.4.2. Determination of Nitric Oxide Radical-Scavenging Activity
2.4.3. Determination of Ferric-Reducing Antioxidant Power (FRAP)
2.4.4. Determination of Cupric-Reducing Antioxidant Capacity (CUPRAC)
2.5. Antiproliferative Activities of Plant Polysaccharide
2.5.1. Cultivation of MCF-7, MRC-5, and Vero Cell Lines
2.5.2. Cytotoxicity Assay
2.5.3. Cell Proliferation Assay
2.6. Elucidation of Crude Polysaccharide Structure by Nuclear Magnetic Resonance Analysis
2.7. Mass Spectrometry Study
2.8. Statistical Analysis
3. Results
3.1. Percentage Yield and the Chemical Composition of Selected Plant Crude Polysaccharide Extracts
3.2. Assessment of Free Radical-Scavenging Activities of Plant Polysaccharide Extracts
3.2.1. DPPH Radical-Scavenging Activity

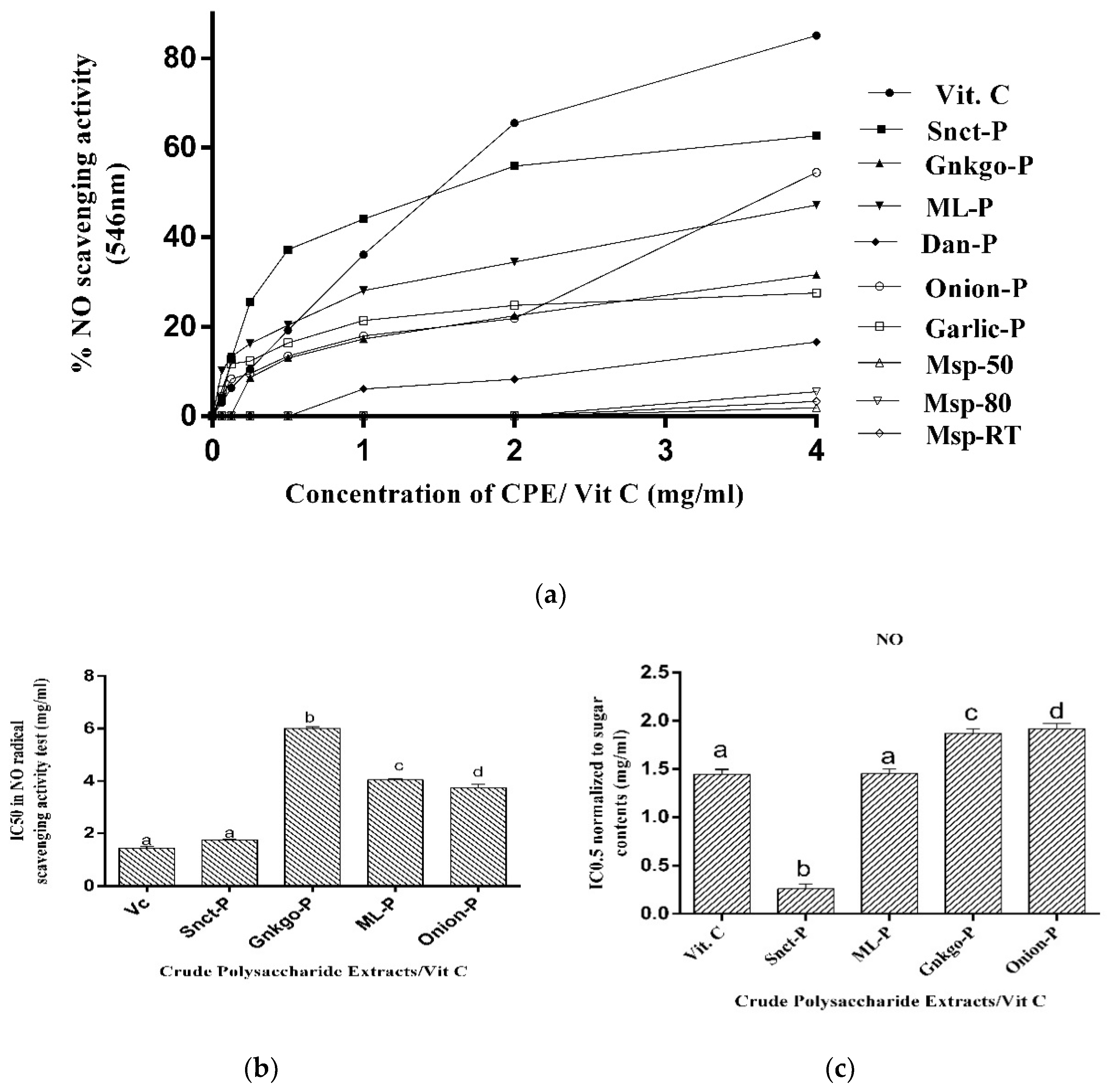
3.2.2. Determination of Nitric Oxide Radical-Scavenging Activity

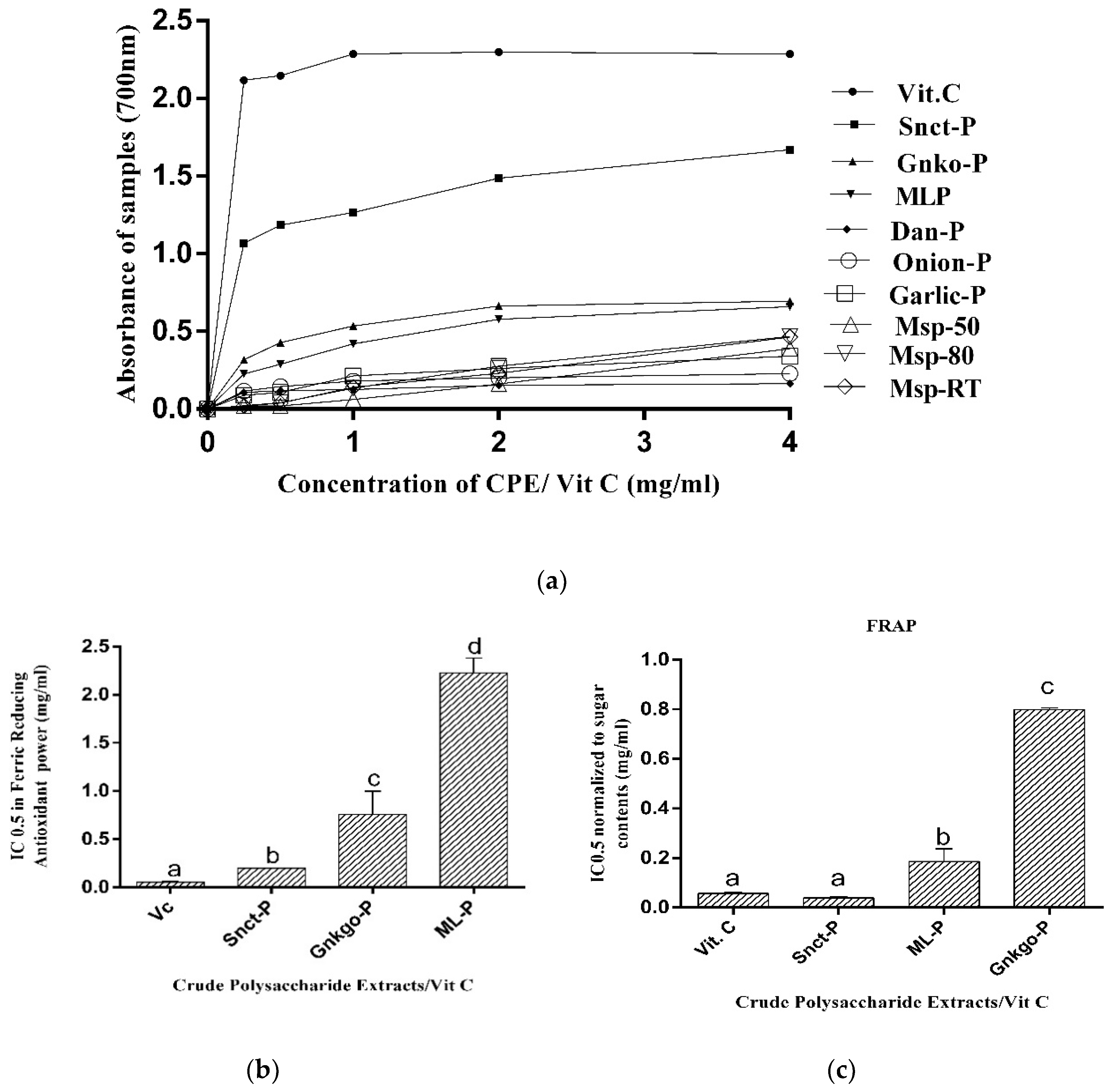
3.2.3. Determination of Ferric-Reducing Antioxidant Power (FRAP)

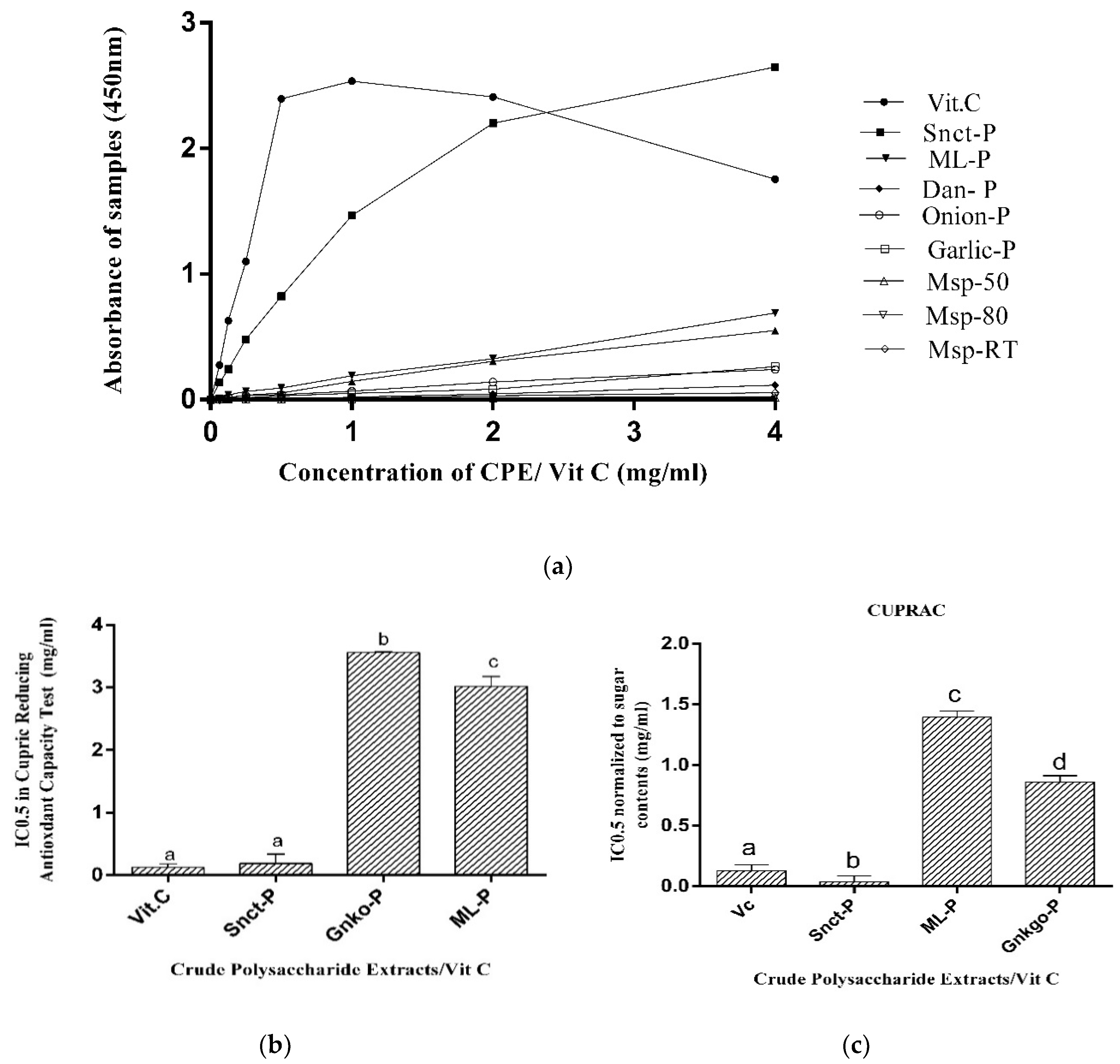
3.2.4. Determination of Cupric-Reducing Antioxidant Capacity (CUPRAC)

3.3. Cytotoxic Activities of Plant Polysaccharides in MCF-7 Cells
3.4. Effect of Plant Polysaccharide on MCF-7 Cell Proliferation
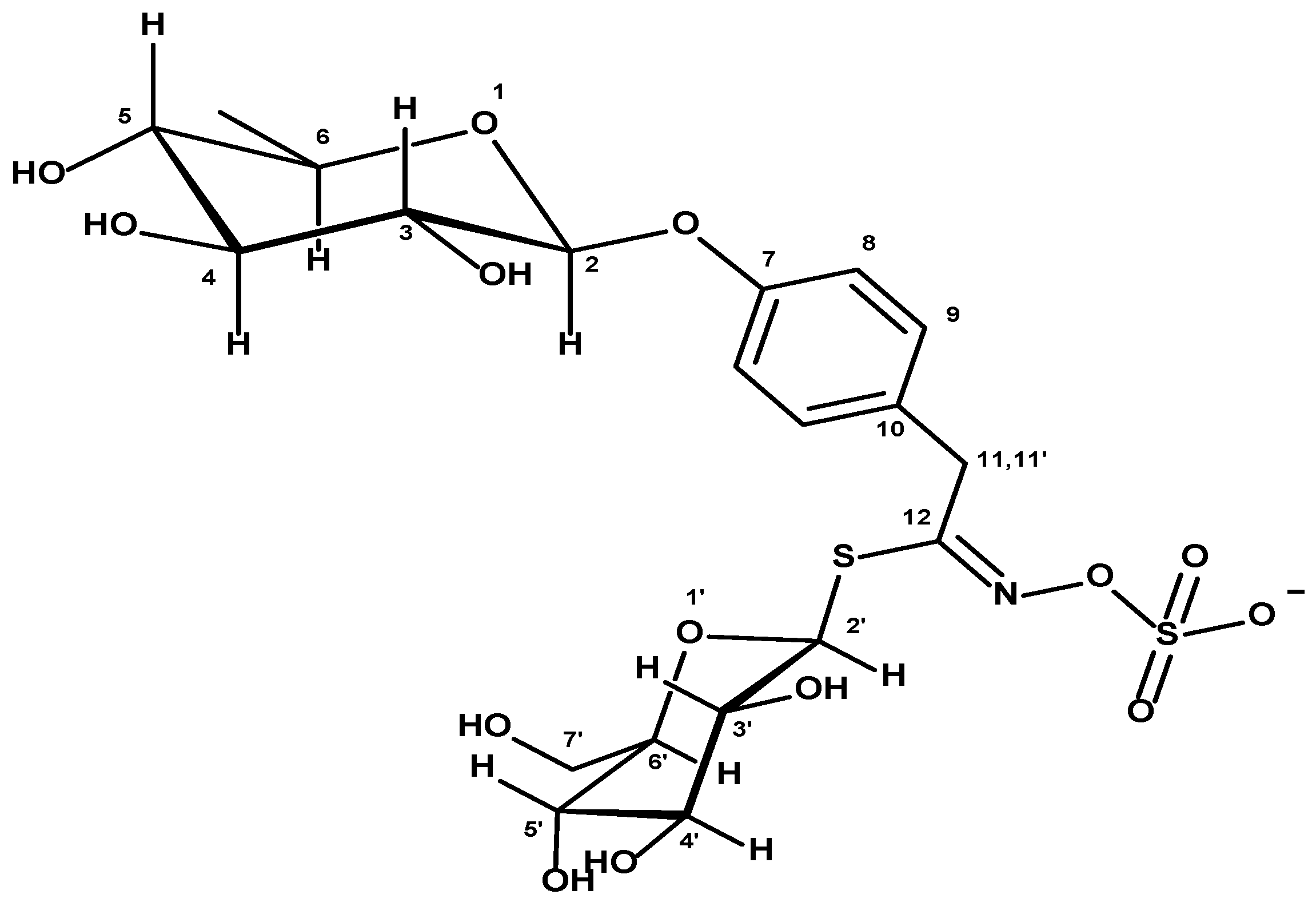
3.5. Elucidation of the MSP-RT-Abundant Compound Structure by Nuclear Magnetic Resonance Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilkinson, L.; Gathani, T. Understanding breast cancer as a global health concern. Brit. J. Radiol. 2022, 95, 20211033. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Spezia, M.; Huang, S.; Yuan, C.; Zeng, Z.; Zhang, L.; Ren, G. Breast cancer development and progression: Risk factors, cancer stem cells, signaling pathways, genomics, and molecular pathogenesis. Genes Dis. 2018, 5, 77–106. [Google Scholar] [CrossRef] [PubMed]
- Testa, U.; Castelli, G.; Pelosi, E. Breast cancer: A molecularly heterogenous disease needing subtype-specific treatments. Med. Sci. 2020, 8, 18. [Google Scholar] [CrossRef]
- Vuger, A.T.; Tiscoski, K.; Apolinario, T.; Cardoso, F. Anthracyclines in the treatment of early breast cancer friend or foe? Breast 2022, 65, 67–76. [Google Scholar] [CrossRef]
- Willson, M.L.; Burke, L.; Ferguson, T.; Ghersi, D.; Nowak, A.K.; Wilcken, N. Taxanes for adjuvant treatment of early breast cancer. Cochrane Database Syst. Rev. 2019, 9, 1–140. [Google Scholar] [CrossRef]
- Du, J.; Dong, Y.; Li, Y. Identification and prognostic value exploration of cyclophosphamide (Cytoxan)-centered chemotherapy response-associated genes in breast cancer. DNA Cell Biol. 2021, 40, 1356–1368. [Google Scholar] [CrossRef]
- Lin, C.; Cui, J.; Peng, Z.; Qian, K.; Wu, R.; Cheng, Y.; Yin, W. Efficacy of platinum-based and non-platinum-based drugs on triple-negative breast cancer: Meta-analysis. Eur. J. Med. Res. 2022, 27, 201. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, S.M.; Pham, A.T.; Nguyen, L.M.; Cai, H.; Tran, T.V.; Shu, X.O.; Tran, H.T. Chemotherapy-Induced Toxicities and Their Associations with Clinical and Non-Clinical Factors among Breast Cancer Patients in Vietnam. Curr. Oncol. 2022, 29, 8269–8284. [Google Scholar] [CrossRef]
- Greenwell, M.; Rahman, P.K.S.M. Medicinal plants: Their use in anticancer treatment. Int. J. Pharm. Sci. Res. 2015, 6, 4103. [Google Scholar] [CrossRef] [PubMed]
- Stępień, A.E.; Trojniak, J.; Tabarkiewicz, J. Health-Promoting Properties: Anti-Inflammatory and Anticancer Properties of Sambucus nigra L. Flowers and Fruits. Molecules 2023, 28, 6235. [Google Scholar] [CrossRef]
- Ogunro, O.B.; Fakayode, A.E.; Batiha, G.E.S. Involvement of Antioxidants in the Prevention of Cellular Damage. In Importance of Oxidative Stress and Antioxidant System in Health and Disease; IntechOpen: London, UK, 2022; p. 10873. [Google Scholar] [CrossRef]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants maintain cellular redox homeostasis by elimination of reactive oxygen species. Cell Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef]
- Didier, A.J.; Stiene, J.; Fang, L.; Watkins, D.; Dworkin, L.D.; Creeden, J.F. Antioxidant and Anti-Tumor Effects of Dietary Vitamins A, C, and E. Antioxidants 2023, 12, 632. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxidative Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Xu, D.; Zhou, Y.M.; Zhang, Y.B.; Zhang, H.; Chen, Y.B.; Cui, Y.L. Antioxidant activities of natural polysaccharides and their derivatives for biomedical and medicinal applications. Antioxidants 2022, 11, 2491. [Google Scholar] [CrossRef]
- Jin, H.; Li, M.; Tian, F.; Yu, F.; Zhao, W. An Overview of Antitumour Activity of Polysaccharides. Molecules 2022, 27, 8083. [Google Scholar] [CrossRef]
- Wang, A.; Liu, Y.; Zeng, S.; Liu, Y.; Li, W.; Wu, D.; Wu, X.; Zou, L.; Chen, H. Dietary Plant Polysaccharides for Cancer Prevention: Role of Immune Cells and Gut Microbiota, Challenges and Perspectives. Nutrients 2023, 15, 3019. [Google Scholar] [CrossRef]
- Lemieszek, M.; Rzeski, W. Anticancer properties of polysaccharides isolated from fungi of the Basidiomycetes class. Contemp. Oncol. 2012, 16, 285–289. [Google Scholar] [CrossRef]
- Garcia, J.; Rodrigues, F.; Saavedra, M.J.; Nunes, F.M.; Marques, G. Bioactive polysaccharides from medicinal mushrooms: A review on their isolation, structural characteristics and antitumor activity. Food Biosci. 2022, 49, 101955. [Google Scholar] [CrossRef]
- Ying, Y.; Hao, W. Immunomodulatory function and anti-tumor mechanism of natural polysaccharides: A review. Front. Immunol. 2023, 14, 1147641. [Google Scholar] [CrossRef]
- Banerjee, S.; Parasramka, M.; Paruthy, S.B. Polysaccharides in cancer prevention: From bench to bedside. In Polysaccharides: Bioactivity and Biotechnology, 1st ed.; Ramawat, K.G., Mérillon, J.M., Eds.; Springer: Cham, Switzerland, 2015; pp. 2179–2214. [Google Scholar]
- Guo, R.; Chen, M.; Ding, Y.; Yang, P.; Wang, M.; Zhang, H.; Ma, H. Polysaccharides as potential anti-tumor biomacromolecules—A review. Front. Nutr. 2022, 9, 838179. [Google Scholar] [CrossRef]
- Chen, L.; He, C.; Zhou, M.; Long, J.; Li, L. Research Progress on the Mechanisms of Polysaccharides against Gastric Cancer. Molecules 2022, 27, 5828. [Google Scholar] [CrossRef] [PubMed]
- Satoh, Y.; Goi, T.; Nakazawa, T.; Kimura, Y.; Hirono, Y.; Katayama, K.; Yamaguchi, A. Polysaccharide K suppresses angiogenesis in colon cancer cells. Exp. Ther. Med. 2012, 4, 370–374. [Google Scholar] [CrossRef] [PubMed]
- Das, T.K.; Wati, M.R.; Fatima-Shad, K. Oxidative stress gated by Fenton and Haber Weiss reactions and its association with Alzheimer’s disease. Arch. Neurosci. 2015, 2, e60038. [Google Scholar] [CrossRef]
- Lis, B.; Olas, B. Pro-health activity of dandelion (Taraxacum officinale L.) and its food products–history and present. J. Funct. Foods 2019, 59, 40–48. [Google Scholar] [CrossRef]
- Kania-Dobrowolska, M.; Baraniak, J. Dandelion (Taraxacum officinale L.) as a source of biologically active compounds supporting the therapy of co-existing diseases in metabolic syndrome. Foods 2022, 11, 2858. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjya, D.; Adhikari, S.; Biswas, A.; Bhuimali, A.; Ghosh, P.; Saha, S. Ocimum phytochemicals and their potential impact on human health. Phytochem. Hum. Health 2019, 23, 1–26. [Google Scholar]
- Islam, Z.; Islam, S.M.; Hossen, F.; Mahtab-ul-Islam, K.; Hasan, M.R.; Karim, R. Moringa oleifera is a prominent source of nutrients with potential health benefits. Int. J. Food Sci. 2021, 2021, 6627265. [Google Scholar] [CrossRef] [PubMed]
- Mehwish, H.M.; Liu, G.; Rajoka, M.S.R.; Cai, H.; Zhong, J.; Song, X.; He, Z. Therapeutic potential of Moringa oleifera seed polysaccharide embedded silver nanoparticles in wound healing. Int. J. Biol. Macromol. 2021, 184, 144–158. [Google Scholar] [CrossRef]
- Kiple, K.F.; Ornelas, K.C. (Eds.) The Cambridge World History of Food; Cambridge University Press: Cambridge, UK, 2000; Volume 2. [Google Scholar]
- Santhosha, S.G.; Jamuna, P.; Prabhavathi, S.N. Bioactive components of garlic and their physiological role in health maintenance: A review. Food Biosci. 2013, 3, 59–74. [Google Scholar] [CrossRef]
- Arreola, R.; Quintero-Fabián, S.; López-Roa, R.I.; Flores-Gutiérrez, E.O.; Reyes-Grajeda, J.P.; Carrera-Quintanar, L.; Ortuño-Sahagún, D. Immunomodulation and anti-inflammatory effects of garlic compounds. J. Immunol. Res. 2015, 2015, 401630. [Google Scholar] [CrossRef]
- Kumari, N.; Kumar, M.; Lorenzo, J.M.; Sharma, D.; Puri, S.; Pundir, A.; Dhumal, S.; Bhuyan, D.J.; Jayanthy, G.; Selim, S.; et al. Onion and garlic polysaccharides: A review on extraction, characterization, bioactivity, and modifications. Int. J. Biol. Macromol. 2022, 219, 1047–1061. [Google Scholar] [CrossRef] [PubMed]
- Jesumani, V.; Du, H.; Pei, P.; Aslam, M.; Huang, N. Comparative study on skin protection activity of polyphenol-rich extract and polysaccharide-rich extract from Sargassum vachellianum. PLoS ONE 2020, 15, e0227308. [Google Scholar] [CrossRef] [PubMed]
- Dare, C.A.; Oyedapo, O.O.; Akinlalu, A.O.; Komolafe, I.J.; Fajobi, A.O.; Ogunsusi, M. Genotoxic Activities of Polysaccharides from Cotyledon and Coat of Fermented and Unfermented Annona squamosa L. Seed. Egypt. Acad. J. Biol. Sci. H. Bot. 2021, 12, 189–207. [Google Scholar] [CrossRef]
- Wu, H.; Shang, H.; Guo, Y.; Zhang, H.; Wu, H. Comparison of different extraction methods of polysaccharides from cup plant (Silphium perfoliatum L.). Process Biochem. 2020, 90, 241–248. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the Principle of protein-dye binding. Anal Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Akyüz, M. Nutritive value, flavonoid content, and radical scavenging activity of the truffle (Terfezia Boudieri Chatin). J. Soil Sci. Plant Nutr. 2013, 13, 143–151. [Google Scholar] [CrossRef]
- Kury, L.T.A.; Taha, Z.; Talib, W.H. Immunomodulatory and anticancer activities of Hyacinthus L.: An in vitro and in vivo study. Plants 2021, 10, 617. [Google Scholar] [CrossRef] [PubMed]
- Bouhlali, E.D.T.; Alem, C.; Zegzouti, Y.F. Antioxidant and anti-hemolytic activities of phenolic constituents of six Moroccan date fruit (Phoenix dactylifera L.) syrups. Indian J. Biotechnol. 2016, 12, 45–52. [Google Scholar]
- Marcocci, L.; Maguire, J.J.; Droyl-lefaix, M.T.; Packer, L. The nitric oxide scavenging properties of Ginkgo biloba extract. Biochem. Biophys. Res. Commun. 1994, 201, 748–755. [Google Scholar] [CrossRef]
- Etim, O.E.; Ekanem, S.E.; Sam, S.M. In vitro antioxidant activity and nitric oxide scavenging activity of Citrullus lanatus seeds. In Vitro 2013, 3, 126–132. [Google Scholar]
- Oyaizu, M. Studies on products of browning reaction: Antioxidative activities of products of browning reaction prepared from glucosamine. Jpn. J. Nutr. Diet. 1986, 44, 307–315. [Google Scholar] [CrossRef]
- Apak, R.; Güçlü, K.; Özyürek, M.; Burcu Bektas, B.; Bener, M. Cupric ion reducing antioxidant capacity assay for food antioxidants: Vitamins, polyphenolics, and flavonoids in food extracts. In Advanced Protocols in Oxidative Stress; Humana Press: Totowa, NJ, USA, 2008; pp. 163–193. [Google Scholar] [CrossRef]
- Niyigaba, T.; Liu, D.; Habimana, J.D.D. The extraction, functionalities, and applications of plant polysaccharides in fermented foods: A review. Foods 2021, 10, 3004. [Google Scholar] [CrossRef]
- Ahmad, M.M.; Chatha, S.A.S.; Iqbal, Y.; Hussain, A.I.; Khan, I.; Xie, F. Recent trends in extraction, purification, and antioxidant activity evaluation of plant leaf-extract polysaccharides. Biofuels Bioprod. Biorefin. 2022, 16, 1820–1848. [Google Scholar] [CrossRef]
- Cai, L.; Chen, B.; Yi, F.; Zou, S. Optimization of extraction of polysaccharide from dandelion root by response surface methodology: Structural characterization and antioxidant activity. Int. J. Biol. Macromol. 2019, 140, 907–919. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Jiang, Y.; Zhao, M.; Shi, J.; Wang, L. Effects of ultrasonic extraction on the physical and chemical properties of polysa-ccharides from longan fruit pericarp. Polym. Degrad. Stab. 2008, 93, 268–272. [Google Scholar] [CrossRef]
- Zhou, S.; Huang, G.; Huang, H. Extraction, derivatization and antioxidant activities of onion polysaccharide. Food Chem. 2022, 388, 133000. [Google Scholar] [CrossRef]
- Awad, A.M.; Kumar, P.; Ismail-Fitry, M.R.; Jusoh, S.; Ab Aziz, M.F.; Sazili, A.Q. Green extraction of bioactive compounds from plant biomass and their application in meat as a natural antioxidant. Antioxidants 2021, 10, 1465. [Google Scholar] [CrossRef]
- Aryal, S.; Baniya, M.K.; Danekhu, K.; Kunwar, P.; Gurung, R.; Koirala, N. Total phenolic content, flavonoid content and antioxidant potential of wild vegetables from Western Nepal. Plants 2019, 8, 96. [Google Scholar] [CrossRef]
- Baliyan, S.; Mukherjee, R.; Priyadarshini, A.; Vibhuti, A.; Gupta, A.; Pandey, R.P.; Chang, C.M. Determination of antioxidants by DPPH radical scavenging activity and quantitative phytochemical analysis of Ficus religiosa. Molecules 2022, 27, 1326. [Google Scholar] [CrossRef]
- Tokumaru, O.; Shuto, Y.; Ogata, K.; Kamibayashi, M.; Bacal, K.; Takei, H.; Kitano, T. Dose-dependency of multiple free radical-scavenging activity of edaravone. J. Surg. Res. 2018, 228, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Adewale, O.O.; Oyelola, R.F.; Oladele, J.O.; Agbaje, W.B. Assessing the ability of Polysaccharides extracted from Date Palm Fruit to salvage Wistar rats from cisplatin-linked hepatic damage. Pharmacol. Res.- Mod. Chin. Med. 2024, 11, 100400. [Google Scholar] [CrossRef]
- Al-Amiery, A.A.; Al-Majedy, Y.K.; Kadhum, A.A.H.; Mohamad, A.B. Hydrogen peroxide scavenging activity of novel coumarins synthesized using different approaches. PLoS ONE 2015, 10, e0132175. [Google Scholar] [CrossRef]
- Lalhminghlui, K.; Jagetia, G.C. Evaluation of the free-radical scavenging and antioxidant activities of Chilauni, Schima wallichii Korth in vitro. Future Sci. OA 2018, 4, FSO272. [Google Scholar] [CrossRef] [PubMed]
- Benzie, I.F.; Devaki, M. The ferric reducing/antioxidant power (FRAP) assay for non-enzymatic antioxidant capacity: Concepts, procedures, limitations and applications. In Measurement of Antioxidant Activity & Capacity: Recent Trends and Applications; Wiley: Hoboken, NJ, USA, 2018; pp. 77–106. [Google Scholar] [CrossRef]
- Ahmad, M.M. Characterization and antioxidant activities of polysaccharides extracted from flageolet bean pods waste. Curr. Res. Green Sustain. Chem. 2021, 4, 100154. [Google Scholar] [CrossRef]
- Gao, J.; Zhang, T.; Jin, Z.Y.; Xu, X.M.; Wang, J.H.; Zha, X.Q.; Chen, H.Q. Structural characterisation, physicochemical properties and antioxidant activity of polysaccharide from Lilium lancifolium Thunb. Food Chem. 2015, 169, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Munteanu, I.G.; Apetrei, C. Analytical methods used in determining antioxidant activity: A review. Int. J. Mol. Sci. 2021, 22, 3380. [Google Scholar] [CrossRef] [PubMed]
- Silvestrini, A.; Meucci, E.; Ricerca, B.M.; Mancini, A. Total Antioxidant Capacity: Biochemical aspects and clinical significance. Int. J. Mol. Sci. 2023, 24, 10978. [Google Scholar] [CrossRef]
- Camarillo, I.G.; Xiao, F.; Madhivanan, S.; Salameh, T.; Nichols, M.; Reece, L.M.; Sundararajan, R. Low and high voltage electrochemotherapy for breast cancer: An in vitro model study. In Electroporation-Based Therapies for Cancer, 1st ed.; Sundararajan, R., Ed.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 55–102. [Google Scholar]
- Ghoderao, P.; Sahare, S.; Kulkarni, A.A.; Bhave, T. Paclitaxel-conjugated magnetic carbon nanotubes induce apoptosis in breast cancer cells and breast cancer stem cells in vitro. In Paclitaxel; Academic Press: Cambridge, MA, USA, 2022; pp. 309–331. [Google Scholar] [CrossRef]
- Patel, M.K.; Tanna, B.; Gupta, H.; Mishra, A.; Jha, B. Physicochemical, scavenging and anti-proliferative analyses of polysaccharides extracted from psyllium (Plantago ovata Forssk) husk and seeds. Int. J. Biol. Macromol. 2019, 133, 190–201. [Google Scholar] [CrossRef]
- Chang, T.; Li, H.; Lv, H.; Tan, M.; Hou, S.; Liu, X.; Zhao, B. Extraction, Physicochemical Properties, Anti-Aging, and Antioxidant Activities of Polysaccharides from Industrial Hemp Residues. Molecules 2022, 27, 5746. [Google Scholar] [CrossRef]
- Chang, P.; XU, Y.; Zhou, D.Y.; WU, J.D. Effects of polysaccharides from Ginkgo biloba on the proliferation of 4T1 breast cancer cells and expression of GLUT family genes. Chin. Pharmacol. Bull. 2018, 34, 1301–1307. [Google Scholar]
- Ren, F.; Wu, K.; Yang, Y.; Yang, Y.; Wang, Y.; Li, J. Dandelion polysaccharide exerts anti-angiogenesis effect on hepatocellular carcinoma by regulating VEGF/HIF-1α expression. Front. Pharmacol. 2020, 11, 460. [Google Scholar] [CrossRef]
- Adebayo, I.A.; Arsad, H.; Samian, M.R. Antiproliferative effect on breast cancer (MCF7) of Moringa oleifera seed extracts. Afr. J. Tradit. Complement. Altern. Med. 2017, 14, 282–287. [Google Scholar] [CrossRef]
- Ramarao, K.D.R.; Somasundram, C.; Razali, Z.; Kunasekaran, W.; Jin, T.L.; Musa, S.; Achari, V.M. Antiproliferative effects of dried Moringa oleifera leaf extract on human Wharton’s Jelly mesenchymal stem cells. PLoS ONE 2022, 17, e0274814. [Google Scholar] [CrossRef] [PubMed]
- Richards, M.; Phoon, C.W.; Goh, G.T.W.; Seng, E.K.; Guo, X.M.; Tan, C.M.F.; Lee, J.M.K. A new class of pluripotent stem cell cytotoxic small molecules. PLoS ONE 2014, 9, e85039. [Google Scholar] [CrossRef] [PubMed]
- Konishi, K.; Yamaji, T.; Sakuma, C.; Kasai, F.; Endo, T.; Kohara, A.; Osada, N. Whole- Genome Sequencing of Vero E6 (VEROC1008) and Comparative Analysis of Four Vero Cell Sublines. Front. Genet. 2022, 13, 801382. [Google Scholar] [CrossRef] [PubMed]
- Abdi Goushbolagh, N.; Farhood, B.; Astani, A.; Nikfarjam, A.; Kalantari, M.; Zare, M.H. Quantitative cytotoxicity, cellular uptake and radioprotection effect of cerium oxide nanoparticles in MRC-5 normal cells and MCF-7 cancerous cells. BioNanoScience 2018, 8, 769–777. [Google Scholar] [CrossRef]
- Swanson, S.K.; Mento, S.J.; Weeks-Levy, C.; Brock, B.D.; Kowal, K.J.; Wallace, R.E.; Cano, F.R. Characterization of Vero cells. J. Biol. Stand. 1988, 16, 311–320. [Google Scholar] [CrossRef]
- Shakeri, M.; Tayer, A.H.; Shakeri, H.; Jahromi, A.S.; Moradzadeh, M.; Hojjat-Farsangi, M. Toxicity of saffron extracts on cancer and normal cells: A review article. Asian Pac. J. Cancer Prev. 2020, 21, 1867. [Google Scholar] [CrossRef]
- Komura, M.; Sato, T.; Yoshikawa, H.; Nitta, N.A.; Suzuki, Y.; Koike, K.; Takahashi, K. Propylene glycol, a component of electronic cigarette liquid, damages epithelial cells in human small airways. Respir. Res. 2022, 23, 216. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, H.; Chen, X. Drug resistance and combating drug resistance in cancer. Cancer Drug Resist. 2019, 2, 141. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Rodriguez, N.A.; Gaytán-Martínez, M.; de la Luz Reyes-Vega, M.; Loarca-Piña, G. Glucosinolates and isothiocyanates from Moringa oleifera: Chemical and biological approaches. Plant Foods Hum. Nutr. 2020, 75, 447–457. [Google Scholar] [CrossRef]
- Bell, L.; Oloyede, O.O.; Lignou, S.; Wagstaff, C.; Methven, L. Taste and flavor perceptions of glucosinolates, isothiocyanates, and related compounds. Mol. Nutr. Food Res. 2018, 62, 1700990. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Tang, H.; Jia, X.; Zheng, C.; Huang, F.; Zhang, M. Distribution of glucosinolate and pungent odors in rapeseed oils from raw and microwaved seeds. Int. J. Food Prop. 2018, 21, 2296–2308. [Google Scholar] [CrossRef]
- Sanlier, N.; Guler, S.M. The benefits of Brassica vegetables on human health. J. Hum. Health Res. 2018, 1, 1–13. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Plant Species | Where They Were Obtained | Part of Plant Used | Form of Plant Used |
|---|---|---|---|---|
| Snct-P | Ocimum sanctum | Eco-Herba, Poland | Leaves | Dried leaves |
| ML-P | Moringa oleifera | Osun State, Nigeria | Leaves | Dried leaves |
| Ginkgo-P | Ginkgo biloba | Eco-Herba, Poland | Leaves | Dried leaves |
| Onion-P | Allium cepa | Local market, Poland | Bulbs | Fresh bulbs |
| Garlic-P | Allium sativum | Local market, Poland | Leaves | Dried leaves |
| Dan-P | T. officinale | Eco-Herba, Poland | Roots | Dried roots |
| MSP-50 | Moringa oleifera | Osun State, Nigeria | Seeds | Dried seeds |
| MSP-80 | Moringa oleifera | Osun State, Nigeria | Seeds | Dried seeds |
| MSP-RT | Moringa oleifera | Osun State, Nigeria | Seeds | Dried seeds |
| Samples | * Yield (%) | Total Sugar Content (%) | Protein Content (%) | Phenolic Content (%) |
|---|---|---|---|---|
| Snct-P | 13.28 ± 0.78 a | 17.70 ± 0.03 a | 9.27 ± 0.30 a | 16.50 ± 0.56 a |
| ML-P | 8.23 ± 1.1 a | 46.14 ± 1.16 c | 11.81 ± 0.30 ac | 18.44 ± 0.80 c,b |
| Gnkgo-P | 7.11 ± 0.45 a,d | 24.08 ± 1.88 d | 1.45 ± 0.07 d,b | 7.78 ± 1.43 d,b |
| * Onion-P | 0.60 ± 0.05 | 51.19 ± 0.72 e | 2.19 ± 0.30 e,b | 1.51 ± 0.03 e,b,d |
| Garlic-P | 9.39 ± 0.57 a | 24.71 ± 2.42 f | 0.67 ± 0.01 f,b | 3.18 ± 0.15 f,b,d,e |
| Dan-P | 27.88 ± 4.56 g | 83.78 ± 1.28 g | 0.15 ± 0.01 g,b | 1.26 ± 0.02 g,d,e |
| MSP-50 | 6.41 ± 0.89 h | 6.30 ± 1.45 h,i | 31.76 ± 1.70 h | 2.18 ± 0.01 h,b,c,d |
| MSP-80 | 11.09 ± 0.31 a,h,i | 7.86 ± 0.38 i | 32.89 ± 0.70 i,h | 2.68 ± 0.02 i,c |
| MSP-RT | 13.97 ± 1.22 i | 9.00 ± 0.89 j,i | 34.38 ± 0.22 j,h | 2.35 ± 0.02 j,c,d |
| Crude Polysaccharides | |||
|---|---|---|---|
| MSP-50 | MSP-80 | MSP-RT | |
| IC50 | 0.3415 | 0.3411 | 0.06073 |
| R2 | 0.9945 | 0.996 | 0.9929 |
| Cell Line | ||
|---|---|---|
| MRC-5 | Vero | |
| IC50 | 0.1574 | 0.1594 |
| R2 | 0.9989 | 0.9626 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adewale, O.O.; Wińska, P.; Krawczyk, H.; Grzechnik, E.; Cieśla, J. Screening of Antioxidative and Antiproliferative Activities of Crude Polysaccharides Extracted from Six Different Plants. Appl. Sci. 2024, 14, 3683. https://doi.org/10.3390/app14093683
Adewale OO, Wińska P, Krawczyk H, Grzechnik E, Cieśla J. Screening of Antioxidative and Antiproliferative Activities of Crude Polysaccharides Extracted from Six Different Plants. Applied Sciences. 2024; 14(9):3683. https://doi.org/10.3390/app14093683
Chicago/Turabian StyleAdewale, Omowumi Oyeronke, Patrycja Wińska, Hanna Krawczyk, Eryk Grzechnik, and Joanna Cieśla. 2024. "Screening of Antioxidative and Antiproliferative Activities of Crude Polysaccharides Extracted from Six Different Plants" Applied Sciences 14, no. 9: 3683. https://doi.org/10.3390/app14093683